黔西南北盘江镇喀斯特高原峡谷区植被演替阶段碳氮稳定同位素特征
2022-07-28吴银菇喻阳华李一彤
吴银菇, 喻阳华, 李一彤, 郑 维
( 1. 贵州师范大学 地理与环境科学学院, 贵阳 550025; 2. 贵州师范大学 喀斯特研究院/国家喀斯特石漠化防治工程技术研究中心, 贵阳 550001 )
碳(C)、氮(N)作为必需生命元素,其生物地球化学循环过程一直都是生态学研究的重点与热点(熊鑫等,2016)。植物-土壤δC、δN组成能够准确记录影响植物新陈代谢和生态系统C、N循环的综合作用(姚凡云等,2012;刘建锋等,2018),揭示植物对特定环境资源利用策略的响应与适应机制。稳定C、N同位素技术可示踪、整合和指示目标物,具有检测迅速、结果准确、不受时间限制等优点(郑秋红和王兵,2009),是研究森林演替过程中C、N循环的理想方法。
近年来,δC、δN值被国内外学者广泛用来指示植物水分利用效率和生态系统C、N循环机理(Zheng et al., 2018; 刁浩宇等,2019)。司高月等(2017)和Twohey等(2018)研究发现叶片δC值可表征植物水分利用率和固C耗水成本,其组成能够揭示C固定的综合特征;姚凡云等(2012)研究认为植物-土壤δN值可作为生态系统N饱和程度和N循环长期变化趋势的指示因子。显然,稳定同位素技术的发展为研究生态系统化学元素循环开启了窗口。叶片-凋落物-土壤作为生态系统C、N循环的主要载体(张萍等,2018),其同位素特征能精准刻画与森林植物群落演替相关的影响因素和环境信息变化,亦能厘清生态系统结构和功能的变化过程,因而研究三者之间C、N及稳定同位素基本特征与内在关联,有助于深入阐明喀斯特高原峡谷区森林生态系统C、N循环特征和养分分布随演替进程的变化格局。目前,针对喀斯特森林演替过程,主要集中在生态化学计量特征、土壤理化属性和土壤微生物等角度(盛茂银等,2015;皮发剑等,2016;吴求生等,2019),但基于C、N稳定同位素方法对喀斯特森林演替过程及规律研究的报道鲜见。对喀斯特生态系统演替过程中δC、δN变化特征的了解尚不全面,特别是森林演替过程中叶片-凋落物-土壤连续体的δC、δN值的分布特征及其如何指示生态学效应等,均需深入研究。
基于此,本文以贵州喀斯特高原峡谷区不同演替阶段植物群落为对象,采用稳定C、N同位素方法,通过测定4个演替阶段植物叶片-凋落物-土壤C、N及稳定同位素,分析其互作效应,试图回答以下2个科学问题:(1)探讨喀斯特森林不同演替阶段植物叶片-凋落物-土壤C、N及稳定同位素特征,阐明森林δC、δN值随演替发生的变化规律;(2)探究随森林演替过程,植物叶片-凋落物-土壤C、N与其稳定同位素间的内在关联,揭示喀斯特森林演替过程中植物资源利用与适应策略的变化规律,为诊断养分状况和制定喀斯特森林生态系统可持续经营措施提供理论支撑。
1 材料与方法
1.1 研究区概况
研究区位于贵州省黔西南州贞丰县北盘江镇(105°38′11″ E、25°40′16″ N),属典型的喀斯特高原峡谷地貌,地表破碎,地势起伏大,海拔为370~1 473 m。区内属亚热带季风气候,年均降水量为1 100 mm,但时间分布不均,集中在5—10月,季节性干旱严重。年总积温为6 542.9 ℃,年均温为18.4 ℃,年均最高、最低温度依次为32.4、6.6 ℃,四季干湿、冷热明显,光热条件丰富。土壤类型以石灰土为主,土层浅薄且肥力较低,基岩裸露率高。植被以亚热带常绿落叶针阔混交林为主,受人为活动干扰,原生植被基本被破坏,次生植被和人工林分布居多(杜家颖等,2017)。突出的生态问题包括水土流失严重、生态系统脆弱等。
草灌阶段以毛葡萄()、叶下珠()、多花木蓝()等为主;灌木阶段以川钓樟()为优势种,滇鼠刺()、清香木()等均有分布;乔灌阶段以朴树()、毛八角枫()、山麻杆()居多;乔木阶段以圆叶乌桕()、翅荚香槐()为主,南酸枣()、构树()等亦有分布。
1.2 群落调查
2019年7—8月,在对研究区进行全面踏查的基础上,参考卢永飞等(2015)对喀斯特植被演替阶段的划分方法,将植物群落划分为草灌、灌木、乔灌和乔木4个演替阶段(表1)。每个演替阶段设置3个20 m × 20 m的调查样地(海拔810 ~ 850 m,经度、纬度和坡度差异均较小,坡向一致),进行植物群落学调查。受喀斯特地表破碎、土层浅薄等特殊地形地貌的影响,采样时将样地间距确定为>10 m。灌木和草本调查样方大小依次为10 m × 10 m、1 m × 1 m,在每个样地内设置4个灌木小样方和4个草本小样方,树高<2 m的木本植物即记为灌木。同时,记录乔本名称、树高、胸径、冠幅;灌木名称、树高、地径、盖度;草本名称、盖度、平均高度。调查的环境因子包括经纬度、海拔、坡度、坡向等。

表 1 样地基本概况Table 1 Basic situation of plots

不同字母表示演替阶段之间存在显著性差异(P<0.05)。下同。Different letters indicate significant differences among the succession stages(P<0.05). The same below.图 1 不同演替阶段植物群落叶片C、N含量及稳定同位素值Fig. 1 Contents and stable isotope ratios of C and N in leaves of plant communities at different succession stages
1.3 样品采集方法
在对样地进行调查分析的基础上,将重要值大于0.2的树种视为优势种,每个样地随机选取5株优势种,摘取东、南、西、北、中5个方位无病虫害、健康成熟的叶片,混匀制成1个样品装入尼龙袋。每个样地按S形路线采集未分解、半分解及全分解层凋落物(全分解层凋落物以不能辨识种类但未形成腐殖质层为判断依据),组成1个混合样装入尼龙袋。同时在采集凋落物的地方,采集0~20 cm土壤,组成混合土样,采用四分法保留鲜土约0.5 kg。4个演替阶段植物叶片、凋落物和土壤样品各12份(4个演替阶段 × 3个标准样地)。
1.4 样品处理与分析
样品带回实验室后,叶片和凋落物在(65±2)℃温度下烘干至恒重后,粉碎、过筛备用;土壤用镊子剔除根系、凋落物、石块和动植物残体,置于室内自然风干后,用研磨机粉碎并依次通过2.00、0.15 mm筛,装入专用玻璃瓶中密闭保存,用于测定C、N含量及稳定同位素自然丰度。
叶片-凋落物-土壤C、N含量及δC和δN值在自然资源部第三海洋研究所实验室采用元素分析仪-稳定性同位素质谱仪联机(Vario ISOPOTE Cube-Isoprime, Elementar公司)完成测定,同位素比值采用千分比单位(‰),用δ表示。
δC值以PDB国际标准作为参考标准,计算公式如下:
(‰)=[()/()-1] ×1000。式中,()为国际标准物PDB(Peedee Belemnite)的C同位素比值,值的分析精度为±0.2‰。
δN值以空气中N作为参考标准,δN计算公式如下:
(‰)=[()/()-1] ×1000。式中,()为空气中N的N同位素比值,值分析精度为±0.25‰。
1.5 数据处理与分析
利用单因素方差分析(One-way ANOVA)检验森林不同演替阶段叶片-凋落物-土壤C、N含量和δC、δN比值之间的差异,利用最小显著差异法(least significant difference,LSD)进行多重比较,采用Pearson相关分析法对参数进行相关性检验。数据整理、分析、制图等采用Microsoft Excel 2010、SPSS 20.0、Origin Pro 2018软件完成。
2 结果与分析
2.1 不同演替阶段叶片C、N含量及δ13C、δ15N值特征
不同演替阶段优势种叶片C以灌木阶段为最小(402.55 g·kg),说明其有机物含量较低,暗示潜在的C积蓄相对较弱。叶片δC值以乔灌阶段为最低(-31.31‰),且与草灌、灌木和乔木阶段间差异显著(<0.05,下同),随演替进程先减小后增加,表明植物水分利用效率随演替进行发生了部分调整。叶片N含量、δN值依次为11.97~29.35g·kg、-3.41‰~1.52‰,均以乔灌阶段为最低,随演替进行无明显变化规律,叶片N含量、δC和δN演替前期总体上大于演替后期(图1)。
凋落物C含量以乔灌阶段为最大(475.9 g·kg),显著高于草灌、灌木和乔木阶段,表明乔灌群落有利于生态系统养分回归。凋落物N含量以乔灌阶段为最小(10.55 g·kg),草灌和乔木、灌木和乔木阶段之间均无显著差异(>0.05,下同)。凋落物δC值为-29.96‰~-20.07‰,以草灌阶段为最高,随演替进行呈降低趋势,说明凋落物演替初期更易于分解。凋落物δN值为-2.61‰~0.99‰,随演替发生表现出正、负值交替出现的现象,表明该值的变异率较大(图2)。
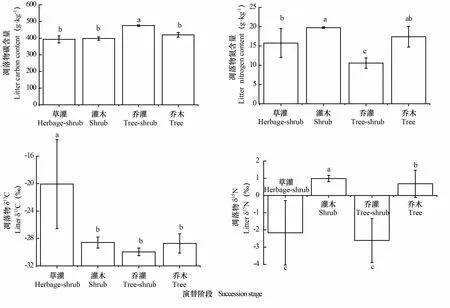
图 2 不同演替阶段植物群落凋落物C、N含量及稳定同位素值Fig. 2 Contents and stable isotope ratios of C and N in litters of plant communities at different succession stages
2.2 不同演替阶段凋落物C、N含量及δ13C、δ15N值特征
2.3 不同演替阶段土壤C、N含量及δ13C、δ15N值特征
土壤C、N含量依次为57.3~147.65 g·kg、5.45~16.15 g·kg,均表现为灌木、乔木阶段最高,乔灌阶段其次,草灌阶段最低,说明C、N循环之间存在紧密的耦合关系。土壤δC值为-26.83‰~-21.14‰,随演替进行呈降低趋势,草灌阶段显著高于其他3个阶段,暗示其土壤有机质分解更彻底。土壤δN值为5.36‰~8.63‰,随演替进行先减小后增大,总体上,土壤更表现出N富集,表明随森林演替,土壤矿化过程N分馏增强(图3)。

图 3 不同演替阶段植物群落土壤C、N含量及稳定同位素值Fig. 3 Contents and stable isotope ratios of C and N in soil of plant communities at different succession stages
2.4 叶片-凋落物-土壤C、N含量及δ13C、δ15N值之间的相关关系
由表2可见,叶片-土壤C、N及其同位素之间存在较强的相关性,表明植物-土壤之间养分循环存在紧密关联,具有显著协同或权衡效应。凋落物N、δC和δN与其他指标均无显著相关关系,凋落物C与土壤N、δN分别呈显著、极显著负相关(-0.777、-0.845),说明凋落物与土壤之间的互作关系强于凋落物与叶片。
3 讨论与结论
3.1 叶片-凋落物-土壤连续体δ13C、δ15N值特征
本文研究区森林4个演替阶段叶片δC值为-31.31‰~-28.23‰,较鼎湖山非喀斯特森林偏正(表3),原因可能是:其一,该研究区属于喀斯特高原峡谷,而鼎湖山基岩为泥盆系的厚层砂岩、砂页岩,属于非喀斯特地区,土壤类型主要为赤红壤,其保水性能较喀斯特地区好,加之鼎湖山降水量(1 955 mm)亦高于该区(1 100 mm),使土壤水分趋于丰富;其二,喀斯特地貌特殊的地上、地下二元结构,导致水分可利用性偏低,而水分可利用性与其利用效率、叶片δC值均呈负相关(Xu et al., 2017),据此推断该区较低的水分可利用性导致叶片δC值更高。叶片δN值为-3.41‰~1.54‰,与滨海沙地人工林植物叶片相比偏正(表3),表明该研究区森林生态系统N饱和程度相对较高,N含量愈加丰富,原因是喀斯特高原峡谷区生境脆弱,资源供应能力相对匮乏,植物通过提高对资源的竞争能力来适应这一特殊生境,将更多养分用于器官构建。

表 2 不同演替阶段叶片-凋落物-土壤C、N含量及δ13C、δ15N值之间的相关分析Table 2 Correlation analyses between for C and N contents and δ13C and δ15N values of leaf-litter-soil at different succession stages
本文4个演替阶段凋落物δC值为-29.96‰~-20.07‰,高于黔西北次生林优势种凋落物C值(表3),原因可能是凋落物种类、微生物数量及活性等均存在差异,加之不同样区生境水分、岩石裸露率等异质性较高,影响了凋落物的分解速率和程度。凋落物δN值为-2.61‰~0.99‰,高于罗绪强等(2014)在贵州清镇研究的结果(表3),原因是研究区采取了植树造林、退耕还林等森林生态修复与保护措施,植物群落发生正向演替,生态系统结构趋于复杂,地表凋落物组成向多样化发展,微生物数量增多,凋落物分解速率加快,有利于N富集的氨挥发增加,从而导致地表凋落物N值偏高(潘复静等,2011; 罗绪强等,2014)。
本研究区4个演替阶段土壤δC值为-26.83‰~-21.14‰,较高寒草甸土壤偏正(表3),原因是不同研究区温度具有较大差异,高寒草甸属高原大陆性气候,而该研究区属亚热带季风气候,温度明显较高,高温促进了土壤微生物活性,使土壤有机质分解速率加快(周咏春等,2019),进而导致土壤δC值升高。土壤δN值为5.36‰~8.63‰,较鼎湖山森林土壤偏高(表3),这与微生物分解凋落物补偿土壤养分过程有关,Collins等(2008)研究认为微生物C限制条件下,土壤矿化过程N分馏加强,本文结果显示凋落物C与土壤δN值极显著负相关,与该结论一致,原因是喀斯特高原峡谷区森林凋落物蓄积量虽逐渐增加,但总体偏少,分解补充到土壤中的C量仍较低,导致其C含量低于非喀斯特地区,微生物主要受C限制,矿化过程N分馏增强,土壤δN值增大。

表 3 贵州喀斯特森林与其他研究区域叶片-凋落物-土壤δ13C、δ15N值比较Table 3 δ13C and δ15N values in leaf-litter-soil in Guizhou karst forest communities in comparison with forests of other regions
3.2 叶片-凋落物-土壤δ13C、δ15N值与其元素相关性随群落演替的关系
不同层次间C含量变异较为显著,原因可能是:其一,随着演替进展,植物群落结构、林内小气候以及凋落物的输入、输出和分解状况等均发生改变,导致归还土壤的养分含量不同(熊鑫等,2016);其二,生态系统 C、N循环之间有着密切关联,固C潜力较大程度受限于土壤供N能力(李德军等,2018),受群落结构、环境因子和供N能力等诸多因素协同作用,森林C含量变异尤为明显。此外,本研究还发现叶片-凋落物-土壤δC、δN随演替进程变化规律较弱,说明森林演替过程发生了同位素分馏,原因是随演替发生,地表植被类型、水热条件以及生态系统养分分配格局等均存在显著差异,植物为适应环境、维持生存,亦采取了相应的资源利用和适应策略。伴随着资源利用策略和生存方式的改变,植物光合作用、呼吸作用也会同步变化,进而影响森林稳定C、N同位素分馏机制,最终导致各组分δC、δN值随演替过程的变化规律各异。但植物新陈代谢如何驱动稳定C、N同位素分馏尚需深入研究。
本文研究结果还显示,乔灌阶段植物群落叶片δC值最低,原因是乔、灌木混交林中,上层乔木枝叶茂盛,其叶片阻挡了进入林中的部分光线,导致林内光照强度和温度降低(刘效东等,2014),致使光合能力下降,林内大气CO浓度上升,导致升高,δC值降低。物种水平上,叶片δC值可表征植物长期的水分利用效率(Yu et al., 2008),指示固C耗水成本,亦能评价植物对逆境的适应能力(Hussain et al., 2018)。通常,叶片δC值越大,水分利用效率越高,固C耗水成本越低(Yu et al., 2008),固定相同数量C所消耗的水分越少。本研究发现,该研究区内川钓樟、圆叶乌桕和翅荚香槐等优势种叶片δC值较高,表明该区灌木和乔木阶段的树种具有较高的水分利用效率,光合作用固C耗水成本较低, 适应水分胁迫生境的能力更强,可作为生态系统恢复的优选树种。

