长链非编码RNA GAS5靶向调控miR-424表达抑制宫颈癌HeLa细胞增殖
2022-08-02周勇徐志渊王骞
周勇, 徐志渊, 王骞
(1.南方医科大学深圳医院肿瘤科,广东省深圳市 518000;2.香港大学深圳医院肿瘤科,广东省深圳市 518058)
宫颈癌是女性最常见的恶性肿瘤之一,其形成涉及多种致癌基因的激活和抑癌基因的失活[1]。长链非编码RNA(long non-coding RNA,lncRNA)是一类长度超过200个核苷酸且没有编码蛋白质潜能的非编码RNA,其在癌症中可表达异常,作为竞争性内源性RNA(competing endogenous RNA,ceRNA)或分子海绵调控微小RNA(microRNA,miRNA)靶基因表达,在肿瘤的发生和发展中发挥重要作用[2]。生长停滞特异性5(growth arrest specificity 5,GAS5)在胃癌中扮演了分子海绵的角色调控miR-23a[3]。在胶质瘤中,GAS5过表达通过靶向多种miRNA分子抑制胶质瘤细胞恶性表型[4]。同时,GAS5被证实在宫颈癌中起抑癌作用,但其潜在分子机制尚不清楚[5]。因此,本研究重点探讨宫颈癌HeLa细胞GAS5与miRNA的相互作用,以及GAS5表达对HeLa细胞增殖的影响,为宫颈癌的药物开发提供参考。
1 材料和方法
1.1 主要试剂与仪器
正常宫颈Ect1/E6E7细胞和宫颈癌HeLa细胞(中科院上海细胞库);Lipofectamine 3000转染试剂盒、阴性对照(siR-NC)、GAS5 RNAi、pCDNA3.0-HA-GAS5质粒、miR-424 inhibitor和miR-424-mimic(上海吉凯生物公司);双荧光素酶检测试剂盒(美国Promega公司);TRIzol试剂(美国Invitrogen公司);mRNA反转录试剂盒和荧光定量PCR试剂盒(北京金豪制药股份有限公司);四噻唑蓝(methyl tihiazolyl tetrazolium,MTT)(日本同仁公司);(Annexin)V-FITC细胞凋亡检测试剂盒(武汉普诺赛生命科技有限公司);DNA甲基转移酶(DNA methyltransferase,Dnmts)、组蛋白甲基化转移酶(enhancer of zeste homolog2,EZH2)、丝氨酸/苏氨酸蛋白激酶3(serine/threonine protein kinase 3,Akt3)抗体(美国Santa Cruz公司);HRP标记山羊抗鼠二抗(武汉博士德生物工程公司);StepOne TM实时荧光定量PCR仪(美国应用生物系统公司);FACSVia流式细胞仪(美国BD公司)。PCR引物序列由吉林省奇健生物技术有限公司合成,GAS5上游5′-TCCTTAGGCATCACCTAGCC-3′,下游5′-GATGGAGGAAGTAGAGTCATTGG-3′;miR-424上游5′-G AGGCGATGCTATATTGTATCGT-3′,下游5′-GATCTTGCTTCTACTCCTACACGG-3′;U6上游5′-CATGCATCGGCATCACA-3,下游5′-AATGCTTCATGAATGTGCGT-3′。
1.2 组织来源
组织样本来自本院2018年1月—2020年12月宫颈活检后病理确诊的原发性宫颈癌标本43例,年龄(49.16±5.43)岁,肿瘤直径≤4 cm 19例、>4 cm 24例,转移22例,FIGO分型Ⅰb~Ⅱa期17例、Ⅱb-Ⅲa期26例。选取同期因子宫肌瘤手术获取的正常宫颈组织20例,年龄(48.23±4.15)岁。术后组织迅速保存于液氮中以备提取RNA,供后续试验用。本研究获得患者知情同意并签署知情同意书,通过本院伦理审查委员会批准。
1.3 细胞培养和转染
37 ℃、5%CO2培养Ect1/E6E7细胞和HeLa细胞,融合至70%时进行下一步实验。将HeLa细胞分为阴性对照组、miR-NC组、GAS5 RNAi组和GAS5质粒组。阴性对照组不处理,miR-NC组、GAS5 RNAi组和GAS5质粒组用Lipofectamine法分别将miR-NC、GAS5 RNAi和pCDNA3.0-HA-GAS5质粒转染到HeLa细胞,继续培养48 h后进行相关检测。
1.4 荧光素酶报告基因检测
通过生物信息学网站(www.Targetscan.org)预测筛选靶向GAS5的miRNA发现,GAS5与miR-424有潜在结合位点。将GAS5野生型(WT)/突变型(MUT)与miR-NC、miR-424 inhibitor和miR-424-mimic转染到对数生长期HeLa细胞中,转染48 h后,用双荧光素酶检测试剂分析荧光素酶活性。
1.5 实时荧光PCR检测GAS5和miR-424 mRNA
取标本组织、对数生长期Ect1/E6E7细胞、HeLa细胞及3组转染细胞接种于96孔板中,48 h后加入TRIzol提取总RNA,测定RNA,根据mRNA反转录试剂盒说明书操作将总RNA逆转录为cDNA,根据荧光定量PCR试剂盒说明操作,制备20 μL反应体系,在PCR扩增仪中进行扩增。反应条件为预变性95 ℃ 30 s、变性95 ℃ 5 s、60 ℃ 44 s、40个循环,采集荧光,2-ΔΔCt法计算GAS5和miR-424 mRNA相对表达量(以U6为内对照)。
1.6 MTT法检测细胞增殖
处理后的细胞接种于96孔板中(5×103个/孔),48 h后每孔加入MTT(20 μL个/孔),继续培养4 h,离心弃培养基,加入二甲基亚砜(200 μL个/孔),振荡10 min,用酶标仪测定光密度值(OD值),计算细胞增殖率。
1.7 流式细胞术检测细胞凋亡
处理后的细胞接种于6孔板中(3×105个/孔),48 h后收获细胞,用预冷磷酸盐缓冲液洗涤,离心弃上清,分别加入5 μL膜联蛋白(Annexin)V-FITC与PI,充分混匀后25 ℃避光孵育15 min,FACSVia型流式细胞仪检测细胞凋亡率。
1.8 蛋白印迹法检测Dnmts、EZH2和Akt3蛋白
处理后的细胞接种于6孔板中(3×105个/孔),48 h后收获细胞,加入100 μL细胞蛋白抽提液,冰上裂解30 min,进行电泳,转膜,将膜与Dnmts(1∶500)、EZH2(1∶200)和Akt3(1∶400)4 ℃孵育过夜,洗膜后用HRP标记的二抗25 ℃孵育30 min,显色后采集图像并分析(以GAPDH为内对照)。
1.9 统计学分析
2 结 果
2.1 lncRNA GAS5对miR-424的靶向调节作用
转染miR-424-mimic可明显降低GAS5-WT的荧光素酶活性,对GAS5-MUT没有影响,而miR-424-inhibitor则表现出相反作用,表明GAS5与miR-424存在靶向调节作用(P<0.05;图1)。

图1 lncRNA GAS5对miR-424的靶向调节作用A为互补配对;B为荧光素酶活性。a为P<0.05,与阴性对照组比较;b为P<0.05,与miR-NC组比较;c为P<0.05,与miR-424 inhibitor组比较。
2.2 各组GAS5和miR-424 mRNA表达的比较
宫颈癌组织、HeLa细胞GAS5和miR-424 mRNA水平分别低于正常宫颈组织或Ect1/E6E7细胞(P<0.05;表1)。以GAS5或miR-424 mRNA均数为标准,将宫颈癌患者分为低表达组和高表达组。

表1 GAS5和miR-424 mRNA在组织和细胞中表达的比较
2.3 不同临床病理特征宫颈癌患者GAS5、miR-424表达的比较
GAS5和miR-424低表达率有转移的宫颈癌患者高于无转移患者,FIGO分型Ⅱb~Ⅲa型患者高于Ⅰb~Ⅱa型患者(P<0.05;表2)。

表2 不同临床病理特征宫颈癌患者GAS5、miR-424表达比较 单位:例(%)
2.4 转染GAS5对GAS5和miR-424 mRNA表达的影响
与阴性对照组和miR-NC组比较,HeLa细胞GAS5和miR-424 mRNA相对表达量GAS5 RNAi组降低,GAS5质粒组增加;GAS5质粒组高于GAS5 RNAi组(P<0.05;表3)。

表3 转染GAS5对HeLa细胞GAS5和miR-424 mRNA表达的影响
2.5 转染GAS5对细胞增殖及凋亡的影响
与阴性对照组和miR-NC组比较,HeLa细胞增殖率GAS5 RNAi组升高,GAS5质粒组降低;HeLa细胞凋亡率GAS5 RNAi组降低,GAS5质粒组升高(P<0.05)。与GAS5 RNAi组比较,GAS5质粒组细胞增殖率降低,凋亡率升高(P<0.05;图2)。

图2 转染GAS5对HeLa细胞增殖和凋亡的影响1为阴性对照组;2为miR-NC组;3为GAS5 RNAi组;4为GAS5质粒组。a为P<0.05,与阴性对照组比较;b为P<0.05,与miR-NC组比较;c为P<0.05,与GAS5 RNAi组比较。
2.6 转染GAS5对Dnmts、EZH2和Akt3蛋白表达的影响
与阴性对照组和miR-NC组比较,HeLa细胞Dnmts、EZH2表达GAS5 RNAi组降低,GAS5质粒组增加;Akt3表达GAS5 RNAi组增加,GAS5质粒组降低(P<0.05)。与GAS5 RNAi组比较,GAS5质粒组Dnmts、EZH2表达增加,Akt3表达降低(P<0.05;图3)。
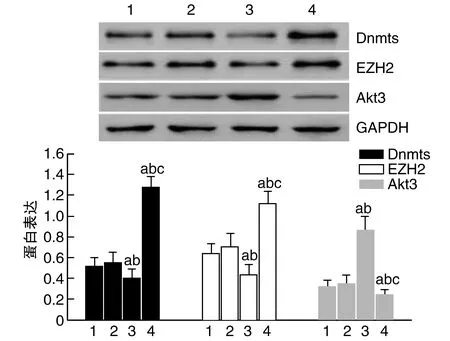
图3 转染GAS5对HeLa细胞中Dnmts、EZH2和Akt3蛋白表达的影响1为阴性对照组;2为miR-NC组;3为GAS5 RNAi组;4为GAS5质粒组。a为P<0.05,与阴性对照组比较;b为P<0.05,与miR-NC组比较;c为P<0.05,与GAS5 RNAi组比较。
3 讨 论
lncRNA是细胞生物学中重要的调控因子,其异常表达与肿瘤发生密切相关。研究表明,GAS5在肿瘤进展中起着重要作用。GAS5下调与结直肠癌和非小细胞肺癌的晚期肿瘤分期和低生存率相关[6]。GAS5在胰腺癌中的异常表达与胰腺癌侵袭及预后不良有关[7]。本研究结果显示,GAS5在HeLa细胞和宫颈癌组织中表达下调。此外,通过基因干预,促进GAS5表达能抑制HeLa细胞增殖,诱导HeLa细胞凋亡。这些结果提示GAS5在宫颈癌中是一种抑癌基因。近年来的研究表明,lncRNA与miRNA之间存在一种新的调控机制。例如,lncRNA MALAT1通过海绵化miR-22作为竞争性内源性RNA促进恶性黑色素瘤的生长和转移[8];lncRNA CASC2通过海绵化miR-18a在结直肠癌中作为内源性竞争RNA发挥作用[9]。本研究通过生物信息学分析和随后的双荧光素酶报告基因检测证明,GAS5作为ceRNA竞争性海绵miR-424。同时,RT-PCR显示HeLa细胞和宫颈癌组织中miR-424表达降低。此外,促进GAS5表达能诱导miR-424表达。在癌症相关miRNA中,miR-424可以作为肿瘤抑制因子,例如在乳腺癌、宫颈癌和骨肉瘤中低表达[10]。在胶质瘤组织中,miR-424水平明显低于邻近正常脑组织,miR-424过表达能促进神经胶质瘤细胞的凋亡,表明miR-424具有肿瘤抑制活性[11]。因此,GAS5可能至少部分通过诱导miR-424的表达来抑制宫颈癌细胞增殖。有趣的是,miR-424启动子区域内的CpG岛甲基化是其在肿瘤组织中下调的原因。然而,对负责miR-424启动子甲基化或其肿瘤抑制活性的机制知之甚少。
DNA甲基化的全基因组模式在恶性肿瘤组织和正常组织之间发生显著变化,与疾病进展和患者存活率相关。Dnmts通过与不同物质形成复合物,严格调节DNA序列特异性和非特异性CpG甲基化。同时,高甲基化基因在EZH2基因中也高度富集,支持EZH2介导DNA从头甲基化[12]。从机制上讲,EZH2和Dnmts之间的直接相互作用充当后者的募集平台,并且是EZH2靶向启动子的DNA甲基化所必需的[13]。同时,EZH2是维持miRNA和lncRNA表达所必需的,这是通过EZH2和Dnmts之间的直接相互作用以及后者与启动子的隔离来实现的。进一步实验表明,GAS5可以上调EZH2,并与Dnmts直接相互作用[14]。在胶质瘤细胞中,过表达EZH2逆转了miR-424启动子内的CpG甲基化,并提高miR-424的表达[15]。对肝癌的研究发现,Akt3是miR-424的直接靶点,可以减弱miR-424的生物学效应[16]。宫颈癌或肺癌的研究也支持miR-424和磷脂酰肌醇3激酶/Akt信号之间的负调控[17]。使用miR-424模拟物或抑制剂治疗胶质瘤细胞时Akt3的变化与多种活性基因的变化相关,包括细胞周期蛋白D1(Cyclin D1)和B细胞淋巴瘤/白血病-2,这表明抑制Akt3可能至少有助于miR-424对细胞生长的影响[18-19]。本研究结果显示,促进GAS5表达后,HeLa细胞中Dnmts和EZH2表达增加、Akt3表达降低,也证实了GAS5通过调控miR-424发挥抗肿瘤细胞增殖的作用。
综上所述,lncRNA GAS5通过靶向调控miR-424的表达,抑制宫颈癌HeLa细胞增殖,可能成为宫颈癌患者治疗的潜在靶点。
