一株喜油嗜热芽孢杆菌G1201 产高温蛋白酶的性质研究及异源表达初探
2022-08-06刘国瑞张军汤伟孙晓雯唐涛赵丽红何增国
朱 檬,刘国瑞 ,张军 ,汤伟,2,孙晓雯,唐涛,2,赵丽红,何增国
(1.中国海洋大学医药学院,山东青岛 266000;2.青岛海洋生物医药研究院,山东青岛 266000;3.青岛百奥安泰生物科技有限公司,山东青岛 266000)
蛋白酶是一种重要的酶制剂,在饲料、食品、医药等领域有着广泛的应用(Goutam 和Arun,2017)。有统计显示,在酶制剂市场份额中蛋白酶占比超过65%(Garg等,2016)。随着人们对产品品质追求的提高和生产分工的细化,普通蛋白酶逐渐无法满足如饲料制粒、食品烘焙中的高温过程,这些行业对高温蛋白酶提出了迫切需求。
微生物以其来源广泛、培养周期短、便于工程化操作成为目前蛋白酶最主要的来源(于建荣等,2015;邓菊云,2008;张燕新等,2007)。更重要的是许多极端自然环境中,如火山口、海底热液区、热泉等天然存在许多耐热的微生物菌种,其中必然孕育了高温蛋白酶的基因资源。对其中微生物种质资源的发掘,可为高温蛋白酶的开发提供筛选基础。已有研究报道,从这些环境中筛选得到了具有良好应用前景的高温蛋白酶(桑鹏等,2019;桑鹏等,2019;陈逍遥,2015;Mashayekhi等,2012)。
目前已报道产高温蛋白酶的菌属有芽孢杆菌属、热球菌属、硫化叶菌、火球菌属等(曾静,2014;吕明生等,2011;闻真珍,2010;黄光荣等,2007)。Geobacillus 属是2001 年被命名的一个耐热菌属,该属微生物来源广泛,具有良好的耐热性(Nazina等,2001)。已有一些从该属微生物中分离得到高温蛋白酶的报道(Goutam 和Arun,2017;廖艳江,2010;Chen等,2004;Nazina,2001)。本文从长期处于高温的烧烤环境中分离得到1 株喜油嗜热芽孢杆菌(Geobacillus thermoleovorans),发现其产高温蛋白酶的功能,并对其产高温蛋白酶进行了表征和酶学性质研究。同时,本文通过基因工程菌手段,尝试对该高温蛋白酶进行异源表达。
1 材料与方法
1.1 材料
1.1.1 主要试剂 T4 DNA Ligase、胶回收试剂盒、IPTG 和硫酸卡那霉素购自上海生工生物工程有限公司;限制性内切酶(EcoR Ⅰ、Xho Ⅰ)购自大连宝生物有限公司;基因组提取试剂盒、质粒小提试剂盒购自北京天根生化科技有限公司;酪氨酸、福林酚购自Sigma;其余非特殊注明试剂均购自国药化学试剂有限公司。
菌株与质粒:大肠杆菌BL21(DE3)和质粒pET-28a(+)由中国海洋大学海洋生命学院惠赠。
1.1.2 培养基及缓冲溶液、试剂 脱脂奶粉筛选培养基参照Thebti(2016)方法并做出改良:脱脂奶粉20 g,琼脂20 g,加蒸馏水至1 L,115 ℃,灭菌20 min。
酪蛋白标准溶液:称取0.500 g 酪蛋白,用少量0.5 moL 的NaOH 溶液润湿,然后加入缓冲液大约40 mL,沸水浴30 min,期间搅拌至完全溶解,冷却后定容至50 mL。
1.1.3 样品来源 土壤样品采集自中国海洋大学浮山校区附近(36.075422°N,120.437054°E)蒸汽排气口下含油土样。
1.2 方法
1.2.1 产高温蛋白酶菌株筛选及理化特征鉴定取采集土样25 g 加入到225 mL 无菌水中,180 r/min摇培30 min,制成样品悬液。取1 mL 样品悬液加入到9 mL 无菌水中进行10 倍梯度稀释。选取合适的稀释梯度,取100 μL 涂布于脱脂奶粉培养基上,65 ℃培养24 h。观察比较水解圈直径与菌落直径比值,选取比值最大的进行液体复筛,70 ℃下测量发酵液酶活,筛选出酶活最高的菌株,编号G1201。对筛选得到的菌株在LB 固体培养基上进行连续3 次划线分纯,甘油管保藏纯化出的单菌落以待进一步研究。
参照《常见细菌系统鉴定手册》对G1201 进行厌氧性、运行性、触酶等生理性质鉴定,并利用O’Meara 法进行V-P 实验鉴定其生化特征。同时,进行革兰氏染色,并用透射电镜(TEM)观察G1201 的菌体形态特征。
1.2.2 16S rRNA 基因序列分析及系统发育构建采用基因组提取试剂盒提取G1201 基因组DNA。以此为模板,采用通用引物 27 F(5'-AGAGTTTGATCCTGGCTCAG-3')和1492 R(5'-GGTTACCTTGTTACGACTT-3')扩 增G1201 的16S rRNA 基因序列。PCR 反应条件(张军,2110),95 ℃55 min;94 ℃0.5 min,55~ 60 ℃0.5 min,72 ℃1 min,35 个循环;72 ℃10 min。PCR 产物送上海生工测序。测序结果在GeneBank 中进行Blast N 比对,用MEGA7.0 软件,采用Neighbor-Joining 法构建系统发育树,进化树分支采用Bootstrap 法并重复1000 次。
1.2.3 粗酶的制备及酶活测定 将G1201 菌株接入LB 培养基中,65 ℃,180 r/min 培养16 h,离心取上清液。在4 ℃条件下,分别用30%、40%、50%、60%、70%和80%硫酸铵对发酵上清液逐级沉淀过夜。之后,4 ℃,10000 r/min 离心30 min,收集沉淀物,将沉淀分别溶于相同体积的PBS(pH 7.5)中测定酶活,以酶活最大的硫酸铵沉淀物为粗酶,进行酶学性质分析。
酶活力单位定义为:在70 ℃条件下,每毫升酶液每分钟水解酪蛋白产生1 μg 酪氨酸,定义为一个酶活力单位,以U/mL 表示。酪氨酸标准曲线绘制及酶活测定参照国标GB/T23527-2009(中国国家标准化管理委员会,2009),酶反应温度设为70 ℃。
1.2.4 粗酶的酶学性质分析 粗酶溶于PBS中,进行最适催化温度和温度耐受性测定。将粗酶液与酪蛋白溶液在30~ 100 ℃条件下反应后,于碱性环境中与福林酚溶液进行显色反应,在680 nm处测量其吸光度,确定最适酶催化温度。温度耐受性测定时,将粗酶液放置30~ 100 ℃下水浴孵育1 h后,在最适温度70 ℃下与酪蛋白反应,测定不同温度处理后,对酶活的影响。
粗酶和酪蛋白分别溶于不同pH(pH 4.0~12.0)的缓冲液中,制成粗酶液和酪蛋白标准溶液,在70 ℃下反应,通过测定不同pH 条件下粗酶液与酪蛋白反应的酶活,确定最适催化pH。pH耐受性测定,将不同pH 缓冲液溶解的粗酶液水浴孵育1 h,在70 ℃,pH 9.0 条件下测定酶活,确定其pH 耐受性。
粗酶溶于pH 9.0 的缓冲液,分别加入终浓度5 mmol/L 的Ca2+、Mg2+、Co2+、Cu2+、Fe2+、Mn2+、Zn2+和Na+,水浴孵育1 h,在70 ℃,pH 9.0 条件下测定酶活,探究不同金属离子对酶活的影响。
在上述粗酶液中分别添加1%的表面活性剂(SDS、吐温20、吐温80 和曲拉通X-100)和终浓度5 mmol/L 的蛋白酶抑制剂[DTNB(2-硝基苯甲酸)、PMSF(苯甲基磺酰氟)和EDTA(乙二胺四乙酸)。水浴孵育1 h后,在70 ℃,pH 9.0 条件下测定酶活,分析表面活性剂和蛋白酶抑制剂对酶活的影响。
1.2.5 高温蛋白酶的异源表达及分离纯化
1.2.5.1 G1201 蛋白基因克隆与表达载体构建以NCBI 数据库中Geobacillus 属微生物编码丝氨酸蛋白酶基因(ser)的序列为基础,使用DNAMAN软件进行多重序列比对,用Primer 5 软件在ser 基因保守区设计上下游引物FE(5'-CGGAATTCATGTTTGGC TATTCAATAGTGCAGTTAGCCC-3')和RX(5'-CGCTCGAGCTAGCGCTGCAGCAGTTGCT CAAT-3'),下划线分别为EcoR Ⅰ和Xho Ⅰ的酶切位点。以G1201 的基因组DNA 为模板,扩增ser 的基因序列,之后分别用限制性内切酶EcoRⅠ和Xho Ⅰ对扩增片段和质粒载体pET-28a 双酶切。T4 连接酶连接过夜后,热激转化大肠杆菌BL21(DE3)中,用含有硫酸卡纳霉素(终浓度50 μg/mL)的平板、菌落PCR 和质粒单双酶切筛选阳性转化子并送上海生工测序。
1.2.5.2 重组蛋白诱导表达及分离纯化 挑取BL21-pET-28a-ser 单菌落于含50 μg/mL 硫酸卡那霉素的LB 培养基中,37 ℃,180 r/min 培养3 h,至OD600到0.4~ 0.6。之后,加入IPTG(终浓度0.4 mmol/L)诱导表达4 h。4 ℃,8000 r/min 离心10 min 收集菌体,用PBS 洗涤2 次。离心后,加破碎缓冲液重新悬浮菌体进行超声破壁(功率750 W)。超声破碎后离心,分别取上清液和沉淀,进行SDS-PAGE 检测,发现目的条带以包涵体的形式存在于沉淀中。用8 mol/L 尿素溶解包涵体蛋白,低温搅拌洗涤30 min。4 ℃,12000 r/min 离心20 min,取上清过Ni 柱纯化目的蛋白并用蛋白测试试剂盒测定纯化蛋白酶的含量。
2 结果与分析
2.1 菌株的分离与鉴定
2.1.1 产高温蛋白酶菌株筛选结果 通过在脱脂奶粉筛选培养基上对样品初筛,在65 ℃培养条件下得到14 株产生透明圈的菌株,挑选透明圈直径和菌落直径比值最大的菌株(图1a),进行液体复筛,筛选出酶活最高的菌株,编号G1201。在LB平板上70 ℃下培养24 h后,G1201 菌落直径约3~ 4 mm,呈微黄色、半透明、边缘不规则。该菌株革兰氏染色呈阴性,通过透射电镜观察,该菌株呈杆状,长3.0~ 3.5 μm,宽0.8~ 1.2 μm,端生孢子,有鞭毛(图1b)。
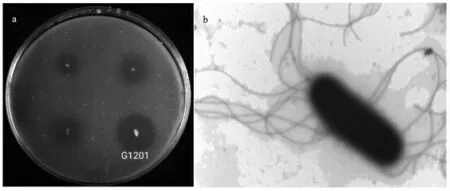
图1 菌株G1201 水解圈(a)和菌株G1201 电镜图(b)
通过厌氧性、运行性、触酶试验等生理生化性质鉴定,G1201 菌株的理化性质如表1 所示。参照《BERGEY’S MANUAL of Systematic Bacteriology Second Edition》(Paul等,2009)所述,确定该菌株与G.thermoleovorans 的理化性质一致。

表1 G1201 的生理生化特征
2.1.2 16S rRNA 基因序列鉴定 通过对G1201的16S rRNA 基因序列扩增后测序,其16S rDNA序列长度为1449 bp,将测得的序列上传至Gen-Bank 数据库(登录号MK208514.1),对16S rRNA基因序列构建系统发育树,结果显示该菌株与G.thermoleovorans 的相似性为99%(图2)。结合电镜观察及生理生化试验,确定该菌株为G.thermoleovorans。

图2 菌株G1201 和相似菌株的系统发育进化树
2.2 粗酶的制备及粗酶学性质的研究
2.2.1 粗酶的制备 参照GB/T23527-2009 建立的酪氨酸标准曲线为Y=0.0099X+0.0111,其R2值为0.9998。以该标曲计算不同硫酸铵沉淀后的酶活,沉淀经PBS(pH 7.5)复溶后,通过福林酚法测定其不同浓度的硫酸铵沉淀物的酶活,发现60%硫酸铵沉淀物复溶后酶活最高。因此以60%硫酸铵下的沉淀物作为粗酶。
2.2.2 粗酶酶学性质 通过测定不同温度下粗酶液的酶活,确定最适催化温度为70 ℃,酶活达16.82 U/mL(图3a)。温度耐受性结果显示,以70℃下测定的酶活为100%,30~ 60 ℃时酶活不变,80 ℃以上酶活有所降低,至100 ℃酶活仍能保持在50%以上(图3b)。
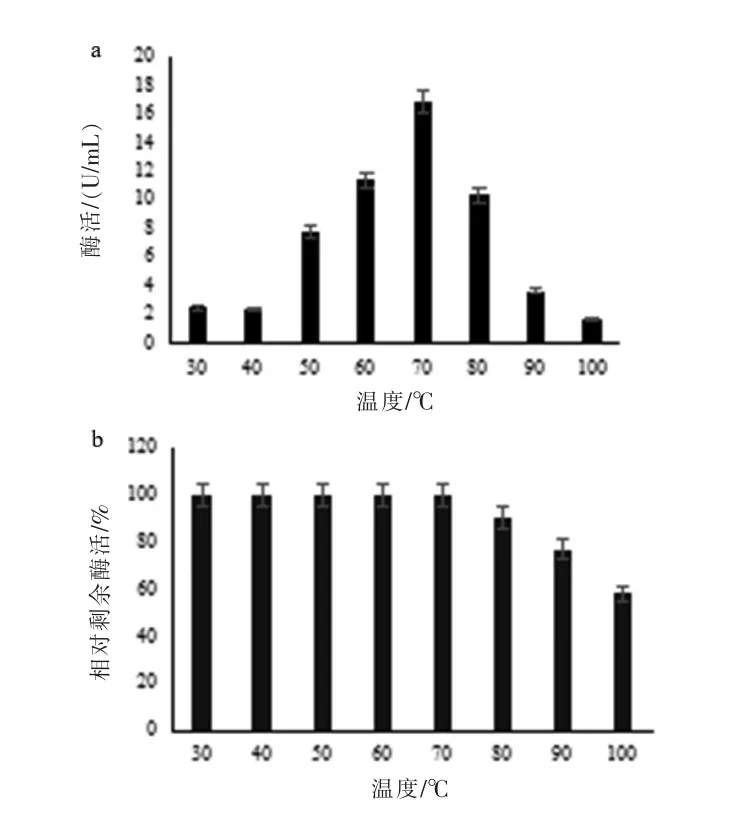
图3 温度对酶活力和稳定性的影响
通过测定不同pH 条件下粗酶液的酶活,确定其最适pH 为9.0(图4a)。pH 耐受性结果显示,以pH 9.0 下测定的酶活为100%,该蛋白酶在碱性环境中耐受性很好,pH 8.0~ 12.0 下的酶活均能达到90%以上,在酸性环境中该酶的的耐受性较差,pH 4.0~7.0下,酶活只有26%~50%(图4b)。

图4 pH 对酶活力和稳定性的影响
粗酶液在含有不同金属离子中孵育1 h后,以没有添加金属离子孵育的酶活视为100%,结果显示Ca2+和Mg2+对酶活具有促进作用,其酶活分别达到131%和117%,Mn2+对酶活基本没有影响,其他离子Co2+、Na+、Fe2+、Zn2+和Cu2+均对酶活力表现出有一定的抑制作用,抑制程度根据其蛋白酶残余酶活判断分别为46%、73%、75%、78%和93%(图5a)。粗酶液在含有不同有机溶剂中孵育1 h 后测酶活,以没有添加有机溶剂孵育的酶活视为100%,结果显示吐温20 和Triton X-100能够促进酶活,其酶活分别达131%和115%,吐温80 对酶活没有影响,而SDS 表现出一定的抑制作用(图6b)。而蛋白酶抑制剂DTNB、PMSF 和EDTA 表现出对酶活较强的抑制作用,其残余酶活分别为5%、60%、28%(图5b)。
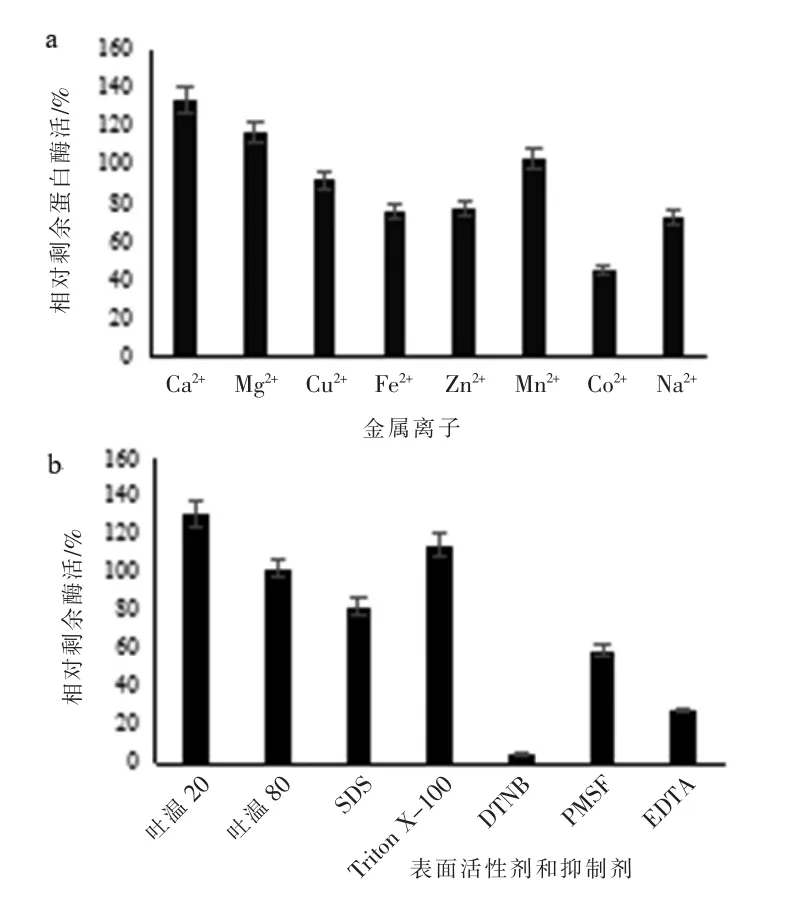
图5 金属离子及表面活性剂和抑制剂对酶活力和稳定性的影响
2.1.5 G1201 蛋白基因的克隆与表达载体的构建通过对克隆基因PCR 产物测序,其基因长度为1434 bp,将核酸序列上传至国家微生物科学数据中心(NMDC),基因编号:NMDCN0000Q22。在MDAMAN 软件中翻译该基因序列,得到其编码蛋白酶序列(图6)。在Genebank 数据库对克隆基因序列和翻译后的蛋白序列进行Blast,结果显示其为编码ser 蛋白酶的基因,属于属于丝氨酸蛋白酶家族,且存在该蛋白酶家族保守的氨基酸基序GTSMATP。在ExPasy 网站中用ProtParam tool,分析其序列特征,结果预测显示该蛋白稳定指数小于40为稳定蛋白,总平均亲水性为负值表明该蛋白为亲水性蛋白(表2)。将克隆的目的基因连接到质粒载体pET-28a上,通过双酶切验证(图7),成功构建了目的基因的重组质粒载体pET-28a(+)-ser。

图6 核酸序列和编码蛋白序列
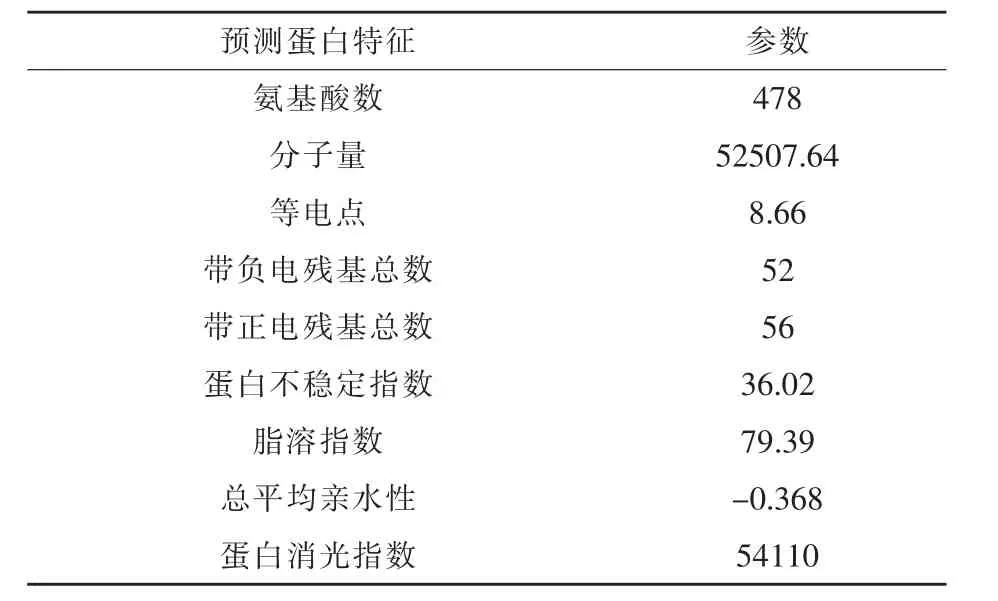
表2 蛋白理化性质预测
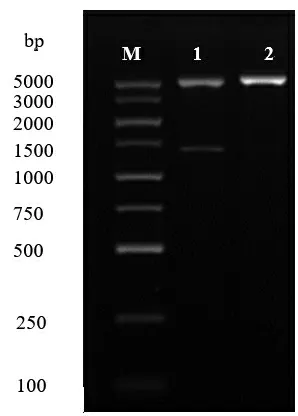
图7 重组质粒酶切
2.3.2 重组蛋白的表达及鉴定 提取重组质粒载体pET-28a(+)-ser 转入表达菌株大肠杆菌BL21(DE3)中,构建表达该蛋白酶的基因工程菌BL21(DE3)-pET-28a(+)-ser。以IPTG 为诱导物诱导目的基因表达,结果显示在诱导4 h 的发酵液中约50 kD 处出现了目的条带(图8)。

图8 蛋白酶诱导表达电泳图
2.3.3 重组蛋白酶的纯化 菌体破壁后电泳结果显示,目的蛋白以包涵体的形式存在于沉淀中,用8 mol/L 尿素对包涵体进行复性。之后,通过Ni 柱进行纯化,得到了目的蛋白(图9)。通过BCA 法测得的纯化后蛋白质浓度为0.108 mg/mL。

图9 目的蛋白的纯化电泳图
3 讨论
耐高温蛋白酶可适应高温加工环境,在饲料制粒、食品烘焙、高温洗涤等行业有重要的应用,一直以来都是研究的热点(Manavalan等,2020;Niu等,2017;Dadshahi等,2016;Charbonneau等,2012)。Geobacillus属的微生物作为2001年从Bacillus属分离出的嗜热菌群,其中蕴含了丰富的耐高温蛋白酶资源。目前,众多国内外的学者从该属微生物中分离得到了耐高温蛋白酶,如:Geobacillus.caldoproteolyticus(Chen等,2004),Geobacillus.subterraneus(Nazina,2001),Geobacillus.uzenensis(Nazina等,2001),Geobacillus.thermodenitrificans(Charbonneau等,2012)。本研究从蒸汽排气口土样中分离出的菌株G1201 能够产耐高温蛋白酶,经鉴定为G.thermoleovorans。比较值得注意的是,Geobacillus 属及近源的Bacillus属微生物革兰氏染色表现为阳性,而本研究中的G.thermoleovorans G1201 革兰氏染色表现为阴性。这 与 《BERGEY’S MANUAL of Systematic Bacteriology Second Edition》中对该种微生物的表述一致,G.thermoleovorans 种菌株革兰氏染色阴性和阳性结果均有发现(Bose等,2016;Paul等,2009)。推测,G.thermoleovorans 在细胞壁的进化中可能扮演了关键性角色。研究显示与G.thermoleovorans 近源的超嗜热古菌(Hyperthermophilic archaea)均为革兰氏阴性菌,两者在进化上逐渐分离,但可能保留了相同的细胞壁构成(Teuku等,2019)。
目前,对G.thermoleovorans 产高温淀粉酶(Mok等,2013)和高温脂肪酶(Abol Fotouh等,2016)有较多的研究,而对其产高温蛋白酶则鲜有报道(Teuku等,2019)。研究显示,Geobacillus 属菌株所产高温蛋白酶的适宜温度为55~ 90 ℃,适宜pH 为6~ 9(Baykara等,2021;Suleiman等,2020;Teuku等,2019;Charbonneau等,2012;Chen等,2004;Nazina,2001)。本研究中经60%硫酸铵沉淀的粗酶,最适温度为70 ℃,最适pH 为9.0,于上述结果类似。耐受性试验结果显示,该酶表现出较强的高温耐受性,在90 ℃以下均能达到酶活的80%以上,温度达到100 ℃时仍能维持50% 的酶活。同时,该蛋白酶在pH 8.0~ 12.0 的酶活均能达到90%以上,表明该酶在中性及偏碱性条件下具有良好的催化能力。与同种微生物产生的高温蛋白酶相比,G1201 所产蛋白酶具有更高的温度耐受性和pH 适应性(Teuku等,2019)。
Ca2+、Mg2+、吐温20 和Triton X-100 能够促进酶活,而Co2+、Na+、Fe2+、Zn2+、Cu2+以及SDS、DTNB、PMSF 和EDTA 则可以不同程度的抑制酶活。对温度和pH、金属离子、表面活性剂和蛋白酶抑制剂的耐受性分析表明,该酶能够耐受饲料制粒、食品烘焙和高温洗涤过程中面临的高温环境,具有良好的应用潜能。
结合该蛋白酶偏碱性的pH 适应性,以及酶活受Ca2+、Mg2+的促进作用。同时,具有丝氨酸蛋白酶家族的保守基因序列:GTSMATP(Laskar等,2011)。因此,可将该高温蛋白酶归为丝氨酸蛋白酶家族。克隆全长为1434 bp 的该高温蛋白酶基因构建大肠杆菌的表达载体pET-28a(+)-ser,在大肠杆菌BL21(DE3)中进行异源表达,结果显示52.5 kD 的目的蛋白以包涵体的形式存在。包涵体是大肠杆菌表达体系中常见形式,对目的蛋白的纯化和酶活具有较大影响。近年来,通过控制细菌热休克反应来改善重组蛋白的折叠,从而避免包涵体的形成已经有一些研究进展(Overton,2014)。将来,可通过该手段改善G.thermoleovorans 源高温蛋白酶异源表达过程中包涵体形成。
