罗非鱼多聚免疫球蛋白受体的生物学特性及对罗非鱼湖病毒增殖的影响
2022-07-29郑树城曾伟伟李莹莹尹纪元王雅慧吕月凤石存斌赵志英姜志勇王英英
李 波,郑树城,曾伟伟,王 庆,李莹莹,尹纪元,王雅慧,吕月凤,石存斌,赵志英,姜志勇,王英英
(1.上海海洋大学水产与生命科学学院,上海,201306;2.中国水产科学研究院珠江水产研究所,农业农村部渔用药物创制重点实验室,广东省水产动物免疫技术重点实验室,广州,510385;3.佛山科学技术学院,动物分子设计与精准育种重点实验室,普通高校动物分子设计与精准育种重点实验室,生命科学与工程学院,广东佛山,440605; 4.厦门大学海洋与地球学院, 福建厦门, 361100;5.海南省海洋与渔业科学院,海口,571126;6.广东省农业技术推广中心,广州,510030)
多聚免疫球蛋白受体是一种Ⅰ型跨膜糖蛋白,能够转运分泌型抗体抑制病原微生物黏附和入侵。哺乳动物的多聚免疫球蛋白(polymeric immunoglobulin,pIg)R分子由胞质区,胞外配体结合区和跨膜区三部分构成,其中胞外配体结合区含有5个免疫球蛋白样功能结构域(ILD1~ILD5),其中ILD1是与pIg结合的重要结构域,与其它脊椎动物中ILD的数量不同,鱼类pIgR分子只发现有对应哺乳动物ILD1和ILD5的两个免疫球蛋白样结构域。
目前pIgR的免疫学功能研究主要集中在粘膜上皮细胞中pIg的调节、与配体的结合以及对pIg的胞内转运等方面。在鱼类研究上则主要涉及pIgR在病原感染过程中的调控作用。研究发现,泥鳅()、鲫()、鲤()、斑马鱼()、草鱼()以及大菱鲆()等鱼类感染细菌后的pIgR表达水平相较健康的鱼体具有显著性差异。虹鳟()以及海鲈()体内的pIgR通过与其共生菌和病原菌相结合而发挥抑制病原菌增殖的作用,表明pIgR在维持微生物群落以及抵御黏膜表面病原菌入侵方面发挥着重要作用。白斑综合征病毒(white spot syndrome virus,WSSV)利用pIgR作为感染日本囊对虾()的细胞表面受体,其胞外部分能够与WSSV的VP24互作,胞内部分则与钙调素作用募集网格蛋白和AP-2复合物促进WSSV的侵入。
本研究通过PCR技术对罗非鱼pIgR基因R完整的开放阅读框序列进行扩增,预测了罗非鱼pIgR的蛋白结构,构建系统发育树,检测罗非鱼感染罗非鱼湖病毒(TiLV)后脾、肾以及皮肤组织中pIgR表达的变化,构建了重组真核表达载体pEGFP-R并将重组质粒转染至罗非鱼脑细胞(TiB)中,初步研究了罗非鱼pIgR对TiLV增殖的影响, 为罗非鱼的抗病毒免疫研究提供理论基础。
1 材料方法
1.1 实验材料
实验采用的健康罗非鱼,平均体重(20±5) g,采购自广东省某养殖场,暂养于120 cm×50 cm×60 cm的玻璃鱼缸中,水温维持在(28±1)℃且持续曝气充氧,定时定量投喂。表达载体pEGFP-N1和克隆宿主5购自北京天根生物科技有限公司;TiB由珠江水产研究所鱼病室建立并保存; TiLV-2017A由Sven.M.Bergennman博士(德国动物健康研究院) 赠送;限制性内切酶EcoRI和KpnI、15 000 DNA marker、反转录试剂盒等购于宝生物工程有限公司;RNA提取试剂盒、质粒提取试剂盒和胶回收试剂盒购自Omega生物技术有限公司;荧光定量试剂盒购自艾克瑞生物工程有限公司。
1.2 总RNA的提取以及cDNA的合成
取健康罗非鱼18尾。15尾用于攻毒,每尾腹腔注射200 μL浓度约为10copies/μL的TiLV,剩余3尾注射相同体积PBS作为对照。攻毒组在注射后的12、24、48、72、96 h参照文献[4]分别取三尾鱼的脾脏、肾脏、皮肤等组织进行后续研究。使用Total RNA Kit I从收集的样品中提取总RNA。经1%琼脂糖凝胶电泳和NANODROP 2000分光光度计确定RNA质量与浓度。参照PrimeScriptRT Reagent Kit反转录试剂盒说明书合成cDNA,-20 ℃备用。
1.3 OnpIgR基因的克隆
根据NCBI上提供的pIgR参考序列(GenBank:MK061423.1),设计R基因ORF序列特异性扩增引物。引物名称及序列如表1所示。 以1.2步骤中获得的cDNA为模板使用高保真酶Prime STAR MAX,按照50 μL反应体系配制反应液,即Prime STAR MAX 25 μL,R-EcoRI-F,R-KpnI-R各1 μL,cDNA 2 μL,无酶水21 μL;反应条件为98 ℃ 10 s,55 ℃ 15 s,72 ℃ 10 s,经35次循环后获得ORF全长。PCR产物经1%的凝胶电泳鉴定后,在1 023 bp左右出现单一的目的条带,采用Gel Extraction Kit(广州飞扬生物工程有限公司,中国)对PCR产物进行纯化。将纯化产物连接至pMD18T载体,转化到DH5α感受态细胞内,使用氨苄抗性平板进行筛选,经PCR检测阳性后,送生工生物工程股份有限公司测序。
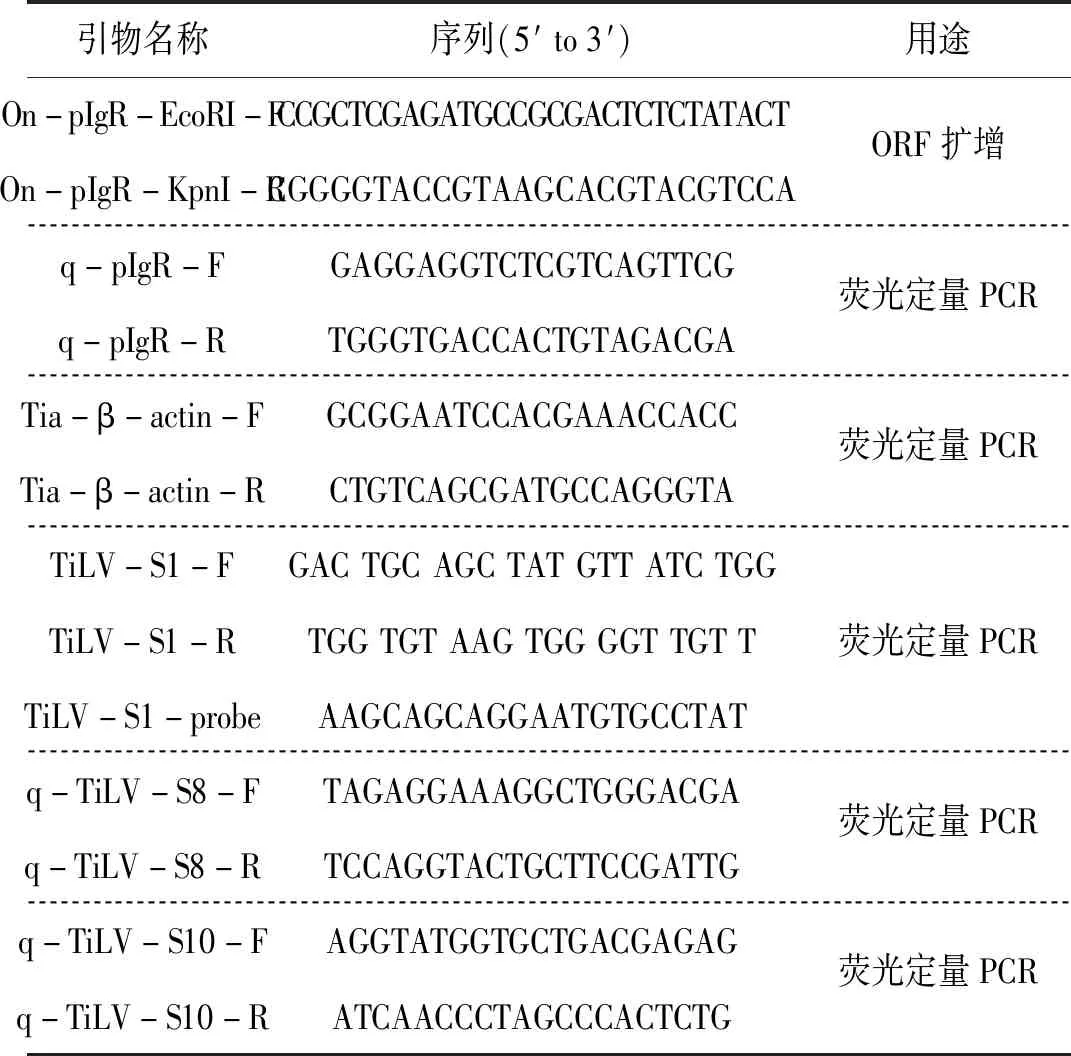
表1 引物信息Tab.1 Primers information
1.4 OnpIgR生物信息学分析
采用SMART在线服务器(https://smart.embl-heidelberg.de/smart/set_mode.cgi?NORMAL=1)预测R的结构域。通过I-tasser在线服务器(https://zhanggroup.org/I-TASSER/)预测R的三维结构。使用Cluster-X2.0程序进行多物种pIgR氨基酸序列比对,使用MEGA-X软件进行系统发生和进化分析,采用邻位相连法构建系统进化树。
1.5 定量分析TiLV感染后组织中pIgR的表达变化
根据得到的ORF序列设计了实时荧光定量引物(q-pIgR-F/q-pIgR-R),以β-actin(Tia-β-actin-F/ Tia-β-actin-R)作为内参基因(序列见表1),按照SYBR Green Premix ProHS qPCR Kit 说明书对1.2步骤中所取各组织中的pIgR进行定量分析。最后根据所测得的Ct值,采用2法计算pIgR基因的相对表达量。
1.6 真核表达载体的构建
使用限制性内切酶EcoRI和KpnI对扩增获得的R基因以及pEGFP-N1载体进行酶切,反应条件为37 ℃,1 h。使用Thermo Fisher 的T4连接酶对酶切产物进行连接,连接条件为22 ℃ 10 min。将连接产物转化后,涂布于卡那霉素抗性平板上筛选鉴定,阳性克隆经PCR以及酶切鉴定后送测序。序列正确的质粒命名为pEGFP-R。使用Omega公司的无内毒素的质粒提取试剂盒提取重组质粒pEGFP-R,测定质粒浓度后-20 ℃保存。
1.7 Western Blot鉴定
将状态良好的TiB细胞经胰酶消化处理后,用含10% 胎牛血清(FBS)的M199培养基制备成含有1×10个/mL的细胞悬液,分别吸取2.5 mL的细胞悬液接种到六孔板的三个孔中,待细胞单层覆盖到80%~90%左右的时候,分别向其中的两孔中转染2 μg的重组质粒pEGFP-R以及pEGFP-N1空载,剩下一孔不做转染。转染方法参照Lipo-fectamine2000(Invitrogen,USA)的说明书进行。转染36 h后,弃上清,加入200 μL 1× SDS缓冲液收取细胞样到1.5 mL的离心管中,混匀后煮沸5 min,12 000离心1 min获得上清。蛋白样品经SDS-PAGE电泳分离后,转移到硝酸纤维素膜(NC膜)上进行Western blot分析。5%脱脂奶室温下封闭1 h,将商品化GFP兔抗按照1∶4 000倍稀释后室温孵育2 h,加入二抗(1∶4 000稀释)室温孵育2 h,加入显色液显色,拍照记录。
1.8 TiB细胞中OnpIgR过表达对TiLV增殖的影响
将状态良好的TiB细胞于转染前24 h接种到底面积为25 cm的6个细胞培养瓶中,平均分为2组。 每一组参照Lipofectamine2000说明书步骤分别转染50 μg的pEGFP-N1质粒和pEGFP-R过表达质粒,每组设置三个平行。并在转染后的36 h时弃掉培养液,用HBSS清洗两遍,分别加入浓度约为10copies/μL 的TiLV病毒液各1 mL,在28 ℃的培养箱中孵育1.5 h,之后弃掉病毒液,补加5 mL的含5%FBS的M199培养基,继续放入28 ℃培养箱中培养。在病毒感染后的48 h和72 h收样。分别对-1基因,-8和-10基因进行定量检测。
1.9 数据分析
使用GraphPad Prism 8 软件对实验组与对照组的数据按照tests方法进行显著性差异分析,>005表示差异不显著,<005表示差异显著,<0.01表示差异极显著。
2 结果
2.1 罗非鱼pIgR分子特征
通过PCR扩增的基因经测序比对后,确认为罗非鱼pIgR 完整ORF 序列。结构域预测显示,罗非鱼pIgR蛋白的第1~20个氨基酸构成一个信号肽,第25~130以及第137~229个氨基酸分别构成两个Ig样结构域(图1-A);同源建模结果显示,pIgR蛋白整体呈扭曲的“L”型,主要由反平行β片层和无规卷曲组成(图1-B);进化树分析显示,pIgR主要分为哺乳类,两栖类和鸟类,以及硬骨鱼类三个亚群。其中罗非鱼pIgR与同属于鲈形目()的大口黑鲈()以及斜带石斑鱼()的pIgR最为接近(图2)。
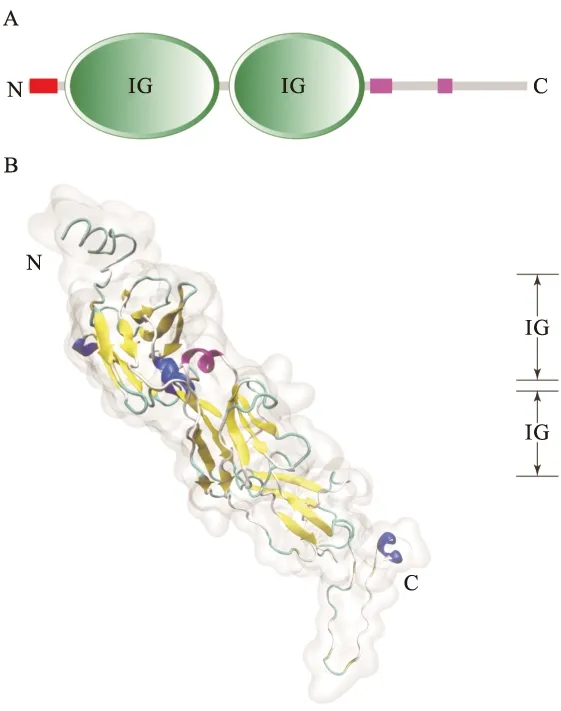
图1 罗非鱼pIgR蛋白结构域与三维结构预测Fig.1 Prediction of tilapia pIgR protein domain and 3D structureA结构域预测:红色区域为信号肽,绿色区域为Ig样结构域,紫色区域为低复杂度区;B:三维结构预测

图2 不同物种pIgR蛋白序列系统进化树Fig.2 Phylogenetic tree of different species pIgR amino acids sequences
2.2 罗非鱼pIgR时相表达模式分析
罗非鱼在感染TiLV后,皮肤、脾脏以及肾脏三个组织器官中pIgR的表达量均呈现先上升后下降的趋势,感染后24 h皮肤中pIgR的表达量达到最高值(图3-B);在脾脏和肾脏中则是48 h时表达量达到最高值随后开始下降(图3-A、3-C)。
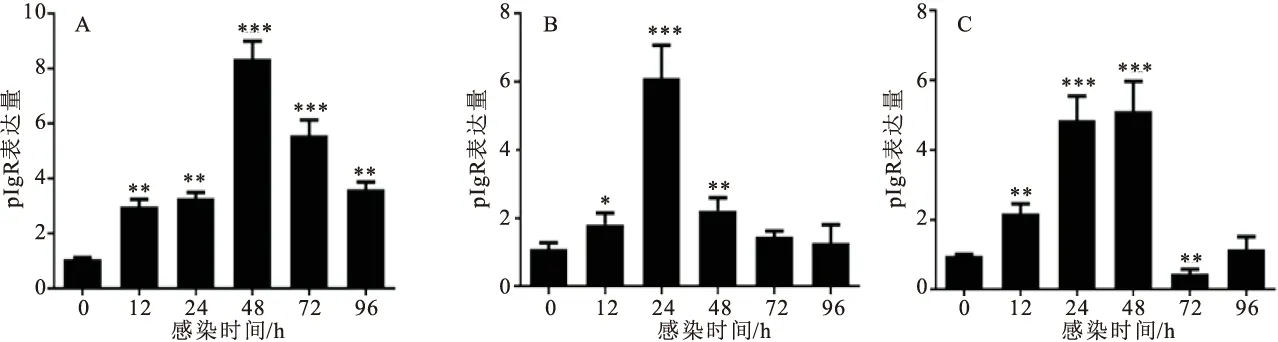
图3 不同组织中的时相表达Fig.3 pIgR expression in different tissues at post infectionA:pIgR在脾脏中的时相;B:pIgR在皮肤中的时相;C:pIgR在肾脏中的时相;*表示差异存在显著性(P<0.05),图5同。
2.3 重组真核表达载体构建与鉴定
结果显示转染pEGFP-R在55~70 kDa的位置出现与预期大小相符的融合蛋白条带(图4),表明重组真核表达质粒pEGFP-R可以在细胞内正确表达出目的蛋白。
图4 Western-blot鉴定重组质粒pEGFP-OnpIgR体外表达情况Fig.4 Expression of recombinant plasmid pEGFP-OnpIgR in vitroM为蛋白Marker;1为转染重组质粒细胞样;2为转染pEGFP-n1空载细胞样;3为空白TiB细胞样
2.4 pIgR过表达对TiLV增殖的影响
图5结果显示,与对照组细胞相比,在过表达pIgR的细胞中TiLV感染后48 h和72 h的-8和-10片段的mRNA水平均明显上调,病毒拷贝数也显著增多,表明pIgR显著促进了TiLV的增殖。
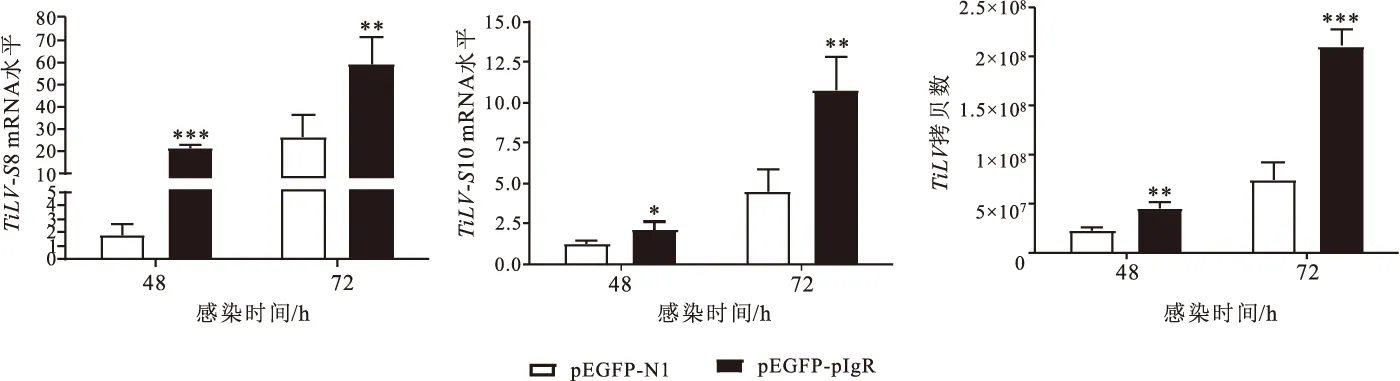
图5 pIgR过表达对TiLV增殖的影响Fig.5 Effect of pIgR overexpression on TilV proliferation
3 讨论
本研究成功克隆了罗非鱼pIgR 完整ORF序列,长度为1 023 bp,蛋白结构域预测显示罗非鱼pIgR蛋白具有一个信号肽,两个Ig样结构域,同源建模结果显示,pIgR蛋白整体呈扭曲的“L”型,主要由反平行β片层和无规卷曲组成,与相关报道中的鱼类pIgR结构一致,表明罗非鱼的pIgR具备结合和转运多聚免疫球蛋白的结构基础;系统进化树结果显示,硬骨鱼类pIgR聚为一支,两栖类和鸟类的聚为一支,哺乳类的聚为一支,且罗非鱼pIgR与同属于鲈形目的大口黑鲈以及斜带石斑鱼的pIgR聚为一个分支,而同属于鲤科鱼类的草鱼、鲤鱼以及斑马鱼又聚为一个分支,表明pIgR具有一定的种属性差异。RT-qPCR定量结果显示,感染TiLV后罗非鱼皮肤、脾脏以及肾脏器官中pIgR的表达量均呈现先上升后下降的趋势,感染后24 h皮肤中表达量达到最高值,随后开始下降;在脾脏和肾脏中则是48 h时达到最高值随后开始下降。表明罗非鱼受到外界刺激时,相对于中枢免疫器官而言,粘液相关免疫组织能够更早地做出免疫反应。成功构建了罗非鱼pIgR过表达受体,并在TiB细胞中进行pIgR过表达后显著促进了TiLV的增殖。与NIU等的发现,WSSV能够利用pIgR促进自身增殖;在LLC-PK1细胞上过表达也能够促进猪流行性腹泻病毒(PEDV)增殖等研究结果一致,初步表明TiLV也可能利用pIgR促进自身感染。
目前关于鱼类pIgR 的研究大多涉及对病原菌的响应和表达模式分析。如pIgR在斜带石斑鱼、牙鲆()、大菱鲆和鲤鱼等硬骨鱼类组织中均有表达,且在胃、肠道、鳃和皮肤等粘液相关组织中的表达量较高。通过实时荧光定量检测发现菌液浸泡处理后的鲤鱼肠道中的pIgR表达量呈先上升后下降的趋势。斑马鱼在感染海豚链球菌后,pIgR表达水平也呈现稳定上升趋势,但是感染乌鳢弹状病毒(SHRV)后的斑马鱼pIgR表达水平则稳定下降。表明病毒刺激引起的pIgR表达变化与细菌刺激引起的变化存在着差异,具体原因有待后续进一步研究证明。草鱼浸泡感染柱状黄杆菌后,pIgR mRNA水平也迅速显著上升,随后逐渐下降到对照水平,并且在草鱼的皮肤中pIgR mRNA的增长最早,幅度最大,且其在皮肤中的应答反应比在肠或鳃中更快。除了病原菌,有报道表明泥鳅在感染了白点虫之后,其脾脏、肾脏、鳃和皮肤中的pIgR表达量也显著升高。尽管对多种不同硬骨鱼类pIgR在应对病原菌或寄生虫感染响应的表达模式有了初步的了解,并提示其在黏膜免疫过程中发挥重要的作用,但其潜在的分子机制仍有待进一步阐明。
除了黏膜免疫上的功能,pIgR作为细胞表面分子受体也常被部分病毒劫持作为入侵的靶点。之前的研究已发现多种病毒包括RNA 病毒和DNA病毒将pIgR作为病毒入侵的受体进入细胞。如pIgR过表达的宿主细胞感染PEDV之后,PEDV的N蛋白、病毒滴度以及病毒mRNA均显著增加,表明pIgR促进了PEDV在宿主细胞内的增殖。王会英等发现,EPC细胞中干扰素的特异性表达能够靶向抑制传染性造血器官坏死病毒的增殖。而pIgR表达量的降低能够导致IFN-γ表达水平的上升,从而减低鼠诺如病毒(MNV)的滴度,在pIgR敲降小鼠中,病毒的急性感染也出现减缓。以上研究均表明pIgR在影响病毒入侵过程中发挥着重要作用。而关于水生动物pIgR及病毒的研究,目前较为清晰的是日本囊对虾pIgR(MjpIgR)与WSSV之间的互作,该病毒可通过与MjpIgR结合后经网格蛋白介导的内吞方式进入细胞完成自身增殖。本研究发现罗非鱼pIgR过表达后也促进了TiLV的增殖,TiLV是否也是利用pIgR通过网格蛋白介导的内吞进入细胞将在后续的研究中进一步阐明。
作为水生动物新发病毒,TiLV的病原学及致病机理仍不清楚,本研究对罗非鱼pIgR的生物学特性及对TiLV的增值影响作用进行了初步研究,为后续TiLV致病机制的研究、抗TiLV药物的研发和抗TiLV苗种的培育具有一定的借鉴意义。
