亚热带喀斯特地区不同地形植物群落物种多度分布格局
2022-07-28安明态余江洪吴墨栩
田 力, 安明态*, 余江洪, 吴墨栩
( 1. 贵州大学 林学院, 贵阳 550025; 2. 贵州大学 生物多样性与自然保护研究中心, 贵阳 550025 )
自从群落构建的确定性进程的问题被提出以来,针对这一方向的探索一直从未间断,也被认为是群落生态学研究领域的关键方向(Chun et al., 2017)。对于这一过程的研究产生了许多理论与假设,其中最具争议的当属生态位理论与中性理论(Dawn et al., 2017),传统生态位理论提出群落中所有物种得以共存主要依赖于物种生态位的分化,是物种及其所处环境互相影响的结果(柴永福和岳明,2016),但在研究过程中,某些物种生态位的生态性状产生相似性的重叠影响了研究成效(Ning et al., 2020),因此适用性受到质疑。群落中性理论的提出者Hubbell等人则认为群落中各物种不存在生态功能的差异,随机效应和扩散限制才是控制物种分布方面的主导因素,这一理论在群落构建研究中另辟蹊径,为传统生态位观点的不足提供了较为完善的补充说明,但是中性理论过度强调随机性假设而忽略确定性过程的发掘(刘浩栋等,2020),因此也饱受质疑。尽管过去两种理论在持久的争论中逐渐相互完善,但是都不能完全兼容群落构建中的现象,因此,近年来大多数生态学家逐渐将精力从两种理论的争论转向了两种理论的整合(牛克昌等,2009)。
群落物种的多度格局在群落生态学研究中作为基础特征之一,是揭示物种组成结构、探索构建过程的关键手段,相较于多样性系数量化群落特性的作用,物种多度格局分析更为直观(苏强,2015)。最初物种多度格局用于动物群落结构研究,后引入植物群落,在揭示群落物种多度过程中,不断有新的理论与模型被提出,丰富了群落物种多度分布的研究方法,建立物种多度拟合模型分析群落构建的生态学过程是目前流行的研究方式(臧丽鹏,2019)。模型按照生态学意义可以大致可以分为生态位模型、中性模型与数学纯统计模型三大类,而数学纯统计模型由于其生态学意义未明晰(李全弟等,2019),因此中性模型与生态位模型成为解释物种多度的两大重要模型,代表着两种生态学过程。目前物种多度格局研究聚焦于群落水平和物种水平,由于研究对象的多样性,对于物种多度内容仍在不断扩展,如程佳佳等(2011)研究了物种多度格局对取样尺度的响应表明了模型拟合效果随取样尺度发生变化,再有闫琰等(2012)研究发现物种多度分布也受群落演替阶段的影响,此外,植物生活型(白浩楠等,2021)、稀有种比例(刘旻霞等,2020)等因素也是物种多度分布的控制因素,但物种多度格局的构建是一个复杂的演替进程,其形成过程也与诸多环境因素密不可分。在大尺度上受气候控制,在小尺度上受区域环境的诸多因素干扰,如地形、海拔等(王家鸣等,2018),尤其地形异质性是影响群落物种多度格局重要因素,但是目前对这方面的研究较少,这限制了我们理解群落物种多度格局形成机制与环境的关联。
地形是控制物种多度分布的主要因素(孙永光等,2015),地形异质性会直接影响光照、土壤养分(廖全兰等,2020)、群落特征(赵鹏等,2019)、植物功能性状等(陈晨等,2015),进而影响群落多样性与空间分布特征,因此地形异质性对群落变化影响的研究已有先例,但是这方面的研究多数聚焦于常态地貌,对于喀斯特地形异质性的关注较为薄弱(张忠华等,2011),相较于常态地貌,后者的生境变异较大,地形态势起伏不定,因此与环境资源的分布产生重要联系(黄明芝等,2021),与群落特征的关联性更强。贵州荔波作为中国南方喀斯特地貌主要组成之一,在喀斯特地貌世界地理分布上有重要地位(盛茂银等,2015),该地区溶岩发育典型且强烈,峰丛错综复杂,土壤瘠薄并且临时性干旱频发(罗丝琼等,2020),在这样严酷生境下,经过长期演替,形成了典型的、原始的喀斯特顶极森林群落。该地区物种丰富多样,生理生态特性高度适应当地环境,其独特的群落结构与形成机制一直是中外学者研究的重点,但是当前对该地区研究集中于土壤水热格局,凋落物动态,微生物等(秦随涛等,2018),但是对该地区群落物种多度格局的研究分析较少,而对于该地区地形异质性下物种多度格局的研究更是欠缺,这限制了我们对亚热带喀斯特群落物种多度构建机制的认识,这也限制了我们了解环境因素对于群落物种多度格局的影响。
本研究以贵州茂兰国家级自然保护区喀斯特顶极森林群落为研究区域,依托该地区山脊、槽谷、鞍部、洼地4种典型喀斯特地形下森林群落,采用物种调查与多度拟合的方法,通过Wilcoxon秩和检验、K-S检验、赤池信息准则检验研究结果,拟探讨以下问题:(1) 该地区不同群落主要物种组成、不同地形下物种数与个体数量及形成因素;(2) 4种典型喀斯特地形下群落物种多度格局的差异及影响因素;(3) 不同地形下群落物种多度构建的生态学过程。以期揭示该地区地形异质性下群落物种多度分布特征与物种多度格局形成过程,为亚热带喀斯特森林群落构建过程的研究提供理论基础。
1 研究区概况与方法
1.1 研究区概况
研究开展于贵州省南部的茂兰国家级自然保护区(图1),保护区的经纬度为107°52′—108°05′ E,25°09′—25°20′ N。保护区最高海拔与最低海拔差约600 m,平均海拔800 m左右。该地区的喀斯特溶岩形成强烈,并且该区域温湿适宜,拥有充沛的光水热资源,全年平均温度15.3 ℃,降水量充沛,全年总计达1 750.5 mm。保护区森林植被构成复杂,乔木层物种丰富,林下物种多样,空间利用率高。形成了在地球上同纬度地带少见的喀斯特森林生态系统。

图 1 研究区示意图Fig. 1 Illustration of the study area
1.2 样地设置和物种调查
经考察选中茂兰国家级自然保护区白鹇山附近,该区域植被原生性好并兼具多种地形。参照李宗发(2011)对贵州喀斯特地貌形态特征的界定,结合实际调查,选中山脊、槽谷、鞍部、洼地4种典型喀斯特地形,地形之间间隔大于150 m且均未受人为干扰。样地面积以种-面积曲线为依据,设置样地调查最小面积900 m。按照国家标准《森林生态系统长期定位观测方法》(GB/T 33027—2016)中的相关规范设置样地。每种地形设置3个30 m × 30 m相连的标准样地,将每个样地划分为9个10 m × 10 m样方。
在调查中发现,该地区群落分层明显,为避免植物生长状况引起统计偏差,因此参照袁泉等(2021)对于植物生长型界定,以群落中木本植物为对象,计树高(H)≥5 m且胸径(DBH)≥5 cm记录为乔木层,树高(H)<5 m且胸径<5 cm的木本植物为灌木层,为避免统计遗漏,部分个体树高(H)≥5 m但胸径<5 cm作为亚乔木种计入乔木层,树高<5 m但胸径≥5 cm的植株则计入灌木层。在统计物种种类、株数、胸径、基径、高度的同时,记录每种地形的海拔、坡向、土壤类型、坡位等信息(表1)。

表 1 样地基本信息Table 1 Basic information of sample plot
1.3 物种多度表示方法
为避免模型在进行数学处理和解释时会造成困难而难以比较不同的物种分布,因此本研究采用累积经验分布函数 (empirical cumulative distribution function, ECDF) (陈俊等,2018),使用Wilcoxon秩和检验对比不同地形的群落之间的物种多度差异。
1.4 主要种组成
采用物种重要值(importance value,IV)衡量群落中物种的重要程度(张田田等,2020)。以4种地形群落样地物种数据为基础,计算不同地形下乔木层与灌木层树种的重要值。重要值=(相对多度+相对频度+相对显著度)/3。其中,相对多度=某一物种多度/所有物种多度和,相对频度=某一物种出现频率/所有物种出现频率,乔木层物种的相对显著度=某一物种胸高断总面积/所有物种胸高断总面积,灌木层物种的相对显著度=某一物种基部总面积/所有物种基部总面积。
1.5 多度模型拟合及检验
1.5.1 中性模型 (1)复合群落零和多项式模型(meta community zero-sum multinomial,mzsm)零和多项式模型存在两点假设:一是假设在群落中所有生物的个体数总体没有变动,群落中不同物种多度变化是一个零和过程(Luís et al., 2012),如果一个物种多度的增加,会导致另一物种多度等量减少,双方得失相抵,总数为零;二是群落中个体物种的生存以及物种形成速率一致。该模型是中性模型的代表:

(1)

(2)

(3)
式中:为样地内的个体数,是基本生物多样性指数。
(2)Volkov模型(Volkov model,Volkov)
Volkov模型是基于零和多项式模型提出的,该模型相较于后者多了1个迁移系数(Volkov et al.,2003),Volkov模型公式如下:
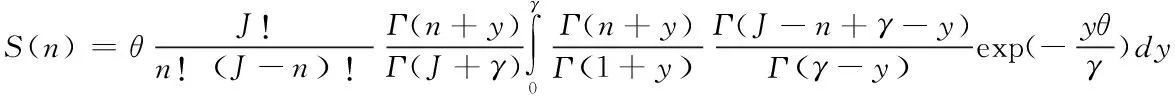
(4)
(5)

(6)
式中:为迁徙的总个体数;为迁移系数。
1.5.2 生态位模型 (1)生态位优先模型或几何级数模型(geometric series model,gs)
该模型假设最先进入的物种优先占有群落资源总和的份,而第2个进入的物种又占有余下总资源的份,即(1-)。后面的物种依次占据先前物种进入后余下总资源的份,直到资源殆尽,群落达到饱和无法再容纳下一物种截止。为第个种的多度(Whittaker, 1965)。公式如下:
(1-)-1(=1,2,3…,)
(7)
(2)断棍模型(broken-stick model,bs)
该模型假设某一环境的群落所有生态位为长为1的小棍,将这一小棍分为段,代表着总体生态位被个物种占据,模型假定个物种拥有相等的系统分类地位与竞争力,且在群落中同时出现。表示群落内物种的个体总数,表示第物种的多度(Robert, 1957)。公式如下:

(8)
1.5.3 纯统计模型
(1)对数级数模型(log-series model,ls)(马克平等,1997),公式如下:

(9)
式中:()表示多为的物种数;代表群落内物种多样性。
(2)对数正态分布模型(log normal model,lnorm)(Preston, 1948),公式如下:
=log()+log()(=1,2,3,…,)
(10)
式中:为正态分布的均值;代表方差;为第个物种的多度。
1.6 拟合效果检验
采取Kolmogorov-Smirnov检验(K-S)来检验模型拟合接受情况,通过比较各自检验的显著性概率();采用赤池信息量准则(Akaike Information Criterion, AIC)来验证拟合优度,其公式为:=-2ln()+2,通过该公式计算出的值来比较物种多度的拟合程度,AIC兼具简约性与最优性。本研究的数据处理与分析在软件Excel 2010中完成,拟合与检验与绘图均在R-3.6.3软件中进行,其中模型拟合、绘图与AIC检验在“sads”包中完成,K-S检验在“Matching”包完成(Anchi, et al., 2019)。
2 结果与分析
2.1 群落物种组成
在4种地形群落样地内共调查到乔木1 442株,根据计算重要值可知(表2),在乔木层存在黄梨木()、化香树()、光皮梾木()、齿叶黄皮()、多脉青冈()等主要物种;调查到灌木9 005株,灌木层有九里香()、南天竹(a)、川桂()、小叶柿()、黄花恋岩花()等主要物种;4种地形的群落个体数与物种组成见表3,由表3可知,鞍部群落的个体数最高(3 483株)。其次是槽谷(2 864株)、山脊(1 626株),个体数最低的是洼地(2 474株),不同地形物种数也存在差异,在乔木层,物种数量顺序为洼地>槽谷>鞍部>山脊,在灌木层,物种数顺序为洼地>鞍部>槽谷>山脊,两个群落层次在物种数量呈相似的格局,且灌木层的物种数在不同地形下均高于乔木层的物种数,其中洼地群落的个体数虽然最少,但是其物种最为丰富(乔木65种,灌木140种),而山脊地形的群落个体数与物种数均较低。

表 2 群落主要物种组成Table 2 Main species composition of community

表 3 不同地形群落的物种组成Table 3 Species composition of communities under different terrains
2.2 不同地形下群落物种多度分布格局
茂兰喀斯特森林群落4种喀斯特典型地形下群落物种的累计经验分布如图2所示。由图2可知,不同地形下群落两种生长型物种的ECDF均呈现一定程度的S型,所有地形下群落均有较高的稀有种比例,且不同地形下群落的稀有种比例相近,而灌木层的比例比乔木层更高。根据Wilcoxon秩和检验得知,不同地形下群落的乔木层ECDF之间并无显著性差异(>0.05),4种地形下群落的灌木层的物种多度存在显著差异,其中山脊与鞍部(=0.049)、洼部(=0.004 8)差异显著,鞍部与槽谷(=0.001 26)、洼部(=0.000 1)也存在显著不同。

A. 乔木层; B. 灌木层。A. Tree layer; B. Shrub layer.图 2 不同地形群落物种的累计经验分布函数Fig. 2 ECDF of species in different terrain communities
2.3 不同地形下群落物种多度模型拟合
2.3.1 不同地形植物群落乔木层物种多度模型拟合 4种喀斯特典型地形下森林群落乔木层的物种多度分布格局模型拟合效果见表4。经K-S检验与AIC检验发现,山脊的拟合优度顺序为中性模型>纯统计模型>生态位模型,其中对复合群落零和多项式模型接受最好(AIC=242.1),对断棍模型拟合也较好(AIC=260.9),而对于几何级数模型接受最差(AIC=1 862.6),从图3中也可直观看出:山脊地形的物种多度等级与几种模型多度等级的拟合效果,表明山脊地形下的物种多度构建机制不仅受中性过程控制,还受一定程度的生态位过程影响;槽谷对于中性模型与纯统计模型接受较好,且两种模型拟合效果差异很小,而对于生态位模型接受较差,断棍模型的拟合值为319.1,拒绝了几何级数模型,说明该地形下乔木层的物种多度构建主要以中性过程为主,断棍模型代表的生态位过程也发挥作用,几何级数模型代表的过程未明晰;鞍部对不同模型拟合效果与山脊相似,同时受生态位过程与中性过程控制,拟合优度的顺序为中性模型>纯统计模型>生态位模型,中性过程更加强烈;而洼地仅对中性模型接受较好,拒绝了两种生态位模型和纯统计模型中的对数正态分布模型,表明该地形物种多度构建的生态位过程不明晰或被掩盖,中性过程成为主要作用机制。由上述可知, 山脊、鞍部这两种地形下乔木层的多度构建机制比较复杂,受两种生态学过程控制,槽谷、洼地则主要受中性过程控制。此外,不同地形下的群落有不同的基础多样性指数与迁移系数,这与喀斯特地区较高的生境异质性有关。

mzsm. 复合群落零和多项式模型; Volkov. Volkov模型; bs. 断棍模型; gs. 几何级数模型; ls. 对数级数模型; lnorm. 对数正态分布模型; 观测值表示物种多度。下同。mzsm. Metacommunity zero-sum multinomial distribution model; Volkov. Volkov model; bs. Broken-stick model; gs. Geometric series model; ls. Log-series model; lnorm. Lognormal model. Observed value represents species abundance. The same below.图 3 不同地形下乔木层物种多度分布及模型拟合Fig. 3 Species abundance distribution and model fitting of tree layer under different terrains

表 4 不同地形植物群落乔木层物种多度的模型拟合Table 4 Model fitting of species abundance in tree layer of different terrains
2.3.2 不同地形植物群落灌木层物种多度模型拟合 4种喀斯特典型地形下森林群落灌木层的物种多度分布格局模型拟合效果见表5,经K-S检验与AIC检验发现,山脊对于中性模型与纯统计模型接受较好且拟合优度接近,而拒绝了两种生态位模型;槽谷与鞍部对于3种理论模型的拟合接受情况与山脊相似,对于中性模型与纯统计模型的接受优于生态位模型;洼地对中性模型与纯统计模型的接受较好,拒绝生态位理论的几何级数模型,仅接受断棍模型的拟合(=0.574),可知洼地灌木层物种多度的构建主要受中性过程主导,断棍模型代表的生态学过程也具有一定作用(图4)。由上述可知,4种地形下植物群落灌木层的物种多度对几种理论模型的接受拟合情况与乔木层相似,中性模型、纯统计模型的适用性优于生态位模型,表明4种地形下植物群落灌木层的物种多度构建也主要以中性过程为主,灌木层物种多度对于生态位模型接受情况比乔木层更差,表明生态位过程在4种地形下灌木层物种多度构建的作用更弱或更难被揭示。而对比不同地形下两种生长型植物的物种多度拟合值发现,乔木层对于模型拟合值要优于灌木层,灌木层物种多度模型拟合值随地形的不同波动也更大。不同地形下植物群落的灌木层也有更高的基础多样性指数。

图 4 不同地形下灌木层物种多度分布及模型拟合Fig. 4 Species abundance distribution and model fitting of shrub layer under different terrains

表 5 不同地形植物群落灌木层物种多度的模型拟合Table 5 Model fitting of species abundance in shrub layer of different terrains
3 讨论与结论
3.1 群落物种组成
群落主要物种组成是决定群落性质的基本要素,而重要值是衡量群落中物种优势度的常用指标(郑俊鸣等,2021)。调查发现,乔木层存在黄梨木、化香树、光皮梾木、齿叶黄皮、多脉青冈等主要物种,在灌木层存在九里香、南天竹、川桂、小叶柿、黄花恋岩花等主要物种,群落的垂直层次存在差异。该地区整体坡度与小生境凹凸度较大,导致群落存在间隙为灌木物种提供生长空间,因此该地区森林群落不同生长型的物种组成差异明显;不同地形下群落有着不同的物种数量与个体数,在喀斯特地区土壤条件比常态地貌严峻,因此水热条件群落维持群落个体数量的重要条件(Chen et al., 2009),洼地、槽谷属于低洼地势,坡度较小,有利于水热能源的积累,因此环境能支持更多个体存活,山脊水土流失性大,因此引起地形之间的群落个体数量差异。群落物种最丰富的是洼地,乔木层与灌木层的物种数均高于其他地形,该地形地势平坦,空间变异性小,为物种进入与共存创造了稳定环境(王艳红等,2020),槽谷与鞍部这两种地形不仅开阔,其结构也适合水分保持,因此两种地形物种数也较高,山脊由于不利于水土养分积累而限制了物种的丰富度与种群扩大。
3.2 不同地形下群落物种多度分布
累计经验函数分布函数与Wilcoxon秩和检验可以表现并验证群落的物种多度分布差异,解释稀有种、一般种和常见种的组成特征(Thomas et al., 2015)。本研究结果表明,4种喀斯特典型地形下群落的乔木层与灌木层均拥有较高稀有种比例,与白浩楠等(2016)、方晓峰等(2021)对同纬度带常态地貌下群落的物种多度研究结果相似,而该地区的典型喀斯特地形下的森林群落有更高稀有种比例。茂兰喀斯特森林群落位于亚热带季风气候区,受东南季风影响,雨热充沛,为植物生长提供养分,而该地区空间异质性较高,限制了优势种的扩散,为稀有种进入创造了机会,一些种群在竞争中也逐渐成为稀有种,因此该地区存在着较高比例的稀有种。此外,稀有种相较于常见种在群落物种多度中具有显著贡献,刘旻霞等(2020)在对甘南亚高寒草甸稀有种的研究时已验证;经Wilcoxon秩和检验发现,4种地形下群落乔木层的物种多度分布不存在显著差异,可能研究对象均处于同一地区,该地区虽然土壤贫瘠稀少,但是充足的雨热、光照资源是该地区鲜明的特征,植物对于资源的需求极大地提升空间和资源利用率,因此乔木层的物种多度格局虽然受地形异质性影响,但是经过长期适应有了更加明显的地区资源格局特征。而灌木层之间的物种多度随地形差异出现显著不同,灌木层的物种进入环境的初期,并处于生长阶段,因此对地形异质性带来的资源差异的响应更加剧烈。
3.3 不同地形下群落物种多度模型拟合
4种典型地形下群落的物种多度格局能很好地接受中性模型与纯统计模型,且不同地形对模型的拟合优度存在差异,而对于生态位模型接受较差。虽然纯统计模型通过数学与生态学结合的方法拟合群落物种多度格局,对于量化生态学过程、表征群落多样性有作用,但是该模型对物种多度构建机制解释不足。本研究的纯统计模型对物种多度拟合结果较优,与康佳鹏等(2021)、郭跃东(2021)等研究结果相似,丰富了该模型的适用范围;本研究也验证了中性理论在该地区森林群落的物种多度格局构建过程中的重要作用,该地区的环境相异性较高,加之溶岩形成微生境较多,所以扩散限制与随机过程发挥作用。各群落的拟合值也随地形不同发生变化,表明中性理论的作用强度受生境异质性影响,其中山脊的拟合最优,可能由于该地形生境严酷导致中性过程更强烈。此外,乔木层比灌木层能更好拟合中性模型,乔木层的物种经历中性过程作用达到稳定,因此其物种多度格局有更深的中性过程印记。而灌木层的物种处于进入、生长时期,扩散限制的作用未显现,导致中性过程未被模型量化出来,加之灌木层的物种较丰富,物种的生态性状重叠也影响了模型拟合;4种典型地形群落的物种多度格局对于生态位模型接受较差,是由于该地区群落属于顶极演替阶段,物种已达到稳定共存,生态位过程已弱化,尤其是洼地、槽谷两种地形的地势平坦,资源相对丰富。但是,不同地形下群落的灌木层多数拒绝了生态位模型,仅洼地接受断棍模型,是因为洼地部位地势平坦,环境相对稳定,利于物种总体生态位的分割,而其他地形的环境异质性较高,物种多度构建受到随机因素影响较深,因此生态位过程表现不明显或被掩盖。
虽然本研究利用不同模型解释了喀斯特地区典型地形下群落的物种多度分布格局,但是作用过程有待进一步揭示(Laroche et al.,2020),并且用物种多度分布格局推测其后形成的生态学机制也存在一定的不确定性,因为不同的生态学过程共同作用可能会导致相同或相似的物种多度格局(Etienne, 2010)。此外,两种理论都不具有良好的兼容性,因此仅用模型拟合去探究群落物种多度分布格局形成具有局限性,目前研究学者更倾向于探索两种理论的整合,量化不同群落组合机制的相对重要性,完善并丰富理论(Liu et al., 2013)。当然,群落物种多度格局的构建是一个伴随着演替且复杂的生态学进程,受影响的因素极多,除了关注理论的创新,还需要综合考虑环境因素,取样尺度和研究方法、土壤因子以及功能性状等因素也会影响研究结果。
