雌激素对骨骼肌成肌细胞增殖、分化及迁移的影响
2022-07-30赵婷郭欣雨郑雨林谭戈李偲张若男吴艳唐俊明
赵婷 郭欣雨 郑雨林 谭戈 李偲 张若男 吴艳,2 唐俊明,2
(湖北医药学院基础医学院 1研究所,湖北 十堰 442000;2生理学教研室)
骨骼肌是支持身体活动最重要的器官之一,其质量和强度随着年龄的增长而下降,这种现象被称为骨骼肌减少症〔1〕,该疾病严重影响老年人活动和生活质量〔2〕。临床观察发现绝经后妇女骨骼肌减少症与雌激素缺乏有关〔3〕,进一步动物实验研究也证实雌激素不足可影响雌性小鼠的肌肉力量和骨骼肌再生,而外源性雌激素的补充可减少肌肉的损失和功能障碍〔4,5〕。骨骼肌细胞是由成肌细胞融合形成的多核细胞。小鼠骨骼肌成肌细胞(C2C12)在形态和特征上是一致的,可以无限次传代培养,雌激素对C2C12的分化作用得到部分学者关注,但目前研究结论仍存在争议〔6,7〕。本研究采用连续单次给予17β-雌二醇(E2)或其受体拮抗剂模拟升高或降低雌激素水平,观察雌激素对C2C12增殖、迁移及分化的影响,探讨雌激素是否通过影响骨骼肌自我更新或修复而参与骨骼肌萎缩过程。
1 材料与方法
1.1细胞与试剂 小鼠C2C12(中科院上海细胞库);DMEM培养基、马血清(HS)和新生胎牛血清(美国Gibco公司);17β-E2(美国Sigma公司);anti-ERα、anti-ERβ(北京Bioss公司);雌激素受体抑制剂ICI 182,780(美国Tocris Cookson Incorporation公司);anti-肌球蛋白重链(MYHC)抗体、anti-肌细胞生成蛋白(MyoG)抗体(英国Abcam公司);四甲基偶氮唑蓝(MTT,碧云天公司)。
1.2C2C12培养和诱导分化 将冻存的C2C12常规复苏,接种于75 cm2培养皿中,培养条件为10%胎牛血清+高糖DMEM培养基即为增殖培养基,置于37℃、5%CO2细胞培养箱中培养,每48 h换液1次,待细胞增殖融合达80%~90%时,按照1∶3的比例传代。传代的C2C12按上述条件继续培养。当细胞融合达70%时,换用含2% HS的高糖DMEM分化培养基进行成肌诱导分化,当细胞分化4 d后,在光学显微镜下观察见已经形成典型的肌管,采用免疫荧光法检测成肌细胞分化标志物MYHC的表达均呈强阳性,表明成肌细胞培养成功。
1.3C2C12雌激素受体表达检测 Western印迹:取C2C12,经放射免疫沉淀试验(RIPA)裂解、离心收蛋白,行二喹啉甲酸(BCA)法进行蛋白定量。按分组各上样50 μg蛋白,经10%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)后,转聚偏氟乙烯(PVDF)膜。用TBST配制5%的脱脂奶粉低速摇床封闭40 min,分别放入含有α-Tubulin、ERα、ERβ等不同一抗的封闭袋中,4℃过夜,将条带放入含有辣根过氧化酶标记的二抗(1∶10 000) 孵育液中2 h。最后用化学发光法显色,在BioRad蛋白成像仪成像,并用Image J软件测灰度值进行半定量分析。
1.4雌激素对C2C12增殖的影响 C2C12传代后,在增殖培养基中培养至50%融合时,分为对照组、10-9mol/L E2组、10-8mol/L E2组、10-7mol/L E2组、10-6mol/L E2组及10-8mol/L E2+ICI182,780组,分别加入雌激素0 mol/L、10-9mol/L、10-8mol/L、10-7mol/L、10-6mol/L及10-8mol/L +ICI182,780,每隔1日换液1次,培养5 d。采用MTT比色法检测细胞增殖:取各组细胞,于37℃、5%CO2培养箱中分别培养1、2、3、4、5 d后,每孔加入10 μl MTT,孵育4 h。在全自动酶标仪上检测570 nm光密度值(OD值),评价C2C12增殖情况。
1.5雌激素对C2C12迁移的影响 细胞分组及干预方法同1.4,培养48 h。Transwell小室放置于预先准备好的 24孔板中,按上述条件培养箱中过夜,向Transwell小室下室中加入600 μl含10%胎牛血清的DMEM高糖干细胞培养基。用无血清的DMEM高糖培养液重悬各组细胞,取200 μl细胞悬液(细胞总数约1×105个)加至Transwell上室。48 h后去除下室中的培养液,将小室置于4%多聚甲醛中固定15 min,擦去小室内室膜上的细胞,结晶紫溶液(0.1%)染色 10 min,磷酸盐缓冲液(PBS)漂洗后,显微镜下观察,每孔随机选取5个视野拍照,计数统计并分析。
1.6雌激素对C2C12分化的影响 2% HS诱导分化培养条件下,细胞分组方法同1.4。每隔1 d换液1次,培养4 d。①免疫荧光化学染色法:随机选取5个视野,在MYHC荧光染色背景下计数核融合≥3的肌管形成数目。②Western印迹:取 C2C12,经RIPA裂解、离心收蛋白,行BCA法进行蛋白定量。按分组各上50 μg蛋白,经10%SDS-PAGE电泳后,转PVDF膜。用TBST配制的5%的脱脂奶粉低速摇床封闭40 min,分别放入含有α-Tubulin、MYHC、MyoG等不同一抗的封闭袋中,4℃过夜,将条带放入含有辣根过氧化酶标记的二抗孵育液中2 h。最后用化学发光法显色,在BioRad蛋白成像仪成像,并用Image J软件测灰度值进行半定量分析。
1.7统计学方法 采用SPSS22.0软件进行方差分析、t检验。
2 结 果
2.1E2受体表达 对照组及不同浓度E2组均有ERα和ERβ表达,见图1。进一步半定量分析显示随着E2浓度升高ERα表达也随之升高,当E2浓度为10-6mol/L时,其表达量达最高值(P<0.05),且10-7、10-6mol/L E2组与对照组差异有统计学意义(P<0.05);而随着E2浓度升高,ERβ在C2C12表达量呈现出先升高再下降的趋势,在E2浓度为10-8mol/L时表达量最高(P<0.05),与对照组差异有统计学意义。见表1。
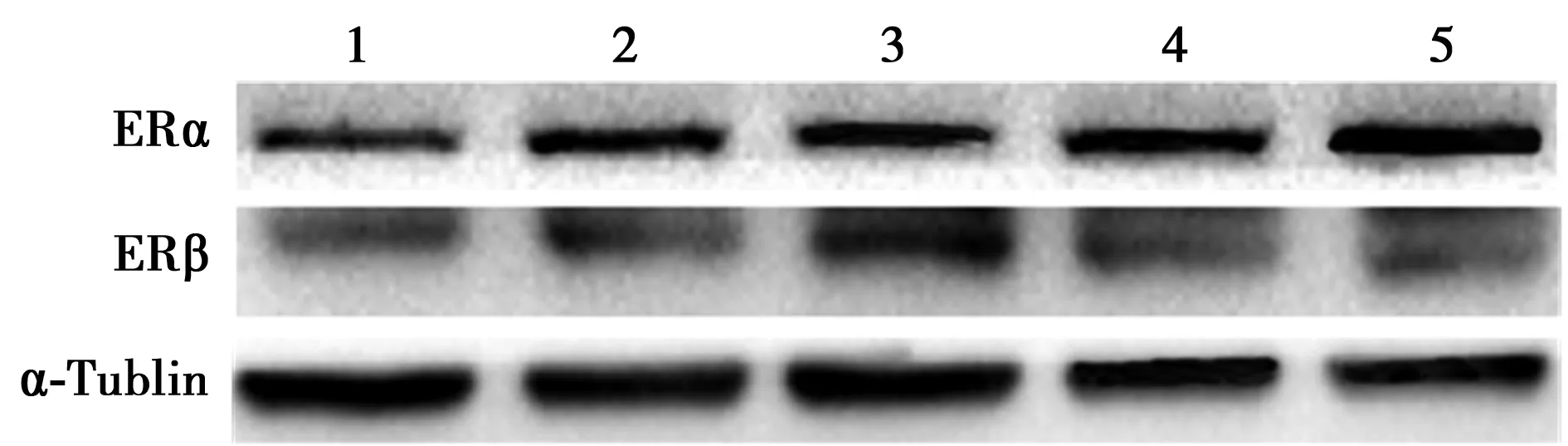
1~5:对照组、10-9 mol/L E2组、10-8 mol/L E2组、10-7 mol/L E2组、10-6 mol/L E2组图1 Western印迹检测不同浓度E2条件下C2C12雌激素受体表达
表1 Western印迹检测ERα及ERβ表达相对值
2.2细胞增殖情况 第3天开始10-8mol/L E2组细胞增殖显著高于对照组,且10-8mol/L E2+ ICI182,178组细胞增殖显著低于10-8mol/L E2组(P<0.05)。见表2。
2.3各组细胞分化情况 与对照组相比,不同浓度E2组肌管形成数均显著升高(P<0.05),以10-8mol/L E2组促进C2C12肌管形成最为明显,即在E2浓度≤10-8mol/L时MYHC表达及肌管形成数目均随着E2浓度升高而增多,但当E2浓度>10-8mol/L时,却呈现逐渐下降的趋势,给予E2受体拮抗剂ICI182,780处理后,肌管形成数目均显著低于10-8mol/L E2组(P<0.05)。见图2,表3。
2.4各组细胞迁移情况 不同浓度E2组细胞迁移数虽然均高于对照组,但并非随着E2浓度升高出现持续升高,而呈现出先升高后下降的变化趋势。E2浓度≤10-8mol/L时,细胞迁移随着E2浓度升高而增多;E2浓度>10-8mol/L时,随着E2浓度升高,细胞迁移数逐渐下降,即10-8mol/L浓度的E2对C2C12细胞迁移促进作用最明显,而给予E2受体拮抗剂ICI182,780处理后,迁移细胞数再次升高,但仍显著高于对照组且显著低于10-8mol/L E2组(P<0.05)。见表3、图3。
表2 不同浓度E2对C2C12细胞增殖的影响

红色代表MYHC染色阳性,蓝色代表DAPI染色细胞核图2 不同浓度E2对分化条件下C2C12细胞MYHC的表达(免疫荧光,×200)
表3 不同浓度雌激素组C2C12细胞迁移数、肌管形成数及MYHC和MyoG水平比较
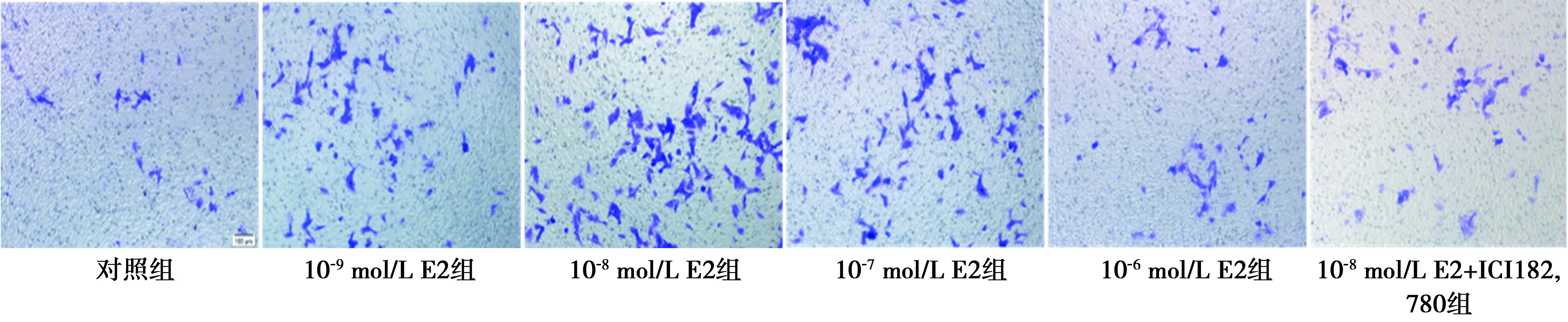
图3 不同浓度E2对C2C12迁移的影响(Transwell法结合结晶紫染色,×100)
2.5分化状态C2C12 MYHC和MyoG表达 C2C12中MyoG的表达变化与MYHC变化规律一致,在E2浓度≤10-8mol/L时MYHC和MyoG表达均随着E2浓度升高而增多,但当E2浓度>10-8mol/L时,却均出现逐渐下降的趋势,即10-8mol/L组MYHC及MyoG表达量最高(P<0.05)。半定量分析也呈现出同样的结果,且统计学分析发现10-8mol/L组MYHC和MyoG的表达量与对照组差异有统计学意义(P<0.05)。给予E2受体拮抗剂ICI182,780处理后,MyoG及MYHC水平均显著低于10-8mol/L E2组(P<0.05)。见表3,图4。

1~6:对照组、10-9 mol/L E2组、10-8 mol/L E2组、10-7 mol/L E2组、10-6 mol/L E2组、10-8 mol/L E2+ICI182,780组图4 Western印迹检测E2对分化状态下C2C12 MYHC和MyoG表达的影响
3 讨 论
骨骼肌分化是一个高度有组织的过程,涉及到多种基因的调节〔8,9〕,特别是在成肌细胞形成肌管的过程中,多种激素和细胞因子参与其中〔10~12〕。雌激素受体是雌激素发挥生物学效应的必要因素,本研究结果表明雌激素促进C2C12细胞增殖是通过与雌激素受体结合发挥作用的,且其浓度依赖性效应与雌激素受体α数量上调有关,这一结论与Ihionkhan等〔13〕报道内皮细胞ERα的激活依赖于雌激素,其表达水平受雌激素的调控相一致。本研究结果表明,雌激素对C2C12细胞的分化效应是通过与雌激素受体结合后促进MyoG表达来实现的。这一研究结果与Scully等〔14〕研究相吻合。
本研究发现雌激素抑制了C2C12细胞的迁移,且随着雌激素浓度升高,抑制作用增强。结合Deng等〔15〕的研究发现,雌激素抑制了肝卫星细胞的迁移,且这一效应与雌激素受体β的表达下降相关,本研究结果表明雌激素浓度依赖性抑制C2C12细胞迁移与ERβ下降有关。
