“蜀脆枣”果实发育过程中糖酸代谢相关基因表达分析
2022-07-22邓群仙张慧芬张小艾李红春
王 羊,邓 倩,2,邓群仙,张慧芬,张小艾,李红春
(1.四川农业大学 园艺学院,成都 611130;2. 四川省广安市邻水县经果技术推广所,四川广安 638500;3. 四川省眉山市彭山区农业农村局,四川眉山 620800)
枣是原产中国的优势特色果树[1]。中国枣产业分为北方产区和南方产区[2],南方产区以栽植酸甜适口的鲜食枣为主[3]。
糖酸是果实最重要的内在品质,其组分、含量及比例共同决定果实风味[4]。桃、梨等成熟果实以蔗糖为主要糖组分[5-6],蔗糖磷酸合酶基因(SPS)、转化酶基因(INV)、蔗糖合酶基因(SS)等共同影响蔗糖含量[7]。苹果以积累果糖为主[8],葡萄以积累葡萄糖为主[9],己糖激酶基因(HK)、果糖激酶基因(FK)、尿苷二磷酸葡萄糖焦磷酸化酶基因(Zj UGP)等协同调控果糖和葡萄糖含量[10]。李主要积累苹果酸,磷酸烯醇式丙酮酸羧化酶(PEPC)、苹果酸酶(NADP+-ME)、NAD+-苹果酸脱氢酶(NAD+-MDH)是影响苹果酸含量的关键因子[11]。柑橘为柠檬酸积累型,柠檬酸合酶基因(CS)、乌头酸合酶基因(Aco)、苹果酸酶基因(ME)等基因共同影响柠檬酸含量[12-13]。
‘骏枣’‘冬枣’‘赞皇大枣’属蔗糖积累型,‘山东梨枣’和‘北京鸡蛋枣’属果糖积累型[14-15];‘冬枣’和‘新郑灰枣’等大多数枣的苹果酸含量最高,‘稷山板枣’奎宁酸含量最高,而‘太古鸡心蜜’琥珀酸含量最高[14,16]。不同品种的枣果实糖酸积累模式不同。目前北方枣的糖酸代谢分子机理研究较为深入,而南方鲜食枣的相关报道较少。
“蜀脆枣”为‘鲁北冬枣’引种四川后发现的早果丰产优质芽变材料,定植第3年进入丰产期,无需环剥亩产量达2 000 kg以上,肉质细脆,酸甜爽口,鲜食品质极佳[17]。课题组前期已开展“蜀脆枣”果实发育规律及基本营养品质的相关研究,但果实糖酸积累规律及相关代谢的机理尚不清楚。本试验以“蜀脆枣”为试材,测定果实发育过程中可溶性糖和有机酸组分与含量,分析糖酸代谢相关基因的表达,探索“蜀脆枣”果实糖酸形成与调控机制,以期为南方鲜食枣的分子辅助育种和品质调控奠定基础。
1 材料与方法
1.1 试验地概况
试验园位于四川省德阳市罗江区白马关镇万佛村五组(31°26′34″N,104°46′18″E),海拔约660 m,气候属于亚热带湿润型,气候温和,四季分明,最高气温36.6 ℃,最低气温-6.7 ℃,年均气温16.5 ℃;年均降水量910 mm,年无霜期278 d,年均日照时数1 260 h[17]。
1.2 供试材料
试验树为露地栽培3 a生嫁接苗“蜀脆枣”,株行距为2 m×3 m,单株为一小区,3次重复。谢花20 d后开始采集大小一致、无病虫害的果实,前期每隔14 d采样一次,中后期每隔7 d采样一次[17]。花后20~34 d为幼果期,花后48~69 d为膨大期,花后76 d为白熟期,花后83 d为着色初期,花后90 d为成熟期(半红期)。果实置冰盒中带回实验室,切碎,液氮处理后立即放入-80 ℃超低温冰箱中保存。
1.3 测定指标及方法
1.3.1 果实糖酸组分及含量的测定 糖组分及含量的测定:用液氮将果实研磨成粉末,准确称取0.5 g,加入5 mL乙醇-水(80∶20,V∶V),80 ℃水浴30 min,常温6 000 r/min离心10 min,上清液移至25 mL容量瓶,重复提取1次,定容后经0.22 μm滤膜过滤待测[18]。色谱柱Innoval NH2(250 mm×4.6 mm, 5 μm),流动相乙腈∶水 (80∶20,V∶V),示差检测器,流速1 mL/min,检测池温度40 ℃,柱温35 ℃,进样量10 μL[19]。
酸组分及含量的测定:用液氮将果实研磨成粉末,准确称取0.5 g,加入3 mL 0.04 mol/L的KH2PO4,超声20 min,常温6 000 r/min离心20 min,上清液过0.22 μm滤膜过滤待测[20]。色谱柱MZ PerfectSil Target C18(250 mm×4.6 mm, 5 μm),流动相4%甲醇水溶液,流速0.5 mL/min,柱温25 ℃,检测波长210 nm,进样量10 μL[20]。
1.3.2 糖酸代谢相关基因相对表达量的分析 总RNA的提取及cDNA第一链的合成:参照改良CTAB法提取RNA,琼脂糖凝胶电泳检测RNA质量,核酸蛋白仪检测RNA浓度。cDNA第一链的合成参照TOYOBO反转录试剂盒(ReverTra Ace qPCR RT Master Mix with gDNA Remover)。
实时荧光定量PCR:根据NCBI上枣的全基因组序列,利用Primer Premier 5.0设计有机酸代谢相关基因的引物,糖代谢引物及内参基因引物序列参考张春梅[21](表1)。采用TOYOBO荧光定量试剂盒(THUNDERBIRD SYBR qPCR Mix Without ROX)在定量PCR仪(Bio-Rad,USA)上进行扩增反应。UBQ为内参基因,反应总体积10 μL,包含:THUNDERBIRD SYBR qPCR Mix Without ROX 5 μL,10 μmol/L 上下游引物各0.4 μL,cDNA模板1 μL和无RNA酶水3.2 μL。PCR扩增程序:95 ℃ 30 s,95 ℃ 5 s,Tm 30 s,72 ℃ 30 s,40个循环。基因表达水平用相对表达量2-ΔΔCt表示。以上所有试验均设3次重复,每次试验设阴性对照。

表1 引物序列Table 1 Primers used in this study
1.4 数据处理与分析
采用Excel 2016软件进行数据处理,采用SPSS 22.0系统进行统计分析。
2 结果与分析
2.1 “蜀脆枣”果实发育过程中的糖酸积累
2.1.1 可溶性糖组分及含量的动态分析 “蜀脆枣”发育过程中总糖含量大致呈逐渐上升趋势,成熟期达到最大值146.83 mg/g(图1)。果糖和葡萄糖含量相当,为14.66.47~27.48 mg/g,发育过程中变化平稳,约占发育前中期总糖的80%。白熟期蔗糖含量迅速上升,并在成熟期上升至最大值107.43 mg/g,约占总糖的73%。

误差线为标准误,不同小写字母表示同一组分不同花后时间在0.05水平上差异显著。下同
2.1.2 有机酸组分及含量的动态分析 “蜀脆枣”总酸含量随果实发育整体呈现“W”型的变化趋势,花后48 d达到峰值6.67 mg/g(图2)。“蜀脆枣”的有机酸组分主要有3种,其含量和变化趋势均不一致。苹果酸含量最高,呈上升-下降-上升趋势,成熟期达3.30 mg/g,占总酸的 65.74%。柠檬酸含量次之,呈“W”型趋势,成熟期含量为1.56 mg/g,占总酸的31.08%。酒石酸发育过程中变化不大,且含量最低,成熟期含量为0.16 mg/g,仅占总酸的3.19%。
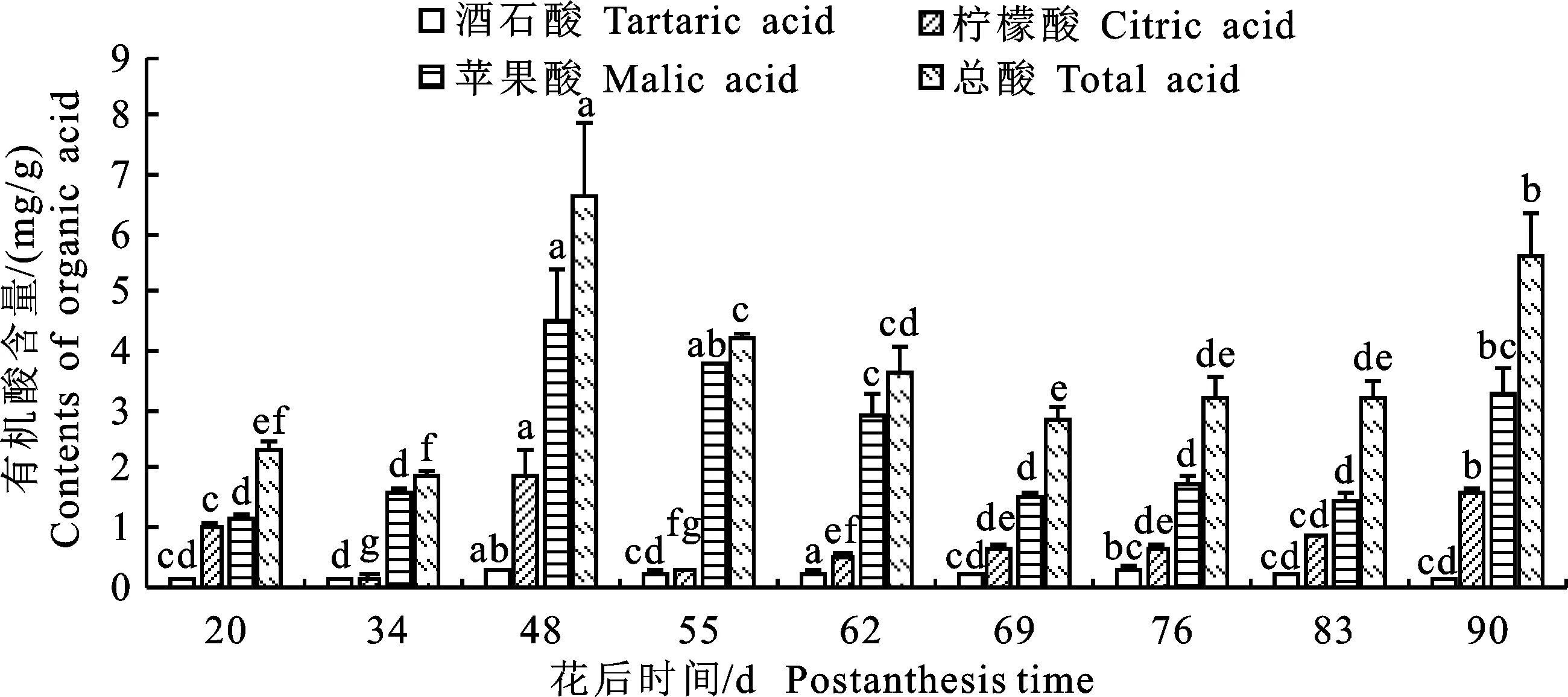
图2 “蜀脆枣”果实发育过程中有机酸组分及含量Fig.2 Components and contents of tartaric acid, citric acid, and malic acid during development of “Shucuizao” fruits
2.2 糖代谢相关基因的表达分析
2.2.1 蔗糖代谢途径中相关基因的表达分析 “蜀脆枣”果实发育过程中蔗糖代谢途径中相关基因的表达模式各不相同(图3)。Zj SS1和Zj SS2在花后20 d的表达最高,随果实发育整体逐渐下降;Zj SS3的表达在成熟期达峰值。Zj SPS1、Zj SPS2和Zj SPS4的表达整体呈先上升后下降的趋势,均于花后83 d达到最大值。Zj nINV1、Zj vINV2和Zj cINV3在幼果期和膨大期有高表达,Zj vINV2在白熟期以后几乎不表达。
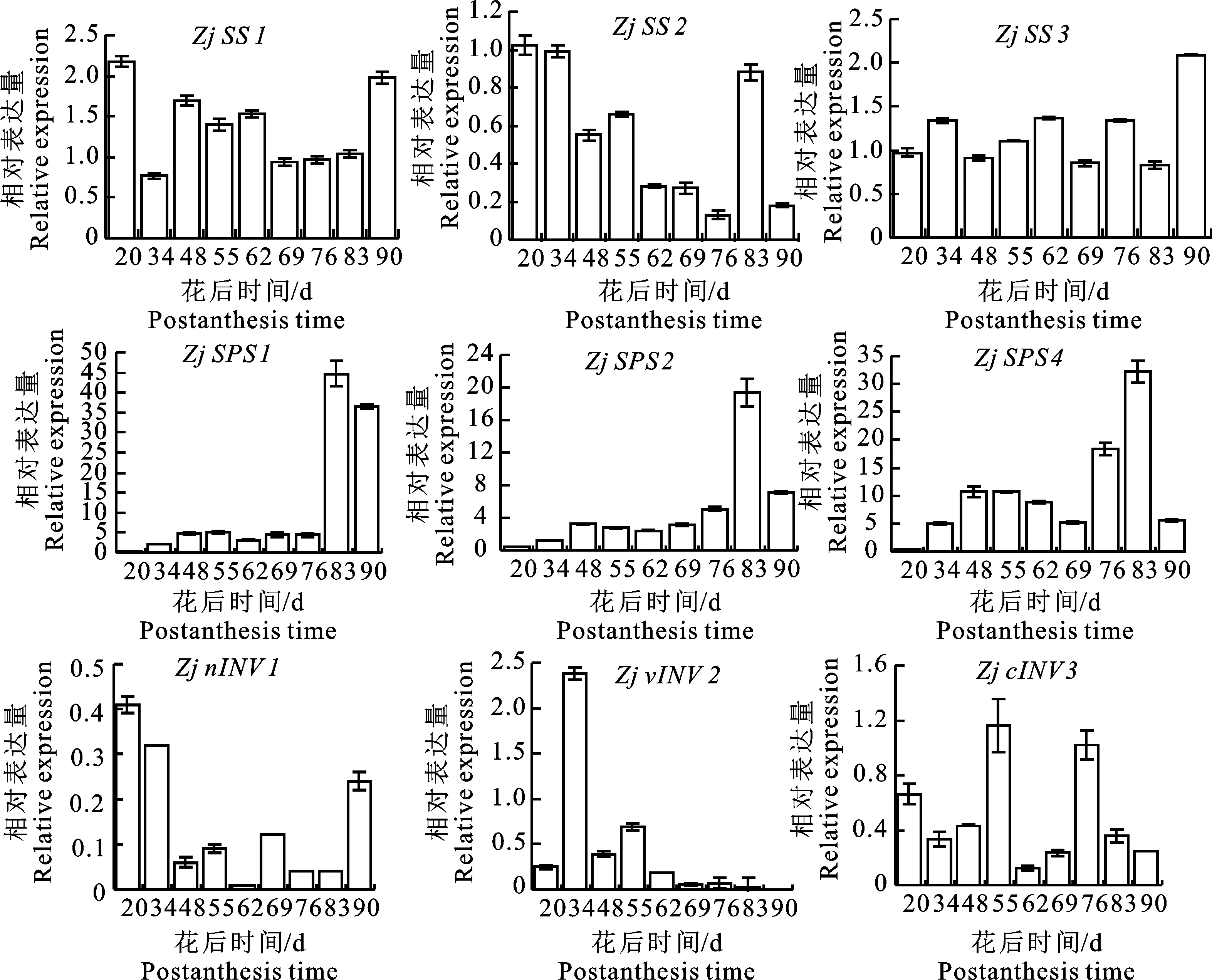
图3 “蜀脆枣”果实发育过程中蔗糖代谢途径中相关基因的表达分析Fig.3 Analysis of relative genes expression in sucrose metabolism during development of “Shucuizao” fruits
2.2.2 己糖代谢途径中相关基因的表达分析 己糖代谢相关基因在“蜀脆枣”果实不同发育时期存在差异表达(图4)。Zj HK1、Zj HK5的表达在幼果期最高,随果实发育下调,成熟期的表达最低。Zj HK3、Zj HK6、Zj UGP、Zj FK2的表达整体呈上升-下降-上升趋势。Zj FK4的表达随果实成熟先上升后略微下降。
2.3 果实发育过程中有机酸代谢相关基因的表达分析
2.3.1 苹果酸代谢途径中相关基因的表达分析 “蜀脆枣”苹果酸代谢相关基因在不同发育时期有不同的表达模式(图5)。Zj NADP-ME1的表达量在成熟期最低;Zj NADP-ME4的表达呈现“M”型趋势。MS在果实发育前中期表达较低,于成熟期急剧上调。Zj MDH1、Zj MDH4的表达在成熟期最高。Zj FUM的表达量在花后69 d达峰值。Zj PEPC1在幼果期和膨大期的表达量较高,随后下调,在成熟期略微上调;Zj PEPC4的表达量在花后20 d最低。Zj PEPCK1-1在果实发育中期几乎不表达,成熟期的表达量最高。
2.3.2 柠檬酸代谢途径中相关基因的表达分析 “蜀脆枣”果实发育过程中柠檬酸代谢相关基因有不同的表达模式(图6)。Zj ATP-CS2和Zj ATP-CS3的表达整体呈先上升后下降趋势,均在花后34 d表达量最高。Zj CS3、Zj NAD-IDH1-2、Zj NADP-IDH2、Zj NAD-IDH1-1、Zj Aco2-1随果实成熟大致上升-下降-上升,在幼果期的表达量均最低。

图6 “蜀脆枣”果实发育过程中柠檬酸代谢途径中相关基因的表达分析Fig.6 Analysis of relative genes expression in critic acid metabolism during development of “Shucuizao” fruits
2.4 果实糖酸含量与代谢相关基因的相关性分析
2.4.1 糖含量与其代谢基因的相关性 表2是“蜀脆枣”17个糖代谢基因表达量与可溶性糖含量的相关系数。果糖含量与Zj nINV1、Zj cINV3、Zj HK1呈极显著或显著负相关,与Zj SPS4、Zj HK2、Zj HK6、Zj UGP呈显著或极显著正相关。Zj SS1、Zj nINV1、Zj cINV3与葡萄糖含量呈显著或极显著负相关,而与Zj HK2、Zj HK6、Zj UGP呈显著或极显著正相关。蔗糖含量与Zj SS2、Zj vINV2、Zj HK1、Zj HK2、Zj HK5、Zj FK4表达量呈显著或极显著负相关,与Zj SS3、Zj SPS1、Zj SPS2极显著正相关。
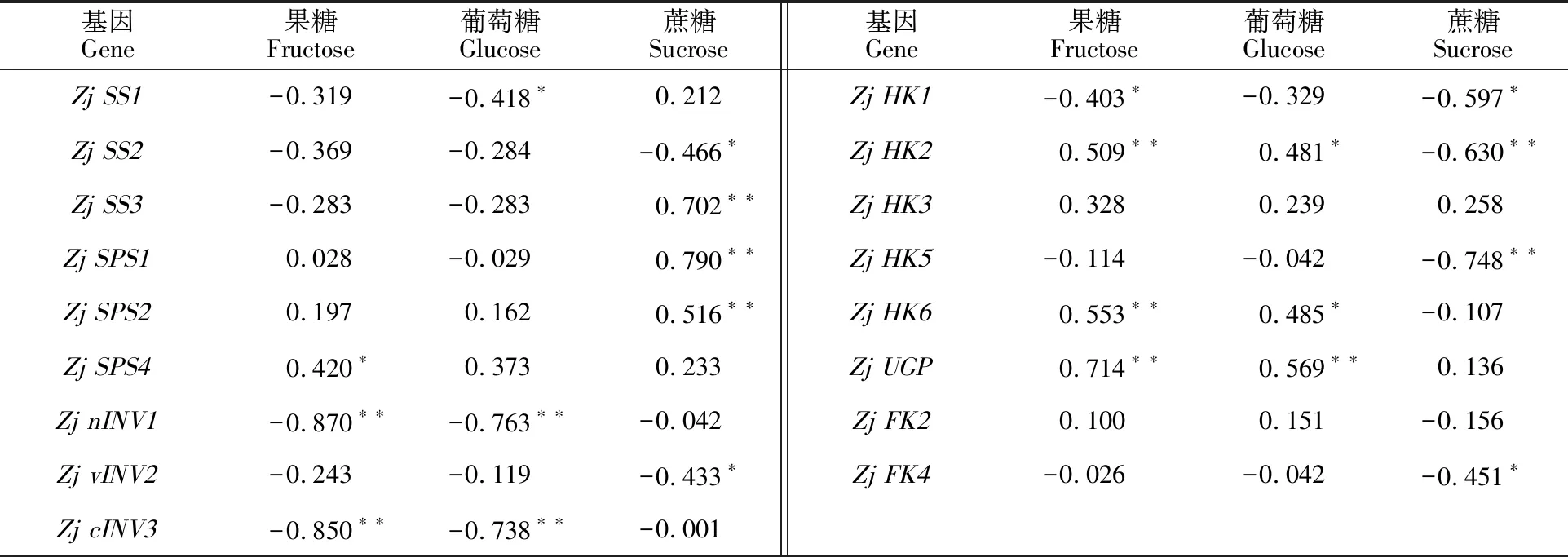
表2 “蜀脆枣”果实发育过程中糖含量与其代谢基因的相关系数Table 2 Correlation coefficient between sugar content and its metabolism gene during development of “Shucuizao” fruits
2.4.2 有机酸含量与其代谢基因的相关性 表3为“蜀脆枣”17个有机酸代谢基因表达量与有机酸含量的相关系数。酒石酸含量与Zj NADP-ME1、Zj PEPCK1-1、Zj ATP-CS2表达量呈显著负相关,而与Zj PEPC1呈显著正相关。柠檬酸含量与Zj NADP-ME1、Zj NADP-ME4、Zj ATP-CS2、Zj ATP-CS3表达量呈极显著或显著负相关,而与Zj MS、Zj PEPCK1-1、Zj CS3、Zj NAD-IDH1-2呈显著正相关。苹果酸含量与Zj PEPC1、Zj PEPC4、Zj NAD-IDH1-1、Zj Aco2-1、Zj NADP-IDH1表达量呈极显著或显著正相关。
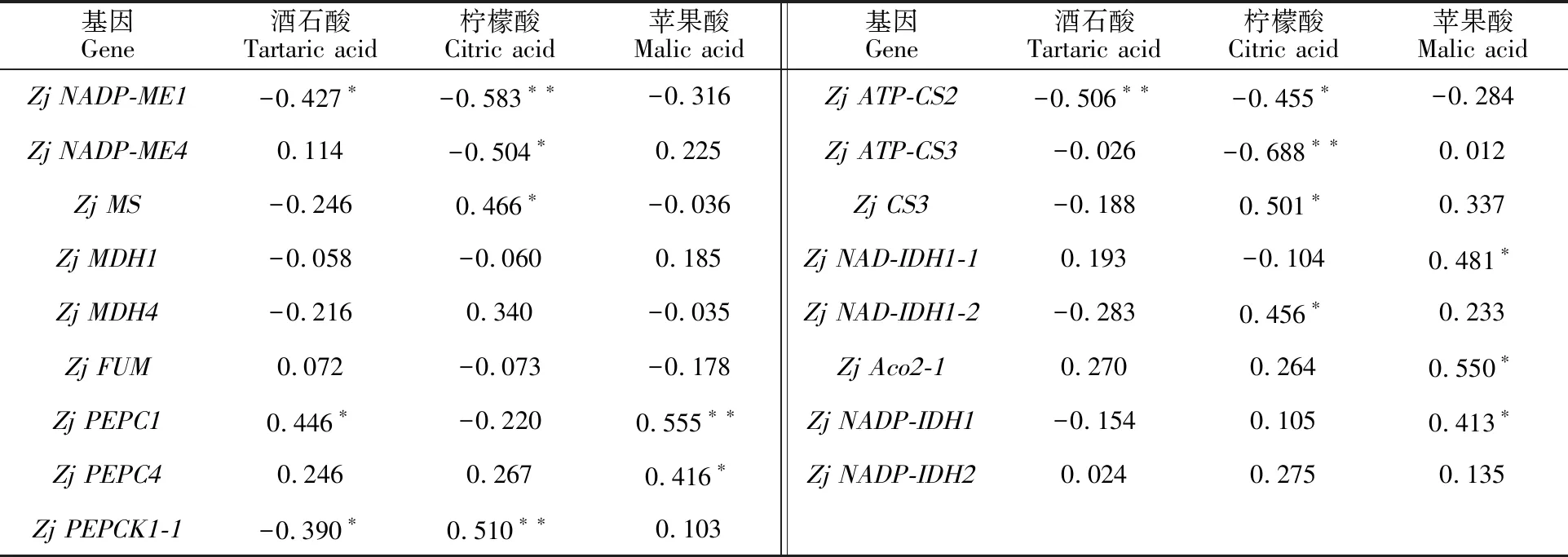
表3 “蜀脆枣”果实发育过程中酸含量与其代谢基因的相关系数Table 3 Correlation coefficient between organic acid content and its metabolism gene during development of “Shucuizao” fruits
3 讨 论
在本研究中,“蜀脆枣”幼果期和膨大期主要积累果糖和葡萄糖,成熟期主要积累蔗糖,与‘金铃圆枣’成熟期主要积累己糖不一致[22]。“蜀脆枣”有机酸主要由苹果酸、柠檬酸和酒石酸组成,苹果酸含量始终最高,与‘梨枣’等一致[23]。“蜀脆枣”属蔗糖、苹果酸积累型的优质鲜食枣。
糖代谢由多基因综合调控,SS调控果糖合成蔗糖,葡萄糖在HK、UGP和SPS作用下合成蔗糖,INV调控蔗糖降解成果糖和葡萄糖[24-25]。本试验中,Zj nINV1、Zj cINV3与果糖和葡萄糖含量呈显著负相关,果实发育中后期INV的表达下调致使果糖和葡萄糖合成减少;ZjHK2、Zj HK6和Zj UGP与果糖和葡萄糖含量呈显著或极显著正相关,果实发育后期其表达量较低或变化不大,致使果糖和葡萄糖降解减少,两方面结合导致果糖和葡萄糖含量变化不大。MdHK1的上调表达显著促进了富士苹果葡萄糖转化[26],与本研究不一致,推测是不同种类的果实糖代谢相关基因差异表达的结果。“蜀脆枣”白熟期以后蔗糖迅速积累,Zj SS3、Zj SPS1和Zj SPS2迅速上调,且与蔗糖含量极显著正相关;Zj vINV2几乎不表达,与蔗糖呈显著负相关。果实发育后期Zj SS3、Zj SPS1和Zj SPS2的高表达与Zj vINV2的低表达协同促进蔗糖的快速积累。本研究结果与蒋爽等[27]研究结果一致,但与Zj SS3表达促进‘骏枣’‘灰枣’和‘冬枣’果实还原糖积累的研究结果[15]相反,推测是不同品种间的糖代谢差异所致。蔗糖的迅速积累是“蜀脆枣”糖代谢的基础,Zj SS3、Zj SPS1、Zj SPS2和Zj vINV2是糖代谢关键基因。
PEPC催化磷酸烯醇式丙酮酸生成草酰乙酸,草酰乙酸经MDH催化生成苹果酸,草酰乙酸与乙酰辅酶A经CS催化形成柠檬酸,运输至液泡,柠檬酸又被Aco可逆催化形成异柠檬酸,NADP-IDH促进异柠檬酸合成α-酮戊二酸,NAD-IDH则催化异柠檬酸生成α-酮戊二酸[28]。CS的高表达促进西瓜[29]、碰柑[30]和酸枣[21]果实中的柠檬酸积累。本试验中,成熟期Zj PEPCK1-1、ZjMS、Zj CS3、Zj NAD-IDH1-2的高表达共同促进果实柠檬酸含量上升。Zj PEPCK1-1与柠檬酸含量相关性最强,是调控“蜀脆枣”柠檬酸代谢的关键基因。花后48~55 d和花后90 d 的Zj PEPC1、Zj PEPC4、Zj NAD-IDH1-1、Zj Aco2-1、Zj NADP-IDH1高表达协同促进了苹果酸高水平积累。Zj PEPC1与苹果酸含量相关性最强,是影响苹果酸含量变化的主要因素。但马倩倩[31]认为Zj PEPC对酸枣中苹果酸的代谢调控作用不大,推测果实种类的不同是造成这种差异的主要因素。苹果酸含量是“蜀脆枣”总酸积累的主要因素,Zj PEPC1对“蜀脆枣”有机酸代谢起主导作用,Zj PEPC4、Zj NAD-IDH1-1、Zj Aco2-1、Zj NADP-IDH1起辅助作用。
4 结 论
“蜀脆枣”发育前期以积累果糖和葡萄糖为主,随后蔗糖迅速积累,成为成熟期的主要糖组分;苹果酸含量在发育过程中始终最高,其次是柠檬酸,酒石酸含量最低。“蜀脆枣”果实发育过程中糖酸代谢相关基因表达模式存在差异,相关基因的表达和调控共同影响果实的糖酸积累。Zj SS3、Zj SPS1、Zj SPS2和Zj vINV2是调控果实糖代谢的关键基因,Zj PEPC1是影响有机酸代谢的主要因素。
