红河橙叶表型遗传多样性研究
2022-08-03高玉红李月艳李凯迪牟凤娟
高玉红,李月艳,魏 萌,2,李凯迪,牟凤娟
(1 西南林业大学西南山地森林资源保育与利用教育部重点实验室,昆明,650224;2 东北林业大学林学院,哈尔滨,150040)
遗传多样性是物种进化的本质,广义指生物遗传信息的总和,狭义指种内不同群体和个体间的遗传变异[1]。同一物种、同一居群的不同个体间往往存在不同形式、不同程度的变异,这些变异由基因和环境共同作用决定,可以说遗传多样性和环境多样性共同造就了物种的表型多样性[2-3]。一般而言,分布越广泛的物种,其遗传多样性也越高。植物表型被广泛用于研究天然种群的遗传变异、亲缘关系及其与地理分布格局的关系等[4-9]。叶片是植物最重要的营养器官之一,也是对环境变化最为敏感的器官之一,其表型具有较大的可塑性,在不同光照、温度等条件下,宏观形态和微观结构均会产生相应的变化[10-12],叶表型可从侧面反映出环境对植物的影响程度以及植物对环境的适应能力。有研究发现,柠檬(Citruslimon)的翼叶长和翼叶宽在种内的变异更丰富,叶片长、叶片宽、翼叶长、叶柄长、单果质量、果实横径、果实纵径、果皮厚、囊瓣数、果心大小、可溶性固形物、可滴定酸含量、烘干率、刺百分比、花瓣长、花瓣宽等16个表型性状包含有较为丰富的遗传信息,且保留了柠檬80%以上的表型遗传多样性信息,可作为优先鉴定的评价指标[13]。贺思腾等[14]分析不同种源多油辣木(Moringaoleifera)叶片和种子形态变异,发现同种源地的多油辣木形态分化程度较高。汪洋等[15]研究了湖北红椿(Toonaciliata)16个天然居群的小叶表型,发现小叶表型变异主要来源于居群间,并提出种质资源保护时应增加不同居群和变异丰富的居群内个体数量。由此可见,研究不同居群间的叶表型变异可为优良种质资源的选择和保护提供参考[15-17]。
红河橙(CitrushongheensisYe et al.)是20世纪70年代在云南省红河县首次发现的野生柑桔种类,当地哈尼族人称其为“阿蕾”,主要分布于海拔800~2 000 m的山坡林地中,与芭蕉(Musabasjoo)、竹类等多种植物混生[18-19]。红河橙是云南特有的珍稀柑桔种质资源,仅在元江、红河南岸有零星分布,其单身复叶具有极为发达的翼叶。与其他大翼橙种类相比,红河橙的翼叶较本叶宽而长,其翼叶的长度可达本叶的1~3倍;花大型,直径达3.0~3.2 cm,常数花组成总状花序,少单花腋生,花丝分离。这些形态特征均显示出红河橙的原始性,同时也为“云南是柑桔的重要起源地之一”这一观点提供了佐证[20-21]。红河橙的树干粗壮、树形高大,叶片浓绿且具光泽,常被当地百姓种植于村寨周边以美化环境,其幼果干燥后可作中药“枳壳”用;成熟叶片的精油含量丰富,常用作牛羊肉的辛香调料。此外,红河橙在云南冬春干旱条件下生长发育良好,且存活年限较长,可作为柑桔抗旱育种以及砧木的优良种质资源[22]。目前,有关红河橙的研究主要集中于系统分类地位[23-27]及分子遗传多样性[28-29]。Yang等[29]通过分子标记研究了红河橙的遗传多样性,结果表明红河橙分子遗传多样性水平较高,其遗传多样性高于贵州苏铁(Cycasguizhouensis)和银杉(Cathayaargyrophylla)等濒危植物。牟凤娟等[22]对比分析红河橙栽培和野生居群叶表型多样性的结果表明,红河橙的多样性水平较高,栽培居群和野生居群变异程度不同。但以上研究均未指出红河橙遗传变异主要来源是居群间还是居群内,缺少对红河橙种质资源利用与保护的探讨。野外调查发现,红河橙大部分是人为移栽于村寨周边,且移栽年限较长,同一居群中的个体来源较为丰富。笔者对云南红河橙11个居群的叶表型多样性进行了研究,以期为红河橙的物种保护及资源开发利用提供参考。
1 材料与方法
1.1 材料
对云南省元江县、红河县和绿春县分布的红河橙11个居群进行调查和采样,即元江县那诺乡打芒村(YJNNDM)10株,元江县那诺乡猪街村(YJNNZJ)11株,元江县羊街乡坝木村(YJYJBM)6株,元江县羊街乡东林村(YJYJDL)5株,元江县羊街乡羊街村(YJYJYJ)11株,红河县甲寅乡阿撒村下寨(HHJYAS)10株,红河县乐育乡龙虾村(HHLYLX)8株,红河县乐育乡阿蕾村(HHLYAL)2株,红河县乐育乡娘普村(HHLYNP)6株,红河县大羊街乡(HHDYJ)14株,绿春县大兴镇瓦那村(LCDXWN)8株。
1.2 方法
1.2.1 数据采集 每个居群随机选取至少20个个体,不足20株的选取所有个体。每株样本植株采集树冠上、中、下不同部位枝条,摘取成熟、完整叶片(要求叶片生长期至少1年以上,无病虫害为害痕迹),混匀后采用随机抽样,每个个体至少取60枚叶备用。
参考前人研究中关于红河橙[22]和大翼厚皮橙(C.macropteravar.kerrii)[30]叶表型时选取的指标和测量方法,本研究测量并计算12个叶表型,包括6个叶片测量以及6个比值表型:叶总长度(Total length of leaf,TLL)、翼叶长度(Length of winged leaf,LWL)、翼叶宽度(Width of winged leaf,WWL)、本叶长度(Length of leaflet,LL)、本叶宽度(Width of leaflet,WL)、总叶柄长度(Length of common petiole,LCP),其中,总叶柄长度(LCP)=翼叶长度(LWL)+叶柄长度(LC);翼叶长/叶总长(LWL/TLL)、翼叶长/翼叶宽(LWL/WWL)、翼叶长/本叶长(LWL/LL)、翼叶宽/本叶宽(WWL/WL)、本叶长/叶总长(LL/TLL)、本叶长/本叶宽(LL/WL)。
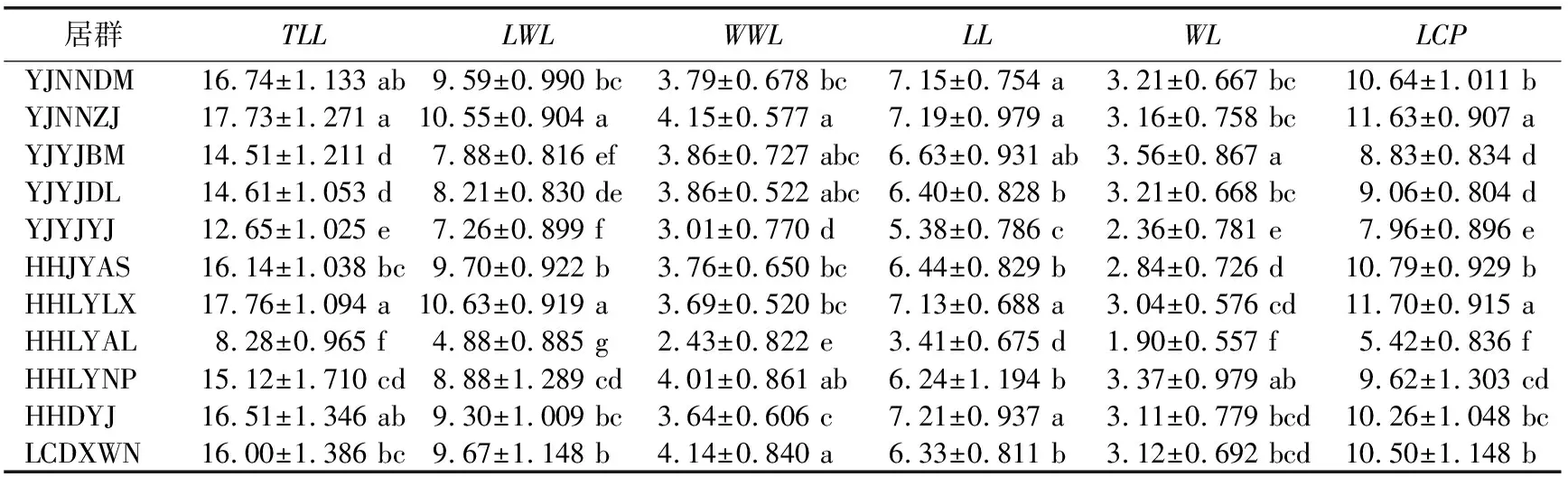
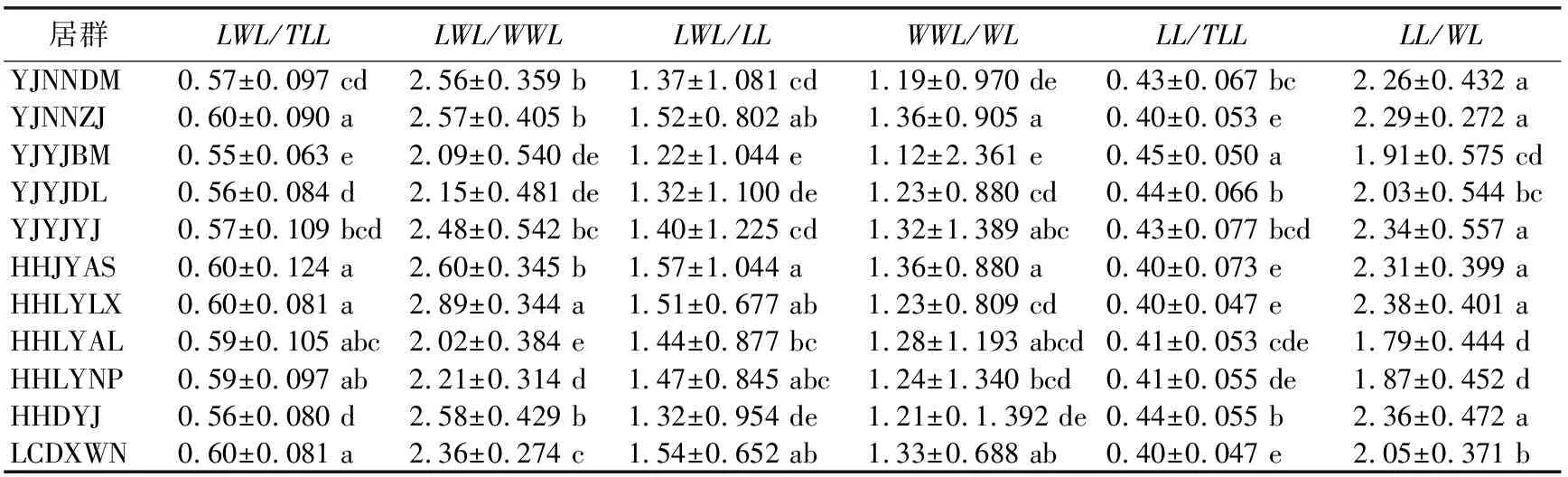
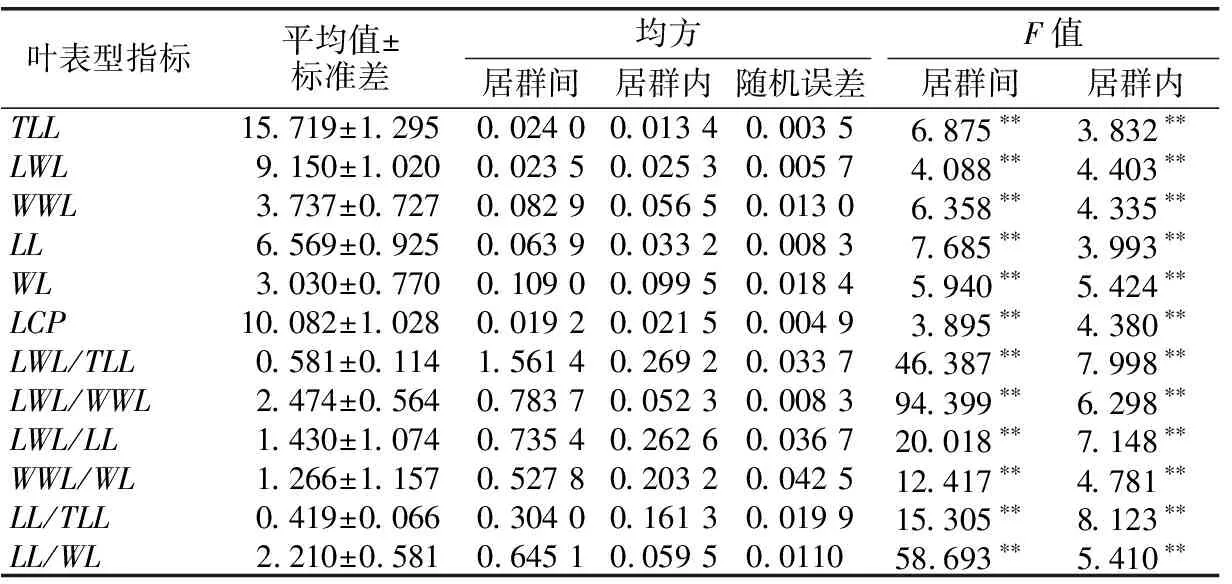
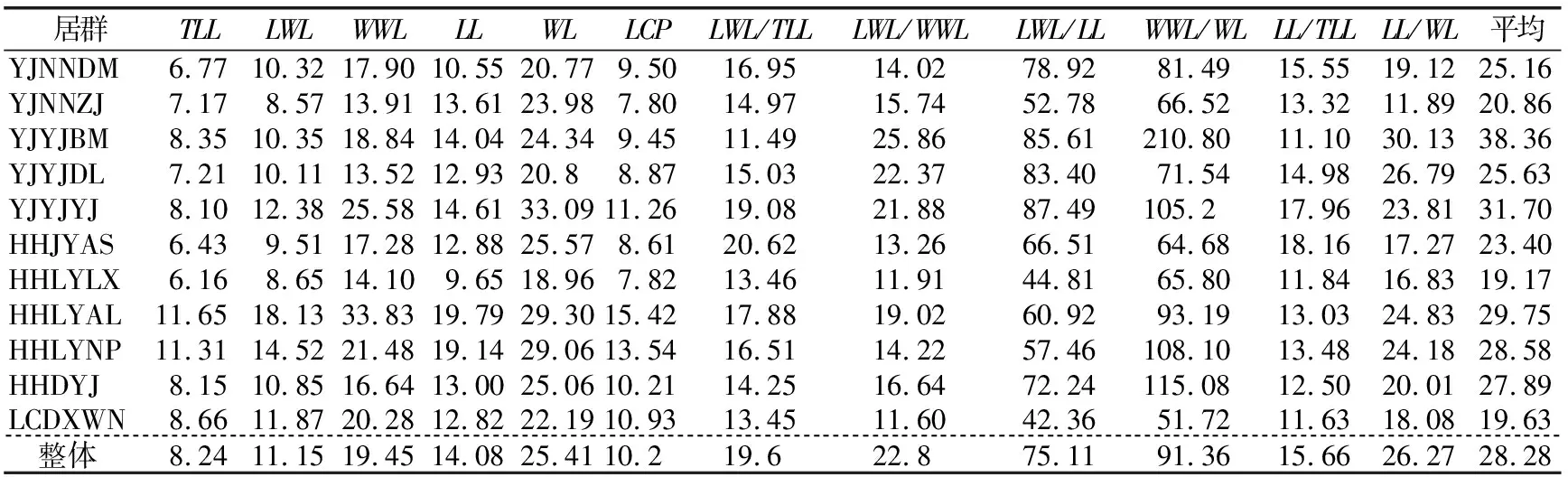
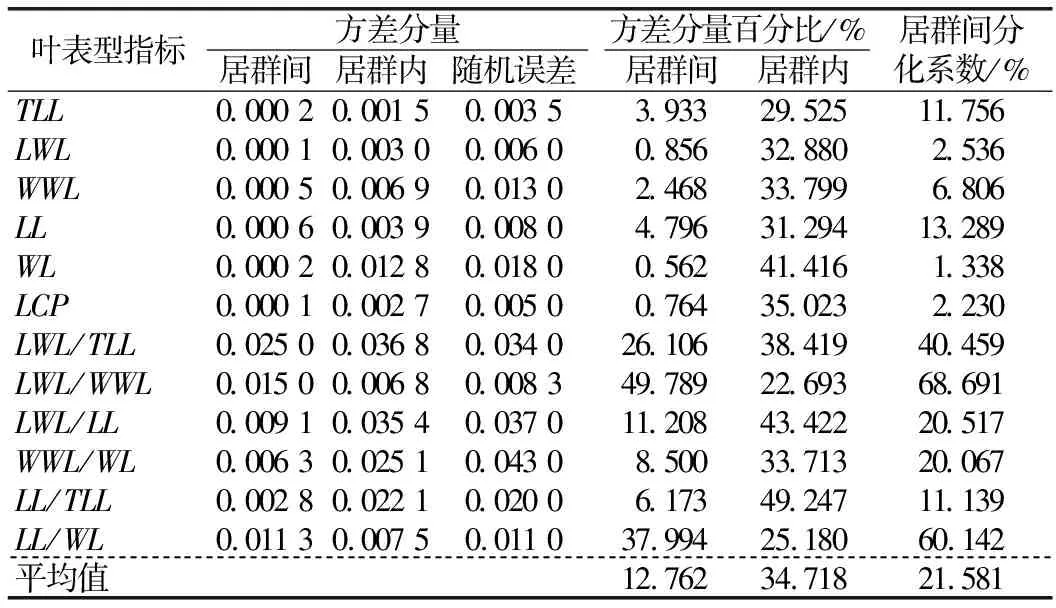
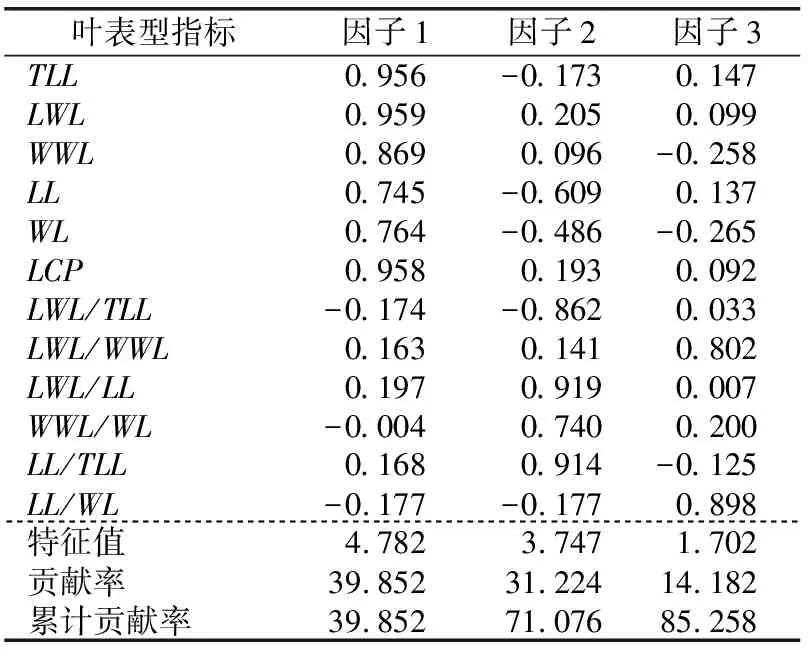
1.2.2 数据分析 用Excel 2010软件进行平均值、标准差和变异系数计算,用SPSS 19.0软件进行方差分析、差异显著性检验、相关性分析以及主成分分析。将原始数据取对数函数进行数据转换,得到映射到[0,1]范围内标准化处理的数据,对标准化后的数据进行变异程度分析、方差分析和多重比较,探索居群内和居群间各表型的变异规律和特征。用变异系数(Coefficient of variation,CV)表示变异程度,变异程度分为低级(CV<10%)、中级(10%≤CV≤20%)、高级(20% 从试验结果看出,红河橙叶表型6个测量表型和6个比值表型在居群间存在显著差异。YJNNZJ居群的翼叶宽(4.15 cm)最大,YJYJBM居群本叶宽(3.56 cm)最大,HHLYLX居群叶总长(17.76 cm)、总叶柄长(11.70 cm)最大,HHDYJ居群本叶长(7.21 cm)最大,上述5个叶表型均是HHLYAL居群最小。HHLYAL和HHLYNP居群的6个比值性状较为接近,除翼叶长/翼叶宽(LWL/WWL)外,其他5个比值性状在两个居群间差异不显著;但是在测量性状中,HHLYAL居群的6个叶表型测量性状平均值分别约为HHLYNP居群的1/2,差异均显著。HHLYAL和HHLYNP居群的地理位置相距较近,海拔相差约100 m,HHLYAL居群所处海拔相对较高,这可能是造成两个居群间叶表型测量性状相差大而比值性状较为接近的原因(见表1和表2)。 表1 红河橙不同居群之间叶表型测量性状差异 表2 红河橙不同居群间叶比值性状差异 方差分析结果显示,12个叶表型在居群间和居群内的差异均达极显著性(p<0.01)(见表3)。表明,红河橙叶表型在居群间和居群内均存在着丰富的变异。 表3 红河橙居群间和居群内叶表型指标方差分析 变异系数可反映性状值的离散特征,变异系数越大则离散度越大,物种的遗传多样性也越丰富。分析结果显示,红河橙居群内叶表型性状的变异系数在6.16%~210.80%之间,最高为YJYJBM居群的翼叶宽/本叶宽(210.80%),属于强变异,其次是HHDYJ居群的翼叶宽/本叶宽(115.08%);最小为HHLYLX居群的叶总长(6.16%),且该表型在11个居群的12个表型中变异最小,说明叶总长这个叶片表型与其他叶表型相比表现最为稳定,可将其作为种类鉴定的一项可靠表型性状(见表4)。 从供试红河橙整体材料(不分居群的所有个体)来看,12个叶表型性状的变异系数在8.24%~91.36%之间。其中,最大的是翼叶宽/本叶宽(91.36%),其次为翼叶长/本叶长(75.11%),再次为本叶长/本叶宽(26.27%)、本叶宽(25.41%)和翼叶长/翼叶宽(22.80%),这5个表型性状的变异系数均大于20%,变异程度高,较不稳定;翼叶长/叶总长(19.60%)、翼叶宽(19.45%)、本叶长/叶总长(15.66%)、本叶长(14.08%)、翼叶长(11.15%)、总叶柄长(10.20%)这6个叶表型性状的变异程度为中等;叶总长(8.24%)的变异程度低(见表4)。 从各居群12个叶表型性状变异系数平均值的大小来看,YJYJBM居群(38.36%)存在丰富的变异,而HHLYLX居群(19.17%)的多样性水平相对较低(见表4)。 表4 红河橙11个居群叶表型指标的变异系数(CV) % 从方差分量来看,同一叶表型性状在居群间和居群内的变异程度不同。12个叶表型性状在居群间的方差分量百分比平均值(12.762%)均小于居群内(34.718%),表明居群内分化程度相对较高,而居群间分化程度较低。12个叶表型性状的居群间分化系数(Vst)平均值仅为21.581%,表明居群间变异在总体变异中的占比小,居群内变异才是红河橙叶表型性状变异的主要来源。在12个叶表型性状中,居群间变异最大的是翼叶长/翼叶宽(68.691%),其次为本叶长/本叶宽(60.142%)和翼叶长/叶总长(40.459%),Vst均大于40%,最小的是翼叶宽(1.338%)(见表5)。 表5 红河橙叶表型指标的方差分量及居群间分化系数 对12个叶表型性状指标进行主成分分析的结果显示,前3个主成分累计贡献率达到85.258%,很好地代表了12个叶表型性状。第一主成分的特征值为4.782,贡献率为39.852%,在其特征向量中,绝对值较高的是叶总长(0.956)、翼叶长度(0.959)、翼叶宽(0.869)、本叶长(0.745)、本叶宽(0.764)、总叶柄长度(0.958),该类表型性状与叶片绝对大小有关,故第1主成分为叶片绝对大小因子。第2主成分的特征值为3.747,贡献率为31.224%,在其特征向量中,绝对值较高的是翼叶长/叶总长(0.862)、翼叶长/本叶长(0.919)、翼叶宽/本叶宽(0.740)、本叶长/叶总长(0.914),与翼叶相对大小有关,故第2主成分为翼叶相对大小因子。第3主成分的特征值为1.702,贡献率为14.182%,在其特征向量中,绝对值最高的表型性状是翼叶长/翼叶宽(0.802)、本叶宽/本叶宽(0.898),与叶片叶形指数有关,故第3主成分为叶形指数因子。比较各叶表型性状特征向量绝对值大小可知,叶总长(0.956)、翼叶长(0.959)、总叶柄长(0.958)、翼叶长/本叶长(0.919)、本叶长/叶总长(0.914)这5个叶表型性状是红河橙叶片变异的主要表现(见表6)。 表6 红河橙12个叶表型性状主成分分析 红河橙单身复叶的翼叶极发达,可达本叶长的1~3倍。其中,翼叶的绝对大小(如长度、宽度)、翼叶形状(如翼叶长/翼叶宽)以及翼叶在单身复叶中的相对大小(如翼叶长/本叶长、翼叶长/叶总长)等叶表型对于红河橙的种类鉴定具有非常重要的意义。在本研究中,红河橙11个居群的翼叶长/叶总长≥0.55,而本叶长/叶总长≤0.45,符合红河橙翼叶较本叶更为发达的形态表现[18]。红河橙叶表型在居群间以及居群内均存在着丰富的变异,与分子标记研究结果[21]一致。 前人认为,当变异系数(CV)大于10%时,则样本内表现出来的差异较为明显[34],且变异系数与植物适应环境的能力成正比,变异系数越大,植物适应的环境越广[4-5]。在本研究的11个居群中,多样性水平较高的是YJYJBM居群(12个表型性状的CV平均值为38.36%),较低的是HHLYLX居群(19.17%)。在红河橙的12个叶表型性状中,叶总长的变异系数(8.24%)最小,表明该叶表型性状的表现最为稳定,可考虑将其作为种类鉴定的一项可靠表型性状。此外,本研究还发现红河橙叶片变异的普遍现象为翼叶相对大小(翼叶宽/本叶宽,翼叶长/本叶长)>叶片形状指数(本叶长/本叶宽,翼叶长/翼叶宽)>叶片绝对大小(长度、宽度)。有研究发现大翼橙亚属下的大翼厚皮橙的平均叶总长为18.68 cm,翼叶长和翼叶宽分别为7.72和4.94 cm,本叶长和本叶宽分别为10.96和6.05 cm[30]。在本研究中,红河橙的平均叶总长(15.719 cm)更小,本叶长(6.569 cm)和本叶宽(3.030 cm)更小,翼叶长(9.150 cm)和翼叶宽(3.737 cm)相对更大。上述研究结果表明,红河橙的翼叶较大翼厚皮橙发达。 在本研究中,比较红河橙叶表型居群间及居群内的方差分量,发现居群内变异是红河橙叶表型变异的主要来源,大部分叶表型(除LWL/WWL和LL/WL外)在居群间分化程度较低。这与南方红豆杉(Taxuswallichianavar.mairei)叶片表型的变异情况类似[17],但与红椿[15]、山苍子(Litseacubeba)[35]、大别山核桃(Caryadabieshanensis)[36]等其他植物的叶表型变异主要来源于居群间的情况不同。在本研究中,红河橙居群间叶表型分化程度较低的原因,可能与该物种分布较为狭窄,地理位置较近,而人为移栽时又有相似的选择倾向有关。鉴于此,在收集保存红河橙种质资源时,应选择多样性高的居群(如元江县羊街乡坝木村),适当增加居群内个体数,并优先收集表型特异的个体。 本研究通过对91株不同来源的红河橙的12个叶表型性状进行主成分分析,结果表明前3个主成分的累计贡献率达85.258%,其中,叶总长、翼叶长、总叶柄长、翼叶长/本叶长、本叶长/叶总长等叶表型性状是红河橙叶片变异的主要表现。红河橙一般生长在山坡杂木林中,光照强度及时间长短均会影响其叶片和植株的生长发育。 红河橙居群间和居群内叶表型均存在丰富的变异,居群内变异是其叶表型变异的主要来源。YJYJBM居群的叶片变异最丰富,其产生优质变异的可能性较大,可考虑将其作为优良种质资源选育的主要来源。YINNZJ居群的叶片总体较大,可以考虑在叶片香料或提取香茅醛精油方面加以开发。在12个叶表型性状中,红河橙表现最稳定的表型性状是叶总长,可考虑将该表型性状用于种类鉴定。在调查收集红河橙种质资源时,应注意适当增加居群内部的个体数量,优先收集有特异表型的植株。2 结果与分析
2.1 居群间叶表型差异
2.2 叶表型变异特征
2.4 叶表型性状主成分分析
3 讨论
3.1 红河橙叶表型变异的特征
3.2 红河橙居群间叶表型分化的影响因素
3.3 红河橙叶表型多样性的主要表型性状
4 结论
