CERKL通过激活SIRT1/E2F1轴减轻蓝光导致的人视网膜色素上皮细胞氧化应激损伤
2022-08-09庄海容吴子东陈雪红李成军
庄海容,吴子东,陈雪红,李成军
0引言
随着科技的发展与电子产品的广泛使用,人们越来越重视蓝光辐射对视网膜的危害。视网膜色素上皮(RPE)位于视网膜最外层,在维持光感受器功能和结构方面起着极为重要的生理作用[1]。大量证据显示,持续蓝光暴露会诱发RPE细胞凋亡,而RPE细胞中自身代谢产物的积累很容易引发氧化应激,导致光感受器功能受损,严重危害视力健康[2-3]。神经酰胺类似蛋白(CERKL)是导致视网膜退化的重要致病基因[4-5]。Mandal等[6]研究发现蓝光可诱导RPE中CERKL的表达。Riera等研究表明,CERKL可以使视网膜细胞形成氧化应激保护[7]。由此推测:CERKL可能通过调节细胞内的氧化应激途径来保护视网膜组织免受蓝光损伤,但该推测的正确性与其中的分子机制仍有待进一步研究。沉默信息调节因子1(SIRT1)是机体内重要的去乙酰化酶,与不同的底物结合可发挥多重作用,例如:抗炎、抗氧化应激与抗凋亡等。有报道显示,视网膜细胞内的CERKL能够通过与SIRT1相互作用,来稳定SIRT1蛋白的表达[8]。最新研究指出,SIRT1可以介导RPE细胞中E2F转录因子1(E2F1)的去乙酰化,而E2F1在RPE细胞抗氧化应激中发挥了重要作用[9]。由此推测,CERKL可能通过调控SIRT1/E2F1轴来保护RPE细胞免受蓝光诱发的氧化应激损伤。本文通过人视网膜色素上皮-19(ARPE-19)细胞体外培养实验探究了CERKL与SIRT1/E2F1轴参与蓝光损伤保护的机制,现将其报告如下。
1材料和方法
1.1材料ARPE-19细胞购自美国ATCC,DMEM/F-12培养基购自美国Gibco公司,RIPA试剂盒、BCA蛋白定量试剂盒、活性氧(ROS)检测试剂盒、脂质氧化检测试剂盒购自上海碧云天生物,CERKL抗体(#ab222828)、SIRT1抗体(#12193)与E2F1抗体(#ab137415)均购自英国Abcam公司,乙酰化的E2F1抗体(乙酰化Lys120,#ABP57453)购自武汉艾美捷科技,Lipofectamine 2000转染试剂购自美国Invitrogen,MTT试剂购自德国罗氏诊断,TUNEL显色法凋亡检测试剂盒购自南京诺唯赞生物,10%甲醛购自上海威奥生物,8-羟基脱氧鸟苷(8-OHdG)检测试剂盒购自上海泽叶生物,山羊血清购自武汉博士德,TritonX-100购自德国Sigma。
1.2方法
1.2.1细胞培养ARPE-19细胞使用含有10%的胎牛血清、100μg/mL链霉素与100U/mL青霉素的DMEM/F-12培养,于5% CO2、37℃的恒温加湿的细胞培养箱(HWS-250B,天津泰斯特公司)中培养,每3d更换一次培养基。
1.2.2蓝光照射将ARPE-19细胞使用胰蛋白酶处理后,以1.5×104个/孔接种于4孔培养板中培养至融合。细胞在接受蓝光暴露前使用磷酸盐缓冲液清洗3次,随后以含有钙、镁和葡萄糖的磷酸盐缓冲液代替培养,使用蓝光(Zensui LED Lamp BlueTM,日本Zensui公司)LED固定于细胞培养层顶部,照射对应时长(0~24h),峰值波长为450nm,光强度为1200lx,对照细胞除了不采用蓝光照射外(黑暗培养),其余操作均相同。
1.2.3细胞形态观察采用相差显微镜(FSX-100,日本Olympus)观察细胞的形态变化,低倍镜(40×)下找到细胞,换用高倍镜(400×)观察,将视野调节清晰后拍照,对照与蓝光照射细胞各选取3个重复进行拍照分析。
1.2.4细胞转染采用Lipofectamine 2000转染试剂将siRNA-NC、siRNA-CERKL、pcDNA3.1、pcDNA3.1-CERKL分别转染至ARPE-19细胞,转染48h后采用PCR法验证转染成功。其中si-CERKL:5’-GAGAAATACCGATGGATGT-3’,siRNA与pcDNA3.1空质粒与重组质粒载体均由广州锐博生物提供。
1.2.5mRNA表达量测定细胞接受对应处理后,使用Trizol法提取细胞的总RNA,紫外分光光度计(NanoDrop 1000,美国赛默飞)测定RNA浓度,采用实时荧光PCR仪(ABI7500,美国ABI)进行PCR扩增,以1μg总RNA作为逆转录PCR模板合成cDNA,以cDNA作为实时荧光PCR模板扩增,以GAPDH作为内参,采用相对定量法(2-ΔΔCt法)计算mRNA表达量,每样本检测3个复孔,计算平均值。
1.2.6蛋白表达量测定采用RIPA试剂裂解细胞,BCA法对蛋白进行定量,取60μg总蛋白进行电泳,转移至PVDF膜,山羊血清封闭1h后,添加一抗(乙酰化的E2F1抗体稀释比例为1∶500,其余抗体为1∶1000)于4℃孵育过夜,取出添加二抗(稀释比例为1∶5000)在室温下孵育1h,ECL显影后进行凝胶成像分析,Image J软件定量分析蛋白的相对表达量,所有处理均设置3个重复,计算平均值。
1.2.7细胞活力测定细胞活力采用MTT法检测,以1×103个/孔的细胞密度接种于96孔板中,蓝光暴露相应时长后取出细胞,加入每孔20μL的MTT试剂,孵育4h,弃上清,加入150μL的DMSO溶解结晶,酶标仪测定490nm处吸光值,所有处理均设置3个重复,计算平均值。
1.2.8细胞凋亡率检测采用TUNEL染色测定细胞凋亡情况,使用10%甲醛固定细胞10min,1×平衡缓冲液孵育10min,使用TUNEL显色法凋亡检测试剂盒,加入制备好的TUNEL反应混合液,于37℃、黑暗条件下反应1h,终止反应后清洗细胞,以链霉素-辣根过氧化物酶覆盖样本,DAB显色,显微镜(Fluo ViewTM FV1000,日本Olympus)下观察TUNEL阳性细胞数,计算TUNEL阳性细胞比例作为凋亡率,所有处理均设置3个重复,计算平均值。
1.2.9免疫荧光染色将ARPE-19细胞使用甲醛固定10min,随后使用0.5% TritonX-100渗透15min,用含10%山羊血清的PBS在室温下封闭1h,一抗(1∶1 000)孵育,4℃过夜,添加二抗(1∶1 000)于37℃孵育1h,DAPI染色5min后,使用激光共聚焦显微镜捕获荧光图像。
1.2.10氧化应激水平的测定采用活性氧检测试剂盒检测细胞内ROS水平,激发和发射波长分别设置为485nm和528nm,使用流式细胞仪(CytoFLEX,美国贝克曼库尔特)分析;采用脂质氧化检测试剂盒测定细胞内丙二醛(MDA)含量,代表脂质过氧化水平;采用8-OHdG检测试剂盒与酶标仪(热电FC,美国赛默飞)测定8-OHdG的含量。所有处理均设置3个重复,计算平均值。
2结果
2.1蓝光照射后ARPE-19细胞形态的变化在正常条件下,ARPE-19细胞始终呈现长轴形状,随着时间推移只存在轻微的变化;而采用蓝光照射后,ARPE-19细胞逐渐收缩变为椭圆形,同时出现空泡(图1),表明蓝光照射对ARPE-19细胞产生了毒性。
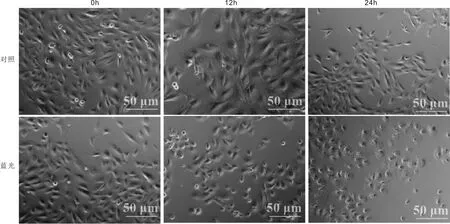
图1 蓝光照射后ARPE-19细胞形态的变化 相差显微镜下观察有无蓝光(BL)照射的ARPE-19细胞形态变化。
2.2蓝光照射对ARPE-19细胞CERKL表达的影响蓝光照射一段时间后,ARPE-19细胞中应激蛋白CERKL的mRNA与蛋白质表达量均呈现增加趋势,与蓝光照射0h比较,蓝光照射6h后CERKL的mRNA与蛋白质达到高峰(t=18.980、14.470,均P<0.001),12h与24h仍维持在较高水平(mRNA:t=10.920、4.021,P<0.001、=0.016;蛋白质:t=13.690、6.682,P<0.001、=0.003),见图2。因此后续实验中蓝光暴露时长设置为24h。
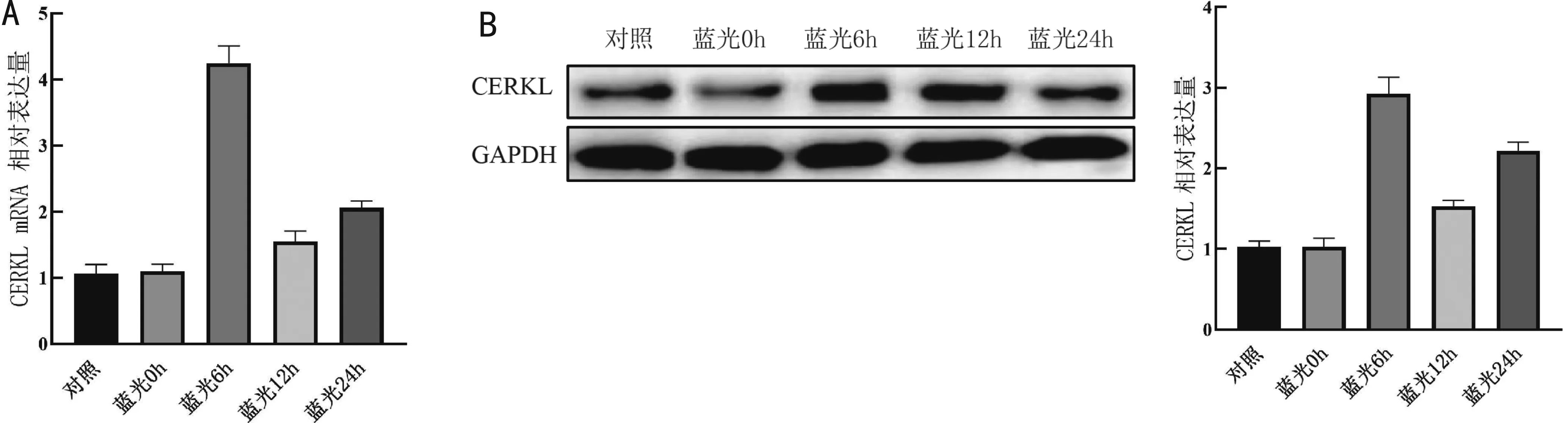
图2 蓝光照射对ARPE-19细胞CERKL表达的影响 A:蓝光照射24h内细胞CERKL mRNA表达的变化;B:蓝光照射24h内细胞CERKL蛋白质表达的变化。
2.3CERKL表达对蓝光照射导致的ARPE-19细胞氧化应激损伤的影响与对照细胞比较,蓝光照射24h后细胞CERKL表达量升高(t=10.890,P<0.05,图3A),相对细胞活力随着时间推移逐渐降低(F=69.550,P<0.001,图3B),凋亡率升高(t=15.610,P<0.001,图3C),ROS、MDA、8-OHdG含量升高(t=20.590、17.940、22.130,均P<0.001,图3D~F);在蓝光照射的基础上,采用siRNA-CERKL抑制CERKL表达会加剧这种变化,表现为6、12、24h细胞相对活力的降低(t=6.286、6.609、5.826,P=0.003、0.003、0.004,图3B),凋亡率升高(t=6.940,P=0.002,图3C),ROS、MDA、8-OHdG含量升高(t=13.610、19.770、18.350,均P<0.001,图3D~F);而在蓝光照射的基础上,采用pcDNA3.1-CERKL上调CERKL表达则能逆转此变化,表现为6、12、24h细胞相对活力的恢复(t=3.836、8.385、8.980,P=0.019、0.001、<0.001,图3B),凋亡率降低(t=9.584,P<0.001,图3C),ROS、MDA、8-OHdG含量降低(t=8.485、10.640、22.020,P=0.001、<0.001、<0.001,图3D~F)。上述结果提示,上调CERKL能缓解蓝光照射导致的ARPE-19细胞氧化应激损伤,提高细胞存活率。
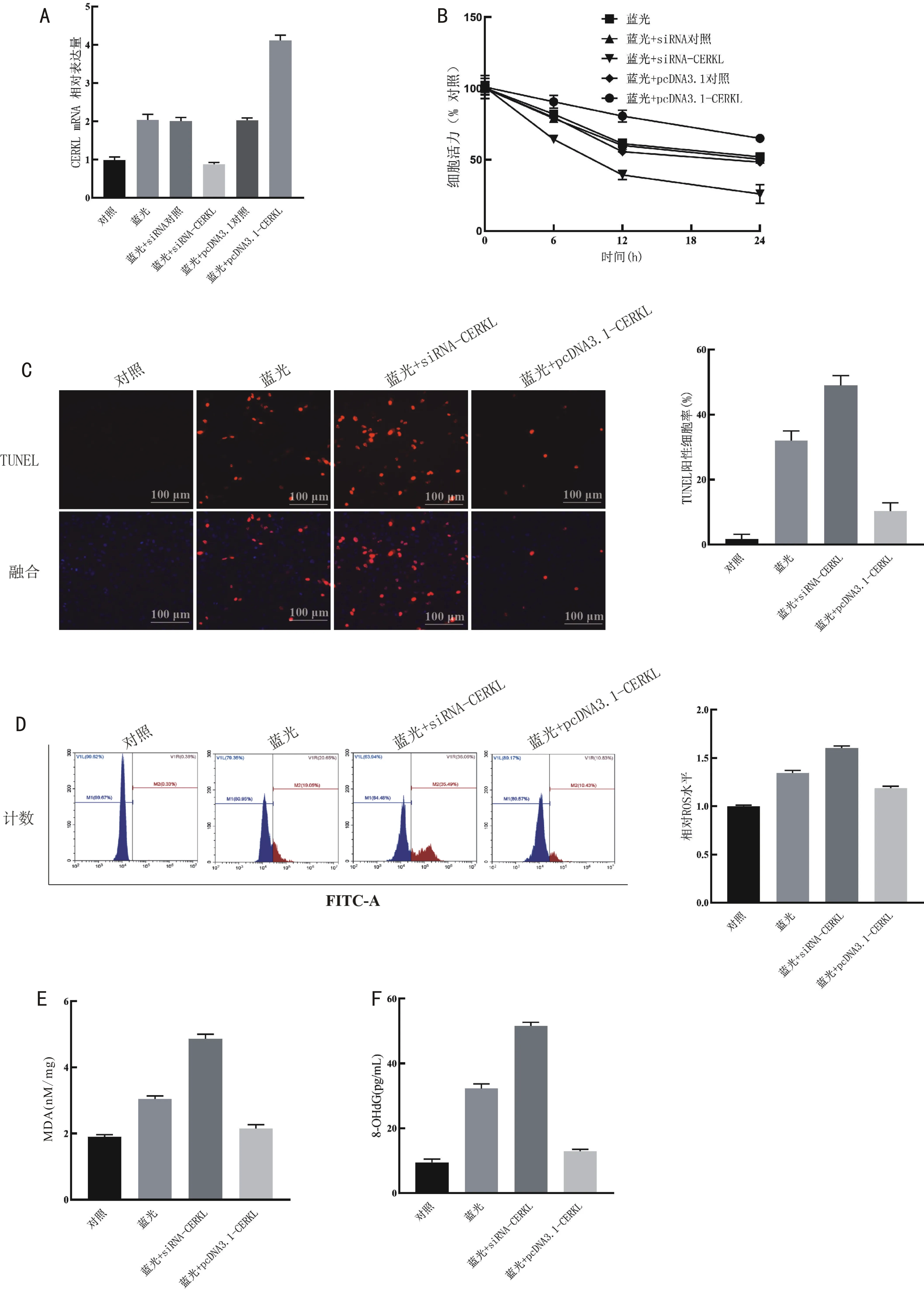
图3 CERKL表达对蓝光照射导致的ARPE-19细胞氧化应激损伤的影响 A:各组CERKL mRNA表达;B:MTT法测定细胞活力;C:TUNEL染色检测细胞凋亡;D:流式细胞仪测定细胞中ROS含量;E:各组MDA含量;F:各组8-OHdG含量。
2.4CERKL与SIRT1/E2F1轴的关系免疫荧光染色分析显示,CERKL与SIRT1在ARPE-19细胞的细胞质和细胞核中共同定位(图4A);蓝光照射增加了SIRT1表达(t=12.810,P<0.001),诱导E2F1的脱乙酰化(t=3.252,P=0.031),抑制CERKL减少了蓝光照射下细胞的SIRT1表达(t=14.170,P<0.001),并增强了E2F1乙酰化程度(t=10.190,P<0.001),而上调CERKL又可增强蓝光照射下细胞的SIRT1表达(t=18.060,P<0.001),促进E2F1的脱乙酰化(t=6.383,P=0.003),见图4B~D。以上提示,上调CERKL能激活SIRT1/E2F1轴。
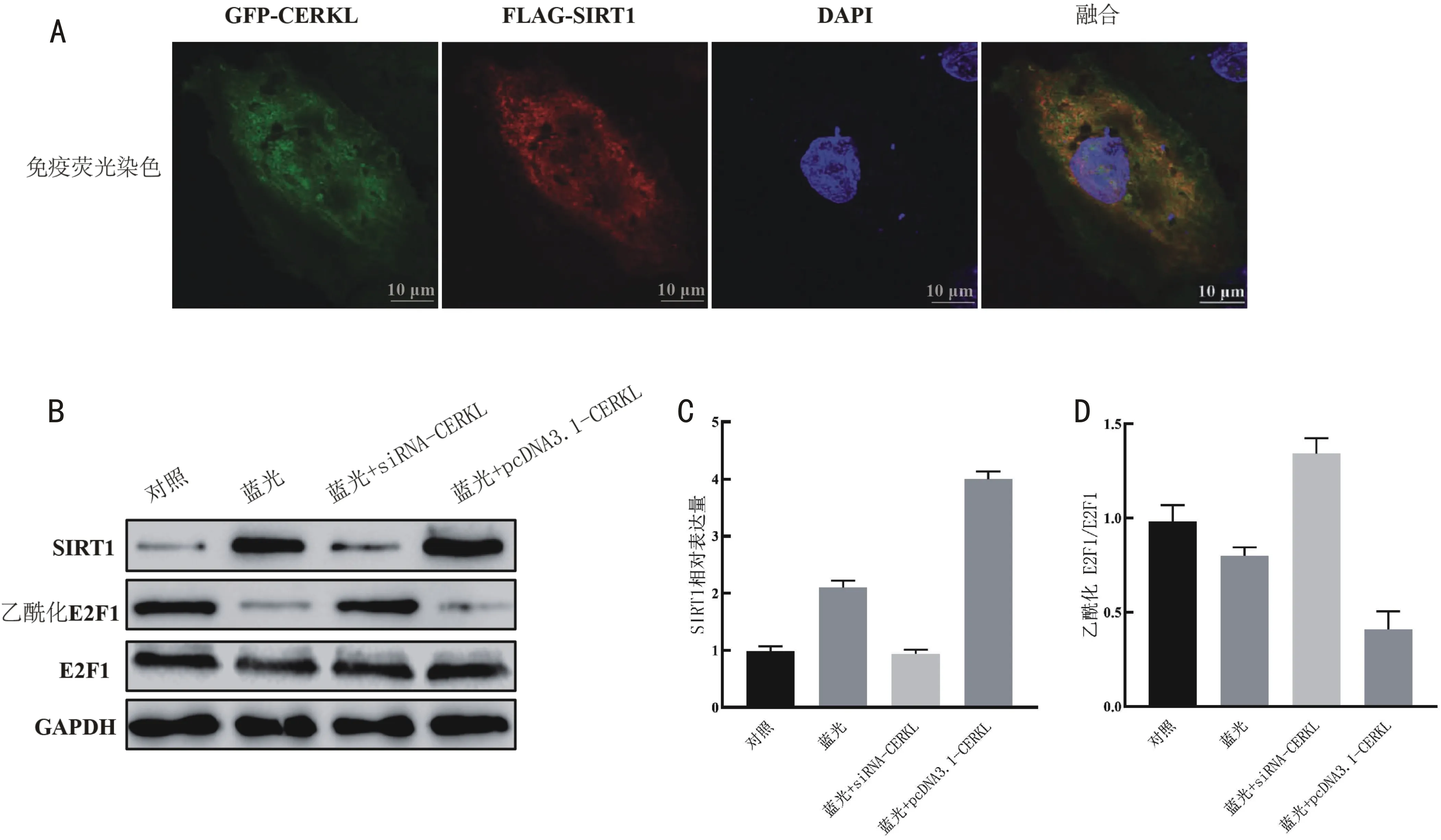
图4 CERKL与SIRT1/E2F1轴的关系 A:免疫荧光染色对ARPE-19细胞中CERKL与SIRT1进行定位;B:各组SIRT1、乙酰化E2F1与E2F1蛋白表达情况;C:各组SIRT1表达情况;D:各组乙酰化E2F1/E2F1表达情况。
2.5沉默SIRT1对CERKL缓解蓝光诱发ARPE-19细胞氧化应激损伤作用的影响为证实CERKL是否通过调控SIRT1轴对蓝光照射下的ARPE-19细胞产生保护作用,分别将pcDNA3.1-CERKL与siRNA-SIRT1转染ARPE-19细胞,蓝光照射24h。转染pcDNA3.1-CERKL上调CERKL表达(蓝光vs蓝光+pc-CERKL:t=15.780,P<0.001),采用siRNA-SIRT1沉默了SIRT1的表达(蓝光vs蓝光+si-SIRT1:t=16.560,P<0.001,蓝光+pc-CERKLvs蓝光+pc-CERKL+si-SIRT1:t=22.040,P<0.001),但不影响CERKL的表达量(蓝光+pc-CERKLvs蓝光+pc-CERKL+si-SIRT1:t=0.389,P=0.717),见图5A,表明转染成功。结果显示,单独沉默SIRT1或在上调CERKL表达的基础上沉默SIRT1,均会导致6、12、24h的细胞活力降低(蓝光vs蓝光+si-SIRT1:t=6.815、8.578、5.362,P=0.002、0.001、0.006;蓝光+pc-CERKLvs蓝光+pc-CERKL+si-SIRT1:t=3.836、8.385、9.329,P=0.019、0.001、<0.001,图5B),细胞凋亡率升高(蓝光vs蓝光+si-SIRT1:t=6.940,P=0.002,蓝光+pc-CERKLvs蓝光+pc-CERKL+si-SIRT1:t=11.680,P<0.001),见图5C,ROS、MDA、8-OHdG含量升高(蓝光vs蓝光+si-SIRT1:t=17.150、27.830、23.520,均P<0.001;蓝光+pc-CERKLvs蓝光+pc-CERKL+si-SIRT1:t=29.520、10.620、59.870,均P<0.001),见图5D~F。以上提示,CERKL通过激活SIRT1轴减轻蓝光诱导的ARPE-19细胞氧化应激损伤。

图5 沉默SIRT1对CERKL缓解蓝光诱发ARPE-19细胞氧化应激损伤作用的影响 A:各组CERKL、SIRT1 mRNA表达;B:MTT法测定细胞活力;C:TUNEL染色检测细胞凋亡;D:流式细胞仪测定细胞中ROS含量;E:各组MDA含量;F:各组8-OHdG含量。
3讨论
蓝光造成的视网膜损伤是当前眼科研究的热点之一。眼睛长时间暴露于各类可见光中,其中以波长在400~450nm的蓝光对眼睛危害最大[10-11]。本研究探讨了CERKL/SIRT1/E2F1分子轴参与蓝光诱发的ARPE-19细胞氧化应激损伤的机制,以期为蓝光损伤干预提供潜在的分子靶点。结果发现,蓝光照射后的ARPE-19细胞逐渐收缩成圆球状,且出现空泡,表明蓝光对细胞产生了毒害作用。Takayama等[12]研究发现,蓝光暴露会导致ROS的增加,并通过氧化应激导致RPE细胞凋亡,并指出这可能是导致视网膜结构损伤的重要途径。本研究观察到蓝光照射后ARPE-19细胞活力降低,凋亡率升高,ROS、MDA、8-OHdG的含量均显著升高。ROS的过度积累会造成细胞内的氧化应激损伤、脂质过氧化物MDA与DNA损伤标志物8-OHdG释放增多,最终导致细胞死亡[13]。
CERKL在眼部视网膜组织中大量表达,并被报道为视网膜色素病变的致病基因[14]。多国研究发现,CERKL的突变会导致视网膜色素性病变[5,15-16]。本研究中发现,在蓝光照射后的不同时间点,CERKL表达水平均有不同程度的升高。随后通过质粒转染调节ARPE-19细胞内CERKL表达,结果显示沉默CERKL会加剧蓝光诱发的细胞氧化应激损伤与凋亡,而上调CERKL则能缓解蓝光损害,表明CERKL能抑制ARPE-19细胞内的氧化应激与凋亡,提高细胞存活率。
SIRT1是细胞内关键的氧化还原调节因子,而E2F1是重要的转录激活因子,SIRT1可以通过结合E2F1抑制其诱凋亡的活性,同时通过使E2F1脱乙酰化促进细胞内的抗氧化因子转录,清除细胞内因蓝光暴露积累的ROS,保护细胞免受ROS诱发的凋亡,提高细胞的存活率[17-18]。本研究结果证实:上调CERKL增加了SIRT1表达,促进了E2F1脱乙酰化,沉默SIRT1能逆转CERKL缓解蓝光诱导的ARPE-19细胞氧化应激损伤。Li等[19]研究指出CERKL可以通过与线粒体中的硫氧化还原蛋白2的相互作用,减少ROS诱发的氧化应激与凋亡。García-Arroyo等[4]研究证明CERKL可以通过调控线粒体大小与形状,减轻RPE细胞内的氧化应激。与上述研究共通之处是,本研究中发现的CERKL下游靶点SIRT1也广泛存在于线粒体中,因此CERKL也是通过与线粒体蛋白的作用来清除ROS;不同的是,本研究中揭示了CERKL激活SIRT1/E2F1轴,促进抗氧化因子转录的信号传导机制,丰富了对CERKL参与调控视网膜细胞氧化应激机制的认识,说明了CERKL保护视网膜RPE细胞的机制是复杂且多通路的。
综上所述,CERKL可以通过激活SIRT1蛋白表达,促进E2F1的脱乙酰化,从而减轻蓝光诱导的ARPE-19细胞氧化应激损伤,随着日后研究的深入,CERKL/SIRT1/E2F1轴可能成为视网膜光损伤的干预靶点。
