长柄扁桃在乌兰布和沙地的生理适应性研究
2022-08-06罗竹梅刘冬林
任 俊,杨 涛,樊 强,罗竹梅*,吕 刚,刘冬林,吴 彪
(1. 巴彦淖尔市沙漠综合治理中心,内蒙古 巴彦淖尔市 015000;2.陕西省林业科学院,西安 710082;3.国家林业和草原局长柄扁桃工程技术研究中心,陕西 榆林 719000;4.长柄扁桃国家创新联盟,陕西 榆林 719000)
长柄扁桃(Amygdaluspedunculata)是蔷薇科(Rosaceae)桃属(Amygdalus)落叶灌木,俗称野樱桃或毛樱桃,广泛分布于陕西北部及内蒙古沙地,是当地乡土树种,目前被列为国家二级濒危珍稀保护植物[1]。长柄扁桃根系发达,萌蘖力强,枝条稠密,耐旱耐寒、耐沙埋,抗逆性强,是中国西北部干旱和半干旱地区沙地造林、水土保持和沙漠治理的优势树种和先锋树种[2-3],同时也是一种难得的木本油料树种、优良药用植物和绿化观赏植物。随着国家防沙治沙工作力度的进一步加强,挖掘和开发适应西部沙地干旱环境且具有较高经济价值的乡土树种就成为沙地造林的主要任务。干旱少雨是大规模沙地造林的最主要限制因子,选择和推广适应干旱环境的乡土树种尤为重要。长柄扁桃长期都处于野生状态,近十年来,其保护、人工栽培、开发利用和基地建设都有了长足发展,取得了许多重要研究成果[4,6]。不同植物在干旱环境下会表现出不同的生理特征,以抵抗恶劣环境维持正常的生命活动[5]。以乌兰布和沙地为环境背景,研究在乌兰布和沙地干旱条件下长柄扁桃生理指标的变化,进而研究其对土壤干旱和气候干旱的反应和适应性,探索长柄扁桃的抗旱阈值,为在乌兰布和及其相似沙地开展大面积长柄扁桃造林具有重要的理论价值。
本研究以长柄扁桃为对象,人工模拟沙地土壤干旱梯度,测定在不同干旱梯度下长柄扁桃的生理指标变化,并进一步探讨长柄扁桃对乌兰布和沙地土壤干旱环境的生理适应性,以期掌握长柄扁桃苗期生长的对水分亏缺的反应,给大面积人工造林水分管理提供参考。
1 研究区概况
乌兰布和沙漠是中国八大沙漠之一,地处内蒙古自治区西部巴彦淖尔市和阿拉善盟境内。试验区位于内蒙古巴彦淖尔市磴口县沙金套海苏木巴音温都尔嘎查,穿沙公路36 km处的万亩长柄扁桃种植基地,乌兰布和沙漠东部边缘,东经106°9′~ 107°10′,北纬40°9′~ 40°57′。磴口县万亩长柄扁桃种植基地总面积2 470 hm2,水、电、路等基础设施齐全,灌溉水源充足,交通便利。
试验地位于万亩长柄扁桃种植基地中心地段,地表为沙丘和沙生植物覆盖,土壤为风沙土,地势平坦,土地肥沃,灌溉便利。温带大陆性季风气候,冬季寒冷漫长,春秋短暂,夏季炎热,降雨量少,日照充足,热量丰富,昼夜温差大,积温高,无霜期短。年均气温为7.6 ℃,极端最高气温38.2 ℃,极端最低气温-34.2 ℃。植物生长期(5-9月)的光合有效辐射40.19 kcal·cm-2,积温约为3 100 ℃,昼夜温差14.5 ℃。年均降水量144.5 mm,年均蒸发量2 397.6 mm。年均风速4.5 m·s-1,≥3 m·s-1的时间达6 800 h以上,≥6 m·s-1风速的时间2 200~2 500 h以上。植物主要有甘草(Glycyrrhizauralensis)、花棒(Hedysarumscoparium)、麻黄(Ephedrasinica)、肉苁蓉(Cistanchedeserticola)、锁阳(Cynomoriumsongaricum)、沙棘(Hippophaerhamnoides)、梭梭(Haloxylonammodendron)等名贵药材和稀有种,并有大小湖泊200多处。
2 材料与方法
本研究采用盆栽试验方法,选取1 a生长柄扁桃幼苗,在具有可移动遮雨棚设施的试验地露天培养,晴天时遮雨棚开放,阴雨天时遮雨棚关闭以防止雨水渗入。试验盆口径25 cm,盆高30 cm。盆栽土壤采自于试验区的壤土和砂土,土壤采回后先风干然后过筛(筛孔直径2 mm),按1∶1配比成盆栽土。填装土壤后向各盆中加水至饱和,静置2 d后,测定盆栽土物理指标,盆栽土的田间持水量208.52 g·kg-1,容重1.52 g·cm-3,饱和导水率10.84 cm·h-1,总有机碳2.66 g·kg-1。选择长势一致的长柄扁桃幼苗移栽至盆中,待植株成活且长势稳定后,每盆定苗3株,盆表面均匀覆盖直经0.5~1.0 cm的砾石,抑制水分蒸发。共设置4个干旱梯度:(1)对照(CK),盆栽土壤含水量保持为田间持水量75%~80%;(2)轻度干旱(T1),盆栽土壤含水量保持为田间持水量55%~60%;(3)中度干旱(T2),盆栽土壤含水量保持为田间持水量35%~40%;(3)极度干旱(T3),盆栽土壤含水量保持为田间持水量15%~20%。
每个处理设置3次重复,水分控制采取人工称重的方法,即每天下午16∶00称重并补充散失的水分,分别在处理后第7、14、21、28 d采集长柄扁桃叶片进行相关测定。参照《植物生理学实验技术》[7]的方法测定各项生理指标,采用硫代巴比妥酸法测定丙二醛(malondialdehyde,MDA)含量;氮蓝四唑光还原法测定超氧化物歧化酶(superoxide dismutase,SOD)活性;愈创木酚法测定过氧化物酶(peroxidase,POD)的活性;用茚三酮显色法测定游离脯氨酸(Pro)含量。采用Excel 2013进行数据处理及统计分析。
3 结果与分析
3.1 不同干旱条件下长柄扁桃MDA含量的变化
丙二醛(MDA)是细胞内膜脂过氧化作用的最终产物之一,它的产生会加剧膜的损伤,细胞膜系统受损后导致质膜透性增大,内容物渗漏,进而阻碍植物的正常生长。所以,经常用细胞内MDA的含量间接判断细胞膜受伤害的程度[8]。MDA含量升高时细胞膜系统受伤害程度加剧;MDA含量下降时细胞膜系统受伤害程度减缓。
从图1可分析得出,长柄扁桃叶片MDA含量随着干旱程度的加剧而增加,且随着干旱时长的延长呈上升趋势。在极轻度干旱(CK)条件下,MDA含量上升缓慢,上升幅度只有1.11 mmol·g-1,随着干旱程度的增加,MDA含量随干旱时长不断提升,提升幅度最大的时段在14 d到21 d之间,21 d后上升幅度有所减缓。在干旱开始的7 d内,轻度干旱与CK的MDA含量差值极小,而中度干旱和极度干旱的的MDA含量则大幅提升,与CK的差值显著。MDA含量变化的趋势间接反映出了长柄扁桃叶对轻度干旱的适应能力。
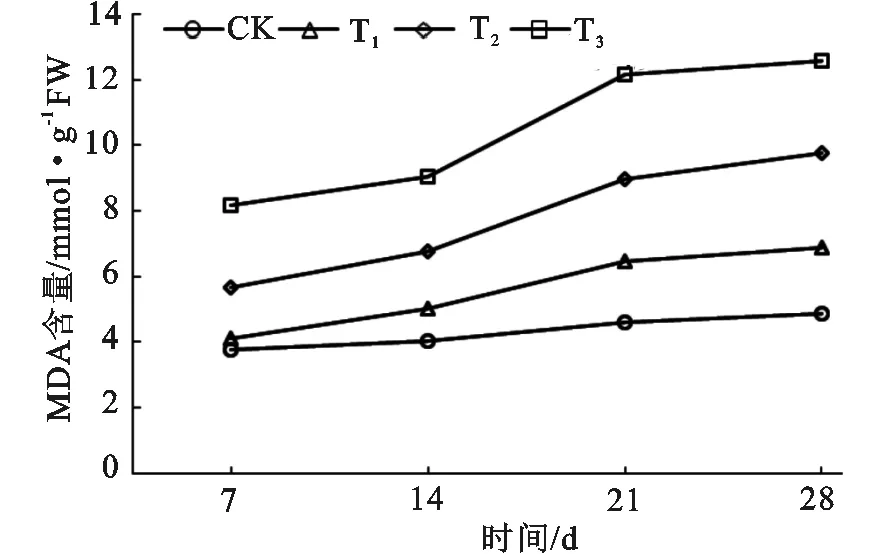
图1 长柄扁桃MDA含量随干旱程度和干旱时长的变化
3.2 不同干旱条件下长柄扁桃SOD活性的变化
超氧化物歧化酶(SOD)是植物活性氧代谢的关键酶,植物对逆境的适应能力与SOD的活性密切相关,它可以防止自由氧基对细胞膜和生物大分子的毒害,维持植物的正常生理代谢[8]。
从图2分析得出,长柄扁桃的SOD活性随着干旱程度的增加而提升,在每个测定节点上,干旱程度越高,SOD活性越高。在极轻度干旱(CK)和轻度干旱(T1)情况下,随着干旱间的延长,SOD活性随然有所提升,但提升的幅度都较小,分别为46.02 U·g-1·min-1和170.06 U·g-1·min-1;在中度干旱(T2)和极度干旱(T3)时,SOD活性急剧提升,至21 d达到峰值,分别为786.35 U·g-1·min-1和1 136.52 U·g-1·min-1,升幅分别达255.07 U·g-1·min-1和403.87 U·g-1·min-1,之后出现小幅回落。在中度干旱和极度干旱条件下,SOD活性值在21 d后出现的小幅回落,表明长柄扁桃对短期适度干旱有一定的适应能力;但在长时间重度干旱胁迫下,这种适应能力有所下降,需要及时补充水分。
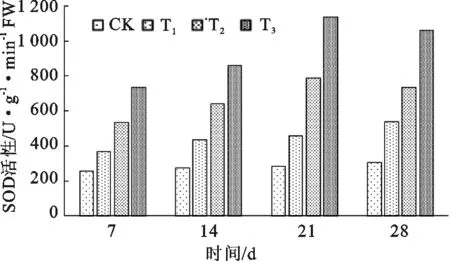
图2 长柄扁桃SOD活性随干旱程度和干旱时长的变化
3.3 不同干旱条件下长柄扁桃POD活性的变化
过氧化物酶(POD)也是一种保护酶,广泛存在于植物细胞内,有利于清除自由基,能降低膜脂过氧化水平,从而减轻膜的伤害程度[8]。长柄扁桃POD活性的变化趋势与SOD活性变化不同,从极轻度干旱到中度干旱,干旱程度越高,POD活性越大,但到了极度干旱时,POD活性稍有回落,反而低于中度干旱的POD水平。从干旱时长来看,都是随着干旱时间的延长POD活性不断提升,只是在极度干旱情况下,当干旱持续到21 d后,POD活性基本维持在同一水平上,没有显著变化(图3)。这一结果间接反映出长柄扁桃对干旱程度适应范围,在极度干旱情况下,长柄扁桃的抗旱能力有所减弱。

图3 长柄扁桃POD活性随干旱程度和干旱时长的变化
3.4 不同干旱条件下长柄扁桃游离脯氨酸(Pro)含量的变化
脯氨酸( Pro) 作为植物细胞内的渗透调节物质,可保持胞质溶液与外界环境的渗透平衡,对原生质起保护和维持保水平衡的作用。当长柄扁桃在受到干旱胁迫之后,脯氨酸( Pro)含量有一个敏感性应对反应,表现出随干旱程度和干旱时长的变化特征(图4)。
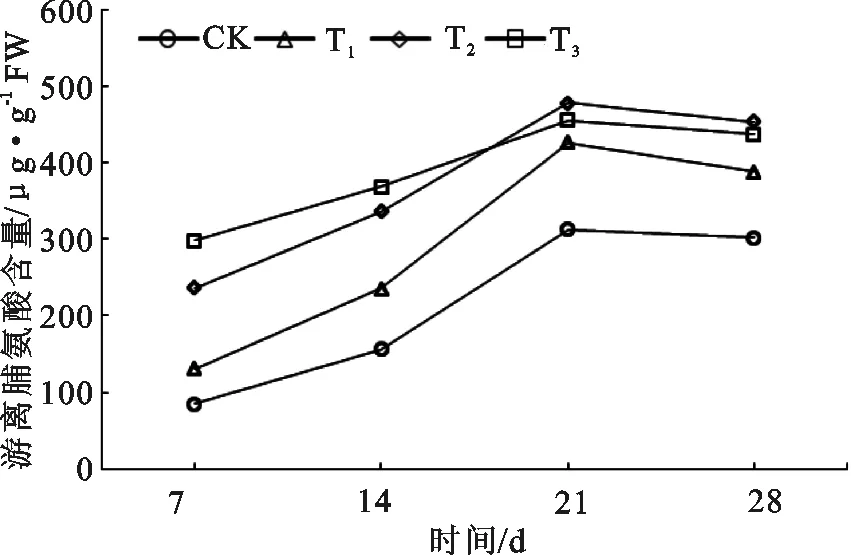
图4 长柄扁桃游离脯氨酸(Pro)含量的变化
从图4可看出,随着干旱程度的加剧,脯氨酸(Pro)含量也随之升高,但在极度干旱条件下,在干旱胁迫的最初阶段,脯氨酸(Pro)含量仍然处于较高水平,当干旱时长达到21 d之后,脯氨酸(Pro)含量反而不及中度干旱的水平。在4个干旱程度水平上,脯氨酸(Pro)含量都呈现出先升后降的趋势,在胁迫21 d时长柄扁桃的Pro含量出现了一个高峰,但在持续干旱28 d后都呈下降趋势,说明其对短期干旱的响应比较敏感,能尽快产生Pro调节细胞质溶液与环境的渗透平衡,但持续性干旱之后,则超过了Pro的调节能力。
4 结论与讨论
长期生长在干旱条件下的植物在长期的进化过程中都形成了一系列生理适应能力,从而能适应干旱环境而得以生存。长柄扁桃是乌兰布和沙地乡土落叶灌木,在长期的生长适应过程中形成了特殊的生理结构和生理特性,比如叶片表面厚的蜡质层,庞大的根系,特殊的气孔结构[8-9]。本试验通过人工模拟干旱胁迫处理,通过测定长柄扁桃幼苗在水分胁迫下随着胁迫时间的延长几个生理指标的变化,间接揭示了长柄扁桃抗旱的原因以及对干旱环境的适应。
研究结果表明:长柄扁桃叶片MDA含量、SOD活性、POD活性、Pro含量都随着干旱程度的加剧和干旱时间的延长而表现出了相应的变化规律。MDA含量随着干旱程度的加剧而增加,且随着干旱时长的延长呈上升趋势;SOD活性也随着干旱程度的增加而提升,但在中度干旱和极度干旱时,当干旱时长达到21 d后,则出现小幅回落的现象;POD活性的变化趋势与 SOD活性变化不同,在极度干旱时,POD活性低于中度干旱的水平,而且当干旱持续到21 d后,POD活性基本维持在同一水平上; Pro含量整体上都呈现出先升后降的趋势,在胁迫21 d时长柄扁桃Pro含量出现了一个高峰,但在持续干旱28 d后都呈下降趋势,当干旱时长达到21 d之后, 极度干旱下Pro含量反不及中度干旱的水平。这些生理指标的变化特征间接反映出长柄扁桃干旱条件的适应特征,在干旱程度不断加剧的情况下,各生理指标对干旱的适应存在阈值,在短期干旱时响应比较敏感,但持续性干旱之后,其敏感度和调节能力都有所下降。
长柄扁桃具有典型抗旱植物的生理特征,对干旱具有一定的适应能力,但在生产中必须重视长柄扁桃抗旱的生理极限,在有条件的情况下尽可能在生理功能正常的情况下及时补充水分以保障其正常生长发育。
