转 GhB301基因棉花抗枯萎病机理研究
2022-07-22朱金成刘戈辉张鹏飞郭文婷
朱金成,刘戈辉,张鹏飞,郭文婷,张 薇
(石河子大学 农学院/新疆生产建设兵团绿洲生态农业重点实验室,新疆石河子 832003)
棉花是重要的经济作物,中国是世界上最大的棉花生产和消费国。棉花枯萎病是棉花的重要病害之一,由于常年连作,病害逐年加重,严重影响棉花的产量和品质,制约着棉花产业的可持续发展,培育抗枯萎病的棉花新品种是解决这个问题的最经济有效的方法。分子生物学的迅速发展,为利用转基因技术培育棉花抗病品种提供了新的途径[1]。
AP2/ERF转录因子是一种具有特殊结构、行使调控基因表达功能的蛋白质分子[2]。Guo等[3]从海岛棉‘7124’克隆得到一个ERF类转录因子GbERF1-like,可以通过激活木质素合成提高对黄萎病菌的抗性。Liu等[4]发现在烟草中过表达GbERFb可以提高烟草对大丽轮枝菌的抵抗力。在拟南芥中,过量表达B3亚组的AtERF14能增强防卫基因的表达,并能调控ERF1、ERF2等其他抗病相关ERF基因的表达。进一步的研究发现erf14突变体对枯萎病菌更加感病,且AtERF14的功能缺失不能被其他ERF互补;表明AtERF14在枯萎病抗性中表现出很重要的作用[5]。过量表达GbERF1可以提高棉花对大丽轮枝菌的抗性,而下调GbERF1基因则增加棉花对大丽轮枝菌的敏感性[6]。此外,ERF(亚)家族转录因子在信号转导途径中也起到了重要作用。ERF(亚)家族转录因子的基因受多种信号分子的诱导而表达[7]。Guo等[8]研究发现,棉花等多种高等植物受到黄萎病等真菌侵染时会发生应激反应而产生许多小分子物质,如乙烯和水杨酸等。Zhang等[9]研究发现,小麦中的ERF(亚)家族转录因子TaERF3在对病原物白粉菌的抗性反应途径中发挥作用,从而激活植物对病原物的防卫相关基因的表达。
综上所述,AP2/ERF转录因子作为植物特有的一类转录因子,在植物对病原菌胁迫响应中具有重要的调控作用。近年来,虽然人们已克隆了一些与抗枯萎病相关的ERFs家族基因,但仍有许多调控棉花抗枯萎病反应的关键基因的功能尚不清楚。刘戈辉等[10]从高抗枯萎病的棉花品种中克隆了1个ERF转录因子基因GhB301,通过农杆菌介导法将其导入野生型棉花品种YZ-1并获得纯合株系,枯萎病抗性鉴定结果表明,过表达GhB301的转基因棉花株系病情指数为 14.77%,显著低于野生型对照(病情指数为 37.50%),显著提高棉花对枯萎病的抗性。本研究在此基础上,通过分析GhB301基因对棉花抗病相关基因的表达及酶活性的影响,探讨GhB301基因在棉花抗枯萎菌中的功能,为棉花抗枯萎病分子育种提供理论依据和基因资源。
1 材料与方法
1.1 棉花材料的种植
供试棉花材料为过表达GhB301转基因棉花纯合株系(OE)与野生型对照YZ-1(WT),其中棉花转基因纯合株系在石河子大学绿洲生态农业重点实验室前期试验中获得。将转基因株系和野生型对照材料的种子分别用50~60 ℃的温水浸泡30 min,放在发芽盒中于37 ℃培养箱中暗培养 3 d。然后将发芽的种子清理好种植于营养钵中[V(花土)∶V(蛭石)=3∶1)],放置于光照培养箱(光照16 h、25 ℃,黑暗8 h、23 ℃,湿度65%)中进行培养,待棉苗第1片真叶完全展平时进行接菌处理。
1.2 枯萎病菌的活化及接菌处理
供试菌株为枯萎病菌7号生理小种强致病菌株F430,由石河子大学农学院棉花分子育种实验室提供。活化后挑取单菌落于查氏液体培养基中,28 ℃、200 r/min暗培养7~10 d,待菌液达到1×107个孢子/mL时对营养钵中棉苗进行伤根处理,用注射器吸取10 mL菌液注射到伤口处,对照注射10 mL的去离子水。每次处理30株,重复3次,用于后续指标测定。
1.3 抗病相关酶活性的测定
分别取转基因棉花材料和野生型材料接菌后0、1、3、5、7 d的叶片,液氮速冻后,保存于-80 ℃冰箱。过氧化物酶(POD)、多酚氧化酶(PPO)、苯丙氨酸解氨酶(PAL)的活性测定参考汪红等[11]的方法,超氧化物歧化酶(SOD)、过氧化氢酶(CAT)活性测定参考Bailly等[12-13]的方法。
1.4 抗病相关基因的表达分析
提取枯萎病菌处理后24 h的转基因株系和对照系的根部RNA,反转录成cDNA后,进行qRT-PCR检测。内参基因为GhUBQ7,qRT-PCR反应体系及程序设计参考TransStart Tip Green qPCR SuperMix说明书。共设置3个重复,采用2-ΔΔCt法计算基因相对表达量[14]。用primer 5软件设计引物(表1),引物由北京华大基因公司合成。
1.5 数据分析
使用Excel 2010、SPSS 22软件对数据进行处理分析。
2 结果与分析
2.1 棉花叶片防御酶活性分析
2.1.1 SOD活性变化 在接种棉花枯萎病菌后,无论是转基因材料(OE)还是野生型材料(WT)中超氧化物歧化酶(SOD)活性都比相应的接水对照(CK-OE、CK-WT)高,并且都随着接菌时间的延长而出现先升高后降低的变化趋势,但其SOD活性大小与峰值出现时间不同(图1)。其中转基因材料的峰值出现较早,在接菌后的第3天,是同期转基因接水的2.66倍,野生型材料峰值出现的较晚,在接菌后的第5天,是同期野生型接水的1.78倍。
2.1.2 POD活性变化 由图2可知,两个接水处理的棉花材料叶片中过氧化物酶(POD)活性均呈现先升高后降低的趋势,并且转基因材料(OE)的POD活性均高于野生型材料(WT),尤其是在第3天,是野生型接水对照(CK-WT)的2.20倍。在接菌处理后,转基因材料和野生型材料POD活性均呈现先升高后降低的变化趋势,且转基因材料的POD活性均高于相应的野生型材料,转基因材料在第3天达到峰值,而野生型材料在第5天才达到峰值;在接种后3 d时,转基因接菌材料是转基因接水材料的2.26倍,野生型接菌材料是野生型接水材料的1.56倍。
2.1.3 CAT活性变化 由图3可以看出,在接菌处理条件下,转基因材料(OE)叶片CAT活性与野生型(WT)相比显著增加;随着接菌时间的延长,两个棉花材料叶片CAT活性均呈现先升高后降低的趋势,转基因材料叶片峰值出现在接菌后的第3天,是同期转基因接水对照(CK-OE)的2.57倍,野生型材料叶片CAT活性在第5天出现峰值,是同期野生型接水对照(CK-WT)的1.41倍。在接水处理的条件下,转基因材料叶片的过氧化氢酶(CAT)活性与野生型材料无明显差异;随着处理时间的延长,野生型棉花叶片CAT活性呈现先升高后降低的趋势,并且在第5天达到峰值,转基因材料叶片中CAT活性变化不明显。
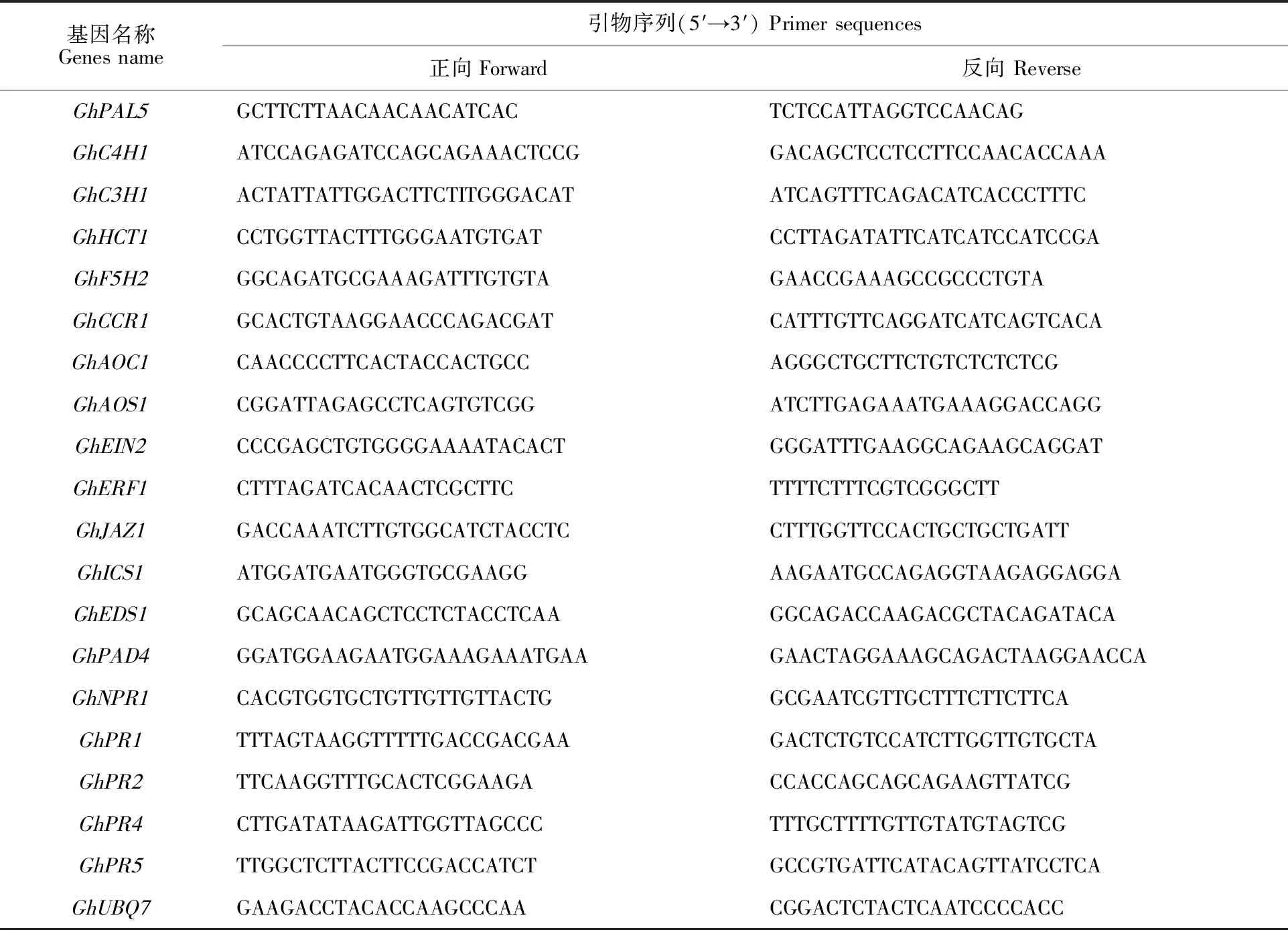
表1 qRT-PCR所用引物Table 1 Primers used for qRT-PCR
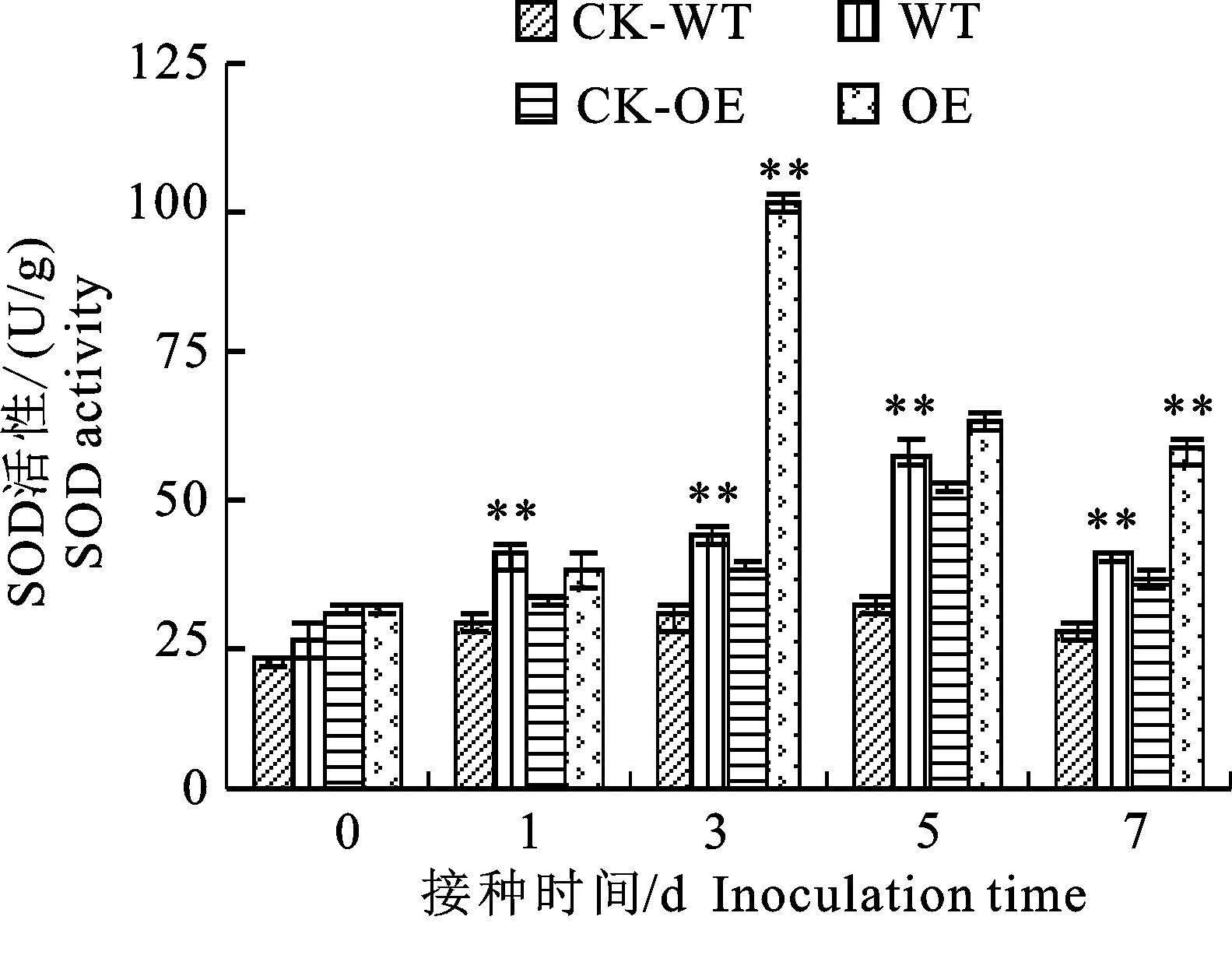
*和**分别表示同期接菌处理与接水对照间在0.05和 0.01水平存在显著性差异;下同

图2 转基因材料与野生型材料接种枯萎病菌后POD活性变化Fig.2 Changes of peroxidase activity in transgenic and wild-type cotton plants after inoculation with Fusarium oxysporum
2.1.4 PAL活性变化 如图4所示,在接菌处理条件下,两个材料PAL活性均出现先升高后降低的变化趋势,与POD、SOD活性不同的是,转基因材料(OE)在第5天达到峰值,是同期转基因接水材料(CK-OE)的3.04倍,但在野生型材料(WT)中PAL活性在第3天达到峰值,是同期野生型接水对照(CK-WT)的1.75倍。在接水处理条件下,两个材料也都表现出先升高后降低的趋势,但不明显。

图3 转基因材料与野生型材料接种枯萎病菌后CAT活性变化Fig.3 Change of catalase activity in transgenicand wild-type cotton plants after inoculation with Fusarium oxysporum

图4 转基因材料与野生型材料接种枯萎病菌后PAL活性变化Fig.4 Change of phenylalanine ammonia lyase activity in transgenic and wild-type cotton plants after inoculation with Fusarium oxysporum
2.1.5 PPO活性变化 图5显示,在接种棉花枯萎病菌后,转基因材料(OE)和野生型材料(WT)中多酚氧化酶(PPO)活性都比相应的接水对照(CK-WT、CK-OE)显著增加,并且都随着接菌时间的延长而出现先升高后降低的变化趋势,两个材料的PPO活性峰值均出现在第3天,但其PPO活性大小不同。在第3天转基因接菌材料是同期转基因接水材料的2.98倍,野生型接菌材料是同期野生型接水材料的2.02倍。在接水处理的条件下,两个材料的PPO活性变化也呈现先升高后降低的趋势,峰值均出现在第5天,并且在第5天时转基因接水材料的PPO活性显著高于野生型接水材料,是野生型接水材料的 1.22倍。
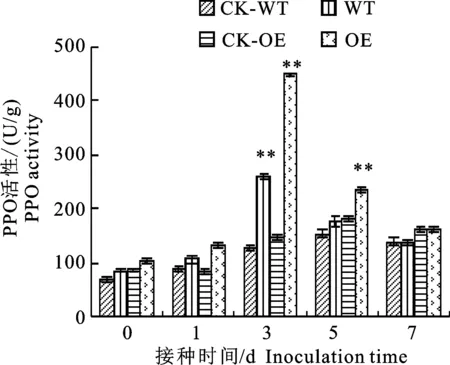
图5 转基因材料与野生型材料接种枯萎病菌后PPO活性变化Fig.5 Change of polyphenol oxidase activity in transgenic and wild-type cotton plants after inoculation with Fusarium oxysporum
2.2 抗病相关基因的表达分析
为了进一步阐明GhB301在棉花枯萎病抗性中的功能,利用qRT-PCR方法测定转基因棉花材料(OE)和野生型材料(WT)在接种枯萎病菌24 h后苯丙烷代谢途径相关基因、JA/ET路径相关基因、SA路径相关基因、PR基因等抗病相关基因的表达,如图6所示。结果表明,与野生型材料相比,转基因材料在枯萎病菌处理后除GhERF1、GhJAZ1、GhNPR1外,大多数基因均上调表达。
在苯丙烷代谢途径中(图6-A),共检测6个基因,其中GhPAL5、GhC4H1、GhC3H1、GhF5H2在转基因材料中的相对表达量均极显著高于野生型材料,GhCCR1在转基因材料中的相对表达量显著高于野生型材料,GhHCT1在转基因材料和野生型材料中无明显差异。
在JA/ET路径中(图6-B),共检测5个基因,其中乙烯合成的两个关键基因GhAOS1在转基因材料中的相对表达量极显著高于野生型材料,GhAOC1在转基因材料中的相对表达量显著高于野生型材料;GhEIN2在转基因材料和野生型材料中无明显差异,仅略高于野生型材料,而GhERF1和GhJAZ1在转基因材料中的相对表达量均低于野生型材料,特别是GhJAZ1在转基因株系中的相对表达量显著低于野生型材料。
在SA路径中(图6-C),共检测4个基因,其中GhICS1和GhEDS1在转基因材料中的相对表达量极显著高于野生型材料,GhPAD4在转基因材料中的相对表达量显著高于野生型材料,GhNPR1在转基因材料和野生型材料中无明显差异。
在PR基因中(图6-D),共检测4个基因,其中只有GhPR4在转基因材料中的相对表达量显著高于野生型材料,另外3个基因在转基因材料中仅略高于野生型材料。
综上所述,在接种枯萎病菌24 h后,过表达GhB301棉花中与苯丙烷代谢途径、JA/ET路径、SA路径、PR基因等多个抗病相关基因被激活,仅有个别基因的表达无明显差异,由此推测过表达GhB301的转基因棉花增强对枯萎病抗性的原因可能与抗病相关基因的激活有关。
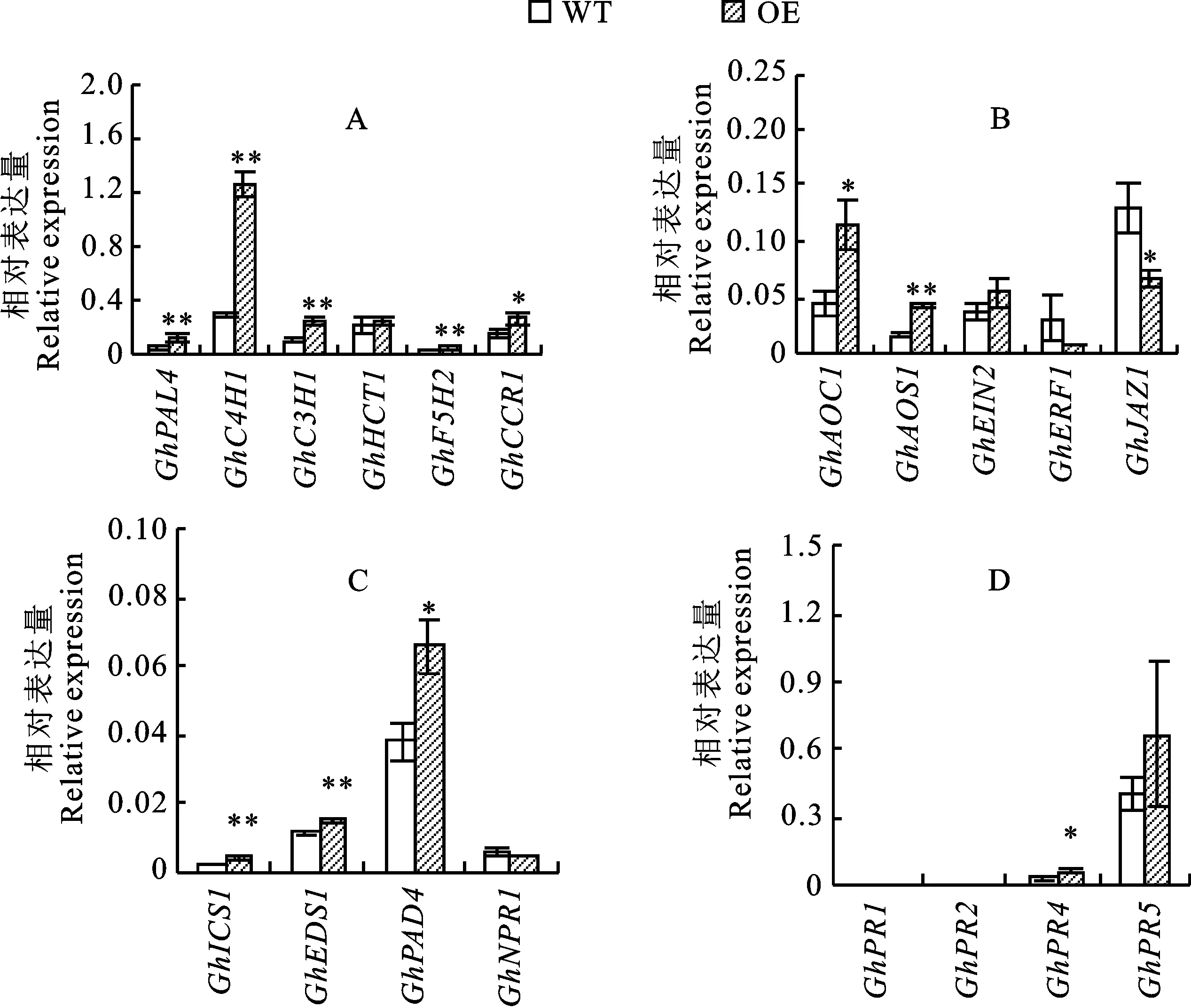
A.苯丙烷代谢途径相关基因相对表达量;B.JA/ET路径相关基因相对表达量;C.SA路径相关基因相对表达量;D.PR基因相对表达量
3 讨 论
当植株受到病原菌侵害时体内会发生一系列的生理生化变化,包括组织细胞结构形态的改变,以及相应的物理化学的抗病反映[15-16]。研究发现接种大丽轮枝菌后不同抗性棉花品种的PAL、POD、PPO、CAT等的酶活性随着品种抗病性的增强而升高[17]。Gayoso等[18]研究发现,携带有Ve基因的抗黄萎病番茄在受到黄萎病菌侵染后,PAL和POD的活性升高并且木质素含量增加。SOD、GPX、CAT和APX可以清除细胞中的活性氧,以保护细胞免受过氧化伤害,并且转基因植株的酶活性更高[19]。李娟[20]对转基因抗病棉花品种与对照材料进行接种枯萎病菌处理,发现其PAL、POD活性的变化趋势基本一致,呈现先增加后降低的趋势,但转基因抗病棉花的酶活性变化更快,峰值更高。本研究以过表达GhB301转基因棉花纯合株系和野生型对照为试验材料,接种棉花枯萎病菌后,发现棉花叶片SOD、CAT、PAL、PPO、POD活性逐渐增高,均呈先增加后降低的变化趋势,转基因棉花材料的SOD、CAT、POD、PAL、PPO活性均高于野生型材料,且SOD、POD、CAT活性峰值均早于野生型材料2 d左右,这与前人研究结果基本一致。
前人研究发现棉花代谢途径相关基因表达量变化与植物抗病性相关。苯丙氨酸/酪氨酸代谢途径是苯丙烷途径的重要分支,通过此途径可以合成木质素[21]。植物木质化程度越高,相应的抗病性就越强[22]。木质素合成相关基因可以通过正向调节木质素合成和提高木质素含量,来提高棉花对枯萎病的抗性[23]。JA/ET和SA现已被证明是植物与病原菌互作中重要的激素,植物受病原菌胁迫后能够显著改变这些激素的生物合成和下游信号相关基因的表达[24-25]。ERF1作为JA/ET响应应答调控因子起作用,超表达ERF1的拟南芥对枯萎病菌及其他一些腐生型真菌的抗性增强,ERF2也具有类似的功能[26]。研究发现GbERF1的表达受黄萎病的强烈诱导,过量表达GbERF1的转基因拟南芥能增强多个病程相关蛋白基因的表达,与野生型拟南芥相比,对黄萎病的抗性也明显提高[27]。李超[28]发现棉花和拟南芥通过增强 JAZ1 来抑制茉莉酸通路激活赤霉素通路,从而降低棉花抗病性来调节植物正常的生长发育。本研究利用实时荧光定量PCR技术测定接种枯萎病菌后转基因棉花株系和野生型株系中抗病相关基因的表达,被检测基因中大部分在转基因株系中的相对表达量显著高于野生型对照,推测接菌后有效提高苯丙烷代谢途径、JA/ET途径、SA途径中相关基因和PR基因的表达,且JAZ1作为一个负调控因子表达量降低也得到验证。结合前期对转基因棉花株系抗病性的鉴定,推测转GhB301棉花株系对枯萎病抗性增强可能是由于GhB301基因的过表达引起棉花抗病相关基因表达量升高进而增加了防御相关酶活性所致。
