NLRP3位点rs12048215多态性对哮喘儿童外周血嗜酸粒细胞的影响
2022-07-28蔡旭龙尹同进徐巧岚
蔡旭龙 尹同进 徐巧岚
支气管哮喘(简称哮喘)是儿童常见的慢性呼吸道疾病,目前发病机制尚不明确,主要由环境因素和遗传因素共同作用引起,其病理特点为慢性气道炎症[1]。研究发现哮喘主要由Th2细胞因子IL-4、IL-5和IL-13驱动,导致气道嗜酸性粒细胞炎症[2]。NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor containing pyrin domain 3,NLRP3)是模式识别受体家族NOD样受体中的一员,参与Th2细胞的分化[3]。NLRP3是组成NLRP3炎症小体的重要部分,使哮喘儿童外周血中,下游因子IL-1β和IL-18水平升高,与哮喘疾病进程关系密切[4]。目前,NLRP3基因多态性是否影响哮喘的易感性及其表型的研究鲜有报道。本研究分析NLRP3基因位点多态性在哮喘患儿和健康儿童中的分布差异,探讨基因多态性在哮喘发病中的意义。
资料与方法
一、研究对象
2020年8月至2021年6月期间在盐城市第三人民医院就诊治疗的哮喘儿童,收集急性发作期的患儿130例,其中女性57例,男性73例,年龄3~14岁,诊断哮喘符合《儿童支气管哮喘诊断与防治指南(2016年版)》[5]。同期收集80例健康儿童,其中女性36例,男性44例,年龄3~14岁,无哮喘、过敏性鼻炎、特应性皮炎史,无遗传免疫缺陷病史,无过敏史,家族中三代以内无哮喘史。与纳入研究儿童家长签订知情同意书,同时本次研究获得医院伦理委员会批准(批准文号:2020-55)。
二、标本采集及DNA提取
空腹抽取外周静脉血2mL于EDTA抗凝管中。DNA的提取严格按照人血DNA提取试剂盒(生工生物工程(上海)股份有限公司)说明操作。提取的DNA标本存放-80℃冰箱。
三、基因目标位点筛选
从“千人基因组计划”项目数据库中下载中国汉族健康人群NLRP3基因多态性位点数据(http://grch37.ensembl.org/index.html)。将获得的数据导入Haploview,设计分析的最小等位基因频率大于20%,依据连锁不平衡获得5个Block区,每个区筛选1个位点,共5个目标位点(rs12137901、rs4998423、rs12048215、rs1539019、rs12130711)(见表1)。
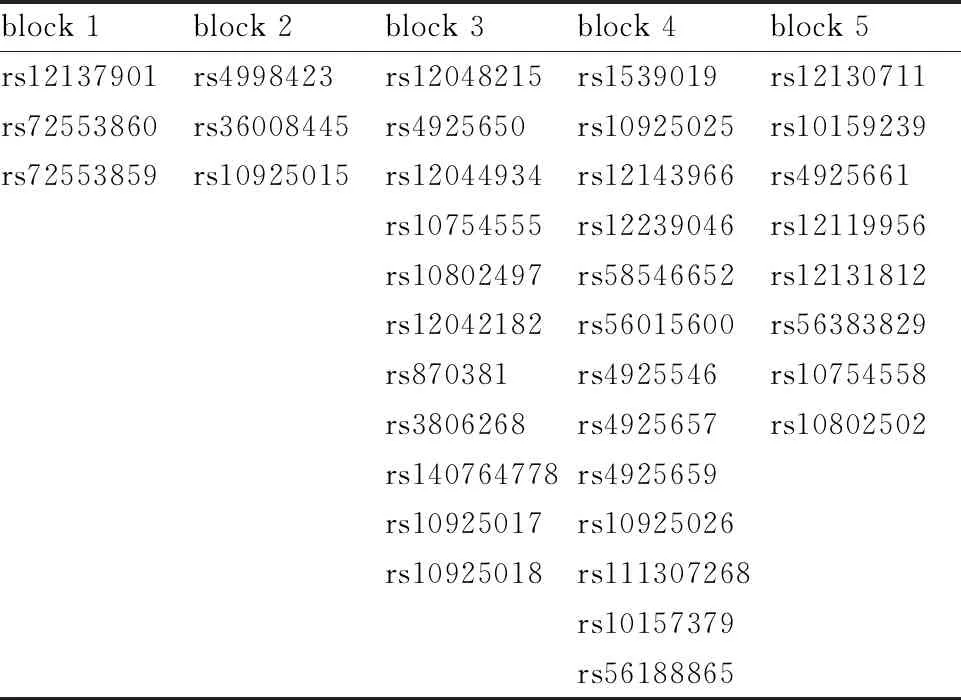
表1 NLRP3多态性位点在不同block区分布情况
四、多重扩增及高通量测序基因分型
设计并合成好一个包含5个SNP位点的引物池(见表2),然后通过两步PCR的方法完成目标SNP位点的扩增和兼容illumina测序文库的制备。第一轮PCR体系如下:DNA模板(10 ng/μL)2μL;上游引物池(10μM)1μL;下游引物池(10μM)1μL;2×PCR Ready Mix 15μl (总体积 25μL)(Kapa HiFi Ready Mix)。配制好反应体系后,在PCR仪(BIO-RAD,T100TM)执行反应程序:98℃预变性3分钟,然后执行8个循环(条件:98℃变性30秒,50℃退火30秒,72℃延伸30秒)。紧接着执行25个循环(条件:98℃变性30秒,66℃退火30秒,72℃延伸30秒)。最后72℃延伸5分钟。反应完成后,维持4℃。

表2 NLRP3多态性位点的引物
然后以第一轮PCR产物为模板执行第二轮PCR反应,以获得测序带分子标签的文库。反应体系如下:DNA模板(10ng/μL)2μL,通用P7引物(含分子标签,10μM)1μL; 通用P5引物(10μM)1μL; 2×PCR Ready Mix15μL (总体积30μL)。配制好反应体系后,执行如下PCR程序:98℃预变性5分钟,然后执行5个循环程序(变性94℃30秒,55℃退火20秒,72℃延伸30秒)。最终72℃延伸5分钟。完成后维持4℃。最终PCR产物使用AMPure XP磁珠纯化回收。各个PCR产物等量混合后,使用HiSeq XTen测序仪(Illumina, San Diego, CA)进行测序。
使用BWA(v 0.7.13-r1126)软件将质控合格的序列比对到参考基因组上,参数为默认参数。根据比对结果,通过samtools软件(版本0.1.18)计算目标位点的基因型结果基因分型。
五、统计学方法
结 果
一、研究对象的一般特征
本次研究纳入哮喘儿童130例,健康儿童80例,通过病例对照研究进行分析。年龄、性别构成在两组间无统计学差异。总IgE、白细胞、嗜酸粒细胞百分比在哮喘组中均明显高于健康组,P<0.001。哮喘组中有过敏史的占60.7%,过敏性鼻炎占42.3%,特应性皮炎的占29.2%,有哮喘家族史的占18.4%(见表3)。

表3 研究对象一般资料
二、多态性位点基因型及等位基因在哮喘和健康儿童中的分布情况
通过Haploview分析获得NLRP3基因Tag位点rs12137901、rs4998423、rs12048215、rs1539019、rs12130711,并进行基因分型(见表4)。5个多态性位点的基因型及等位基因在哮喘和健康儿童中分布无统计学差异。
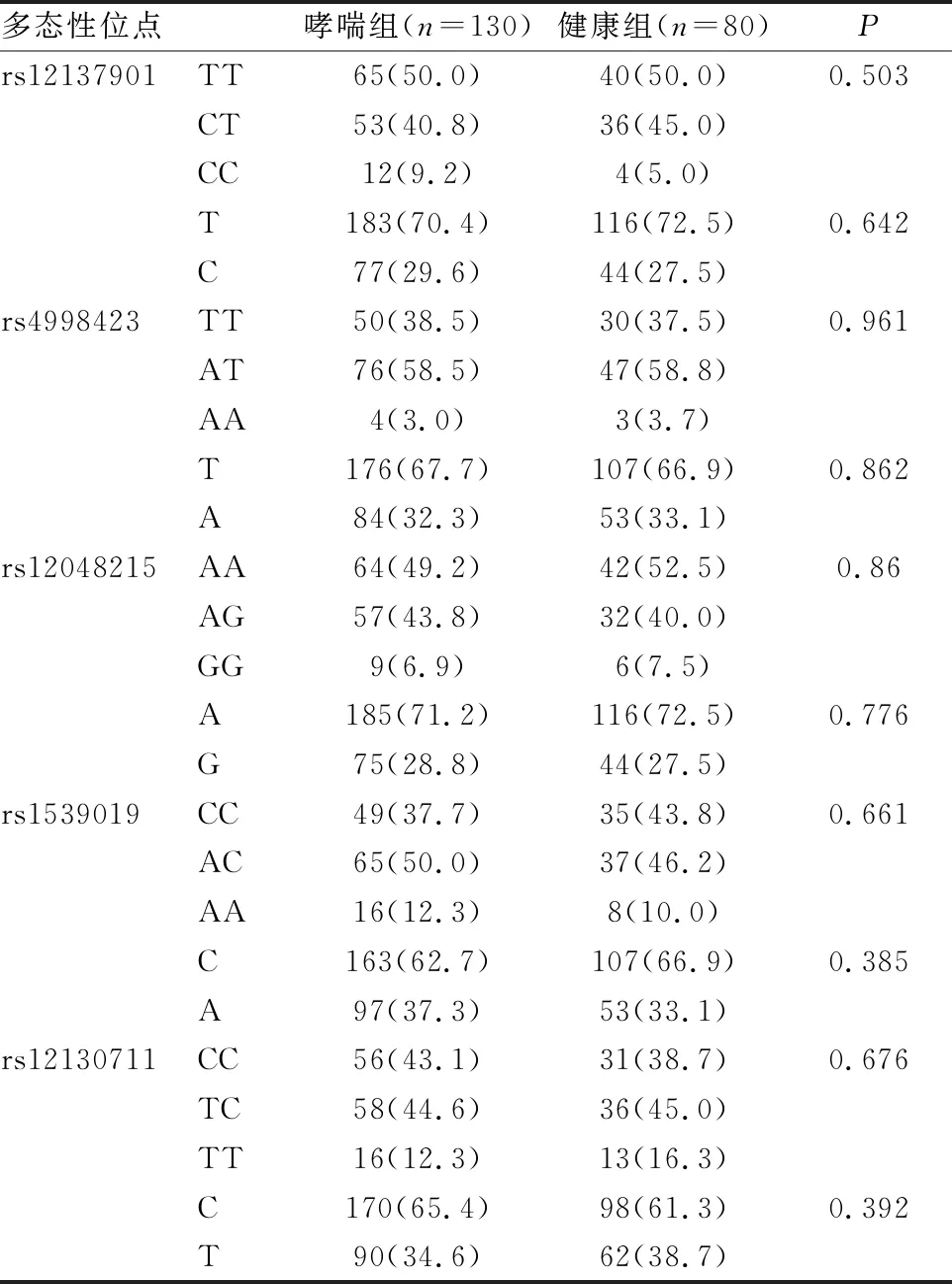
表4 NLRP3基因位点多态性在哮喘和健康儿童中的分布[n(%)]
三、不同基因型在嗜酸粒细胞表型中的分布情况
取嗜酸粒细胞比例3%为分界,将哮喘儿童分两组:嗜酸粒细胞%>3%(n1=63),嗜酸粒细胞%≤3%(n2=67),并进行基因型分布统计分析(见表5)。rs12048215位点有AA、AG、GG三种基因型。嗜酸粒细胞%>3%组中构成比为AA(60.3%)、AG(33.3%)、GG(6.4%)。嗜酸粒细胞%≤3%组中构成比为AA(38.8%)、AG(53.7%)、GG(7.5%)。rs12048215位点基因型在两组不同比例嗜酸粒细胞中分布有差异,P=0.045。共显性模型AA vs (AG+GG)在两组中的分布比较(P=0.014,OR [95%CI]=2.397[1.185~4.848])。rs12048215位点等位基因A、G在两组中有统计学差异(P=0.044,OR [95%CI]=1.748[1.012~3.021])。
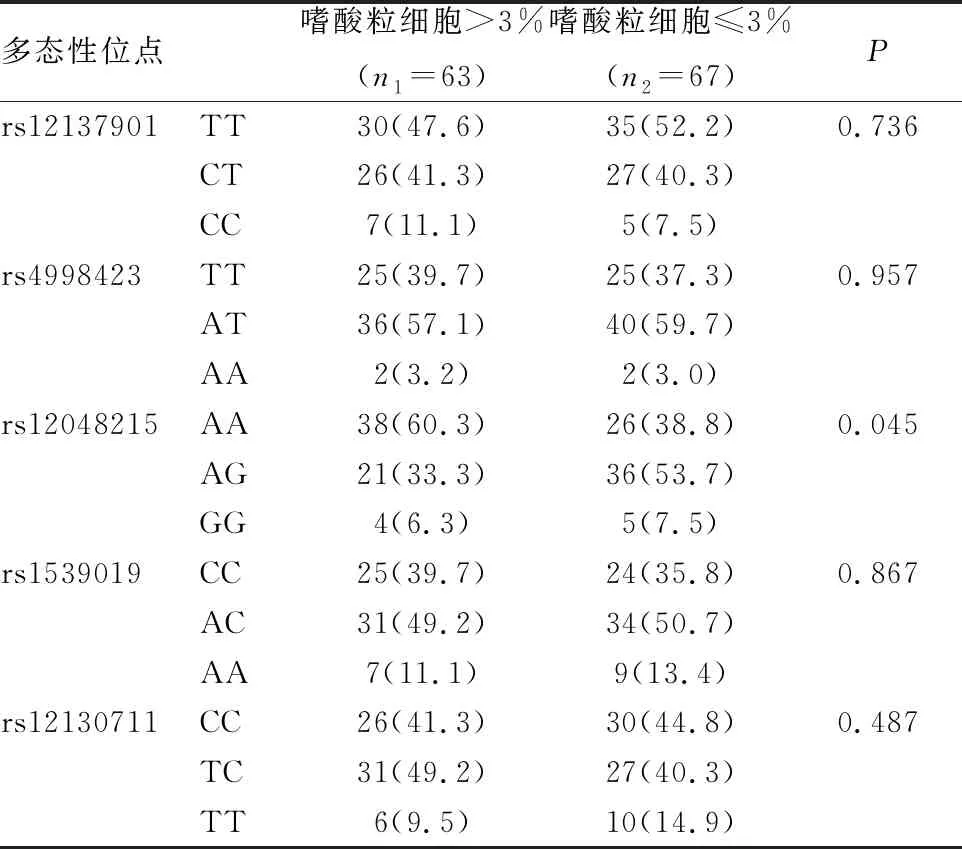
表5 NLRP3基因位点多态性在哮喘儿童不同嗜酸粒细胞比例中的分布[n(%)]
四、rs12048215位点多态性的功能预测
通过3DSNP v2.0预测NLRP3基因rs12048215位点的功能[6](见表6)。分析出rs12048215位点可能影响FOS和STAT3。FOS是AP-1转录因子的亚单位,存在于细胞核质内,参与调节嗜酸粒细胞[7]。STAT3是信号转导和转录激活因子,存在于细胞核质和胞浆中,参与调节嗜酸粒细胞[8]。

表6 与rs12048215位点相关的蛋白因子
讨 论
遗传因素在哮喘发病中起着重要作用,遗传率在35%到95%之间,研究确认存在数百种与哮喘风险增加相关的基因变异[9]。遗传密码翻译方式的表观遗传变异也被证明在哮喘的发展中起作用[10]。目前对哮喘的理解涉及到大量的遗传多样性,这些遗传多样性通过表观遗传和转录因子进行可变的翻译和环境影响,导致慢性气道炎症的组织病理学特征,从而表现出主要的哮喘症状[9]。
病原体和过敏原能够活化NLRP3[11]。活化的NLRP3能够募集PYCARD和Caspase-1,形成NLRP3炎症小体,参与Th2细胞的分化及Th2类型的过敏炎症反应[12-13]。一项在乳清蛋白诱导的过敏性气道炎症小鼠模型中的研究,发现NLRP3参与过敏性气道炎症的过程[14]。有研究证实NLRP3可通过上调IL-4的表达促进M2巨噬细胞极化[15]。因此,NLRP3参与哮喘的发病机制。但本次研究NLRP3的位点的多态性在哮喘组和健康儿童中分布无差异,可能不是哮喘易感性的因素。
富含嗜酸性粒细胞的慢性气道炎症是哮喘的一个重要特征[16]。嗜酸性粒细胞是由CD34+前体细胞产生的终末分化粒细胞。IL-5、CCR3和CD34与受体结合可促进前体细胞分化为嗜酸性粒细胞。基质细胞、肥大细胞和活化T细胞产生IL-5、GM-CSF和IL-3是骨髓中嗜酸性粒细胞生长和成熟的重要组成部分[17]。在哮喘病程进展过程中,嗜酸粒细胞引起气道慢性炎症发挥重要作用[18-19]。
有研究发现在NLRP3缺失的小鼠模型中,HDM诱导的过敏性肺部炎症显示出肺和肺泡灌洗液中嗜酸粒细胞明显增加[20],NLRP3的缺失对嗜酸粒细胞调节失控。本次研究中,NLRP3基因位点rs12048215的基因型AA、AG、GG在嗜酸粒细胞高比例组和低比例组中,分布有明显差异。在哮喘儿童中,基因型AA与嗜酸粒细胞增高表型有相关性。通过基因位点功能的预测,rs12048215位点多态性与FOS和STAT3有关联。而FOS和STAT3均能够对嗜酸粒细胞产生影响。FOS和STAT3的表达可能受到rs12048215位点多态性的影响,进而可能影响嗜酸粒细胞表型。
NLRP3基因rs12048215位点多态性可能是哮喘儿童嗜酸粒细胞增高表型的因素。但本次研究的样本量、基因位点偏少,需要更广泛系统的研究证实位点多态性对哮喘表型的影响。
