池杉种皮特性和种子休眠原因*
2022-07-20杨治华陈雪梅陈春桦张丽苗李昌晓
杨治华 陈雪梅 陈春桦 张丽苗 董 智 谭 雪 李昌晓
(三峡库区生态环境教育部重点实验室 重庆市三峡库区植物生态与资源重点实验室 西南山地生态循环农业国家级培育基地 西南大学生命科学学院 重庆 400715)
三峡水库自成功运行以来,采取“蓄清排浑”的运行方式,水位海拔145~175 m。由于水库水位周期性涨落,水库两岸形成了两条平行的垂直落差达30 m,总面积约348.93 km2的季节性水位消落带(樊大勇等, 2015)。新形成的水库消落带具有水淹时间长、水淹深度大、水淹反季节性等特点。生长环境的巨大变化和反季节水淹的频繁干扰导致原有大部分植物因不能适应而逐渐消亡(卢志军等2010; 樊大勇等2015)。三峡库区消落带面临生物多样性降低、水土流失严重以及生态系统退化等一系列问题(揭胜麟等, 2012; 朱妮妮等, 2015)。因此,三峡水库消落带植被的恢复重建对维持三峡库区生态环境的健康和稳定具有十分重要的意义。研究表明,池杉(Taxodiumdistichumvar.imbricatum)能够耐受三峡水库蓄水导致的长时间和反季节水淹的频繁干扰,为三峡库区消落带适生木本植物(李昌晓等, 2008; 王朝英等, 2012; 李波等, 2015; 贺燕燕等, 2018; 刘明辉等, 2020)。
池杉为裸子植物杉科(Taxodiaceae)落羽杉属(Taxodium)(落羽松属)落叶阳性大乔木,是古老的孑遗植物(郑万钧等, 1978; 于永福,1994),原产于美国东南部和墨西哥湾沿岸,现在我国长江南北及珠江、淮河流域广范栽培(杨美凌, 2010)。除了具有耐水淹特性,池杉还具有耐盐碱、适应性强、生长迅速、木材性能好、观赏价值高等优点(汪企明等, 1993,1995),是库区消落带植被恢复和江河湿地造林的优良树种,因此研究其生产扩繁技术具有非常重要的生态和经济价值(李晓玲等, 2016)。种子繁殖是植物最常见的繁殖方式,但池杉种子不加处理直接播种育苗后发芽迟缓,苗木出土参差不齐, 连续时间长达 60余天,少数甚至隔年出苗,因此,耗种量大,成苗率低,生产管理困难(史忠礼等, 1984; 邹雪生,1991; Liuetal., 2009)。
种子休眠是具有正常活力的种子在适宜的光照、温度、水分和氧气等环境条件下仍不能萌发的现象(Baskin,1998; Finch-Savage, 2006)。种子的休眠特性是植物为适应气候变化、温度差异、水分条件等环境因素并保持自身繁殖发展而形成的一种生物学特性。不同标准将种子休眠划分为不同类型。Baskin 分类系统(Baskinetal., 2004)使用类型、级别和模式作为分类系统的3个层次,将种子休眠划分为5种休眠类型: 物理休眠(physical dormancy, PY)是因种皮或果皮透水、透气不良或产生机械阻力而导致的种子无法萌发; 生理休眠(physiological dormancy, PD)是因种子内部存在抑制物质而导致的种子无法萌发; 形态休眠(morphological dormancy, MD)是因种胚没有发育完全,经过一段时间使胚发育完全后种子才能萌发; 形态生理休眠(morphophysiological dormancy, MPD)是因种胚未发育完全而导致的生理休眠; 复合休眠(combinational dormancy, PY+PD)是因种皮或果皮不透水,同时种子内含有萌发抑制物,即兼具物理休眠和生理休眠。从生物学角度来讲,种子休眠是植物经长期演化所获得的一种对环境条件及其季节性变化的生物学适应(Guillaumeetal., 2017),对植物的生存、延续和种质资源的可持续利用均有积极作用,但也给造林、育种、引种和繁殖等带来困难。
Murphy等(1975)研究表明,池杉种子不存在胚休眠现象,萌发率低是由于硬且不透水的种皮物理障碍造成的。史忠礼(1980)研究认为,池杉种子胚本身具备发芽能力,只是由于种子的其他部位存在抑制物质,从而抑制了胚的萌发,且似属ABA(abscisic acid, ABA)的抑制物大部分存在种皮中。Downie等(1996)研究指出,从针叶树种子中切除的胚置于水中时萌发,这表明包围胚的种子组织(种皮、珠心残体和雌配子体)起着抑制和防止胚根出现的作用。目前,对池杉种子休眠原因的研究较为初步且不够全面,鉴于此,本试验旨在通过探索种皮特性,主要包括池杉种子结构、吸水性、内源抑制物等与种子休眠的关系,系统研究休眠原因,进一步明确休眠类型,以期为采取措施提高种子的发芽率和整齐性提供理论依据。
1 材料与方法
1.1 供试材料
池杉种子产自江苏省常州市(31°09′—32°04′N、119°08′—120°12′E),于 2019 年 12月下旬采收,置于4 ℃冰箱储存备用。内源抑制物发芽试验所用小白菜(Brassicachinensis)种子品种为快菜538,于2020年7月15日购于兴农种业,种子纯度≥95%,发芽率≥85%,水分含量≤7%。
1.2 种子形态观察及特征指标测定
观察池杉种子形状、质感、色泽等; 用四分法随机选取3×100粒池杉种子,用游标卡尺测定种子的纵径、横径、厚度(以种子腹面到背面种脊的最大距离),以平均值作为种子大小; 依据《GB2772-1999 林木种子检验规程》,采用百粒法测定种子千粒质量。
1.3 种子结构及种皮超微结构观察
取经98%浓硫酸处理15和30 min、60 ℃热水处理15 min的池杉种子各10粒,参考Ren等(1999)的研究方法将其置于FAA固定液中固定即经过乙醇系列(50%,2 h; 70%,17 h; 90%,2 h; 100%,2 h)脱水,然后用100%乙醇脱水3次(每次12 h),最后转入乙酸异戊酯置换后进行临界点干燥、喷金。在飞纳扫描电子显微镜下选择近胚端观察种子横断面及表面等结构,以未进行处理的种子为对照并进行记录拍照。
1.4 种子生活力和发芽率
1.4.1 种子活力测定 依据《GB2772-1999 林木种子检验规程》,采用氯化三苯基四氮唑(TTC)染色法测定种子生活力。参考宋廷茂(1984)的研究方法,对染色结果进行鉴定: 凡种胚全部染成红色,种胚的基部少许未染色(少于胚茎长度的1/3),胚茎少许未染色,或子叶少许未染色者,均称为有生活力的种子; 而种胚全部没有染上颜色,或仅有浅红色斑点,或胚茎呈环形未染色者,均视为无生活力的种子。
1.4.2 种子萌发试验 选取饱满池杉种子,用2%硫酸铜溶液消毒15 min,清水洗净后置于内垫双层滤纸的培养皿(φ9 cm),每皿40粒,5个重复,保持滤纸湿润,在人工气候箱25 ℃条件下恒温培养,周期性光照(12 h·d-1),光强3 000 lx,胚根超过2 mm即为萌发,每2天观察记录萌发情况,计算萌发指标。本试验用萌发时滞(germination time lag,GTL)、发芽势(germination energy,GE)、萌发率(germination percentage,GP)3个参数评价萌发效果。GTL(d)为萌发启动时间,指从萌发试验开始到第1粒种子开始萌发所用时间; GE(%)=第15天萌发种子数/供试种子数×100%; GP(%)=萌发种子数/供试种子数×100%。萌发时长设定为30天。
1.5 种皮透水性及种子各部分含水量的测定
分别取自然风干的完整种子、刻伤处理种子(刀片割破种皮)、98% H2SO4酸蚀30 min的种子各50粒,参考山金凤等(2019)的研究,测定种子吸水率。
种子吸水率(%)= (Wt―W0)/W0× 100%,式中:Wt为种子吸水t时间后3次测量的质量平均值,W0为种子吸水前起始质量。吸水完毕以时间为横坐标,吸水率为纵坐标,绘制吸水曲线。
在种皮透水性测定结束后,将饱和状态的池杉种子剥离出种皮和剩余部分(主要包括雌配子体和胚,以下简称种仁),以未处理的完整干种子为对照,将其种皮与种仁分别放入铝盒中,采用恒高温烘干法测定并计算种子各部分含水量,以了解在吸水试验中池杉种皮和种仁的吸水能力以及种皮是否阻碍种仁吸水。
各部分含水量(%)=(m1-m2)/(m1-m3)×100%,式中:m1为样品盒和盖及样品的烘前质量(g);m2为样品盒和盖及样品的烘后质量(g);m3为样品盒和盖的质量(g) 。
1.6 池杉种子内源抑制物的提取和生物测定
1.6.1 种子各部分甲醇浸提液的提取 准确称量池杉完整种子、种皮、种仁10 g,在预冷的研钵内研碎后过1 mm筛,用4~10倍体积的80%的甲醇溶液,在4 ℃下恒温密闭浸提,期间多次取出摇动使其充分浸提。24 h后过滤,然后再重复2次,混合3次所得浸提液,56 ℃减压浓缩,蒸干后用甲醇溶解瓶内干物质,定容至25 mL(即浓度为0.4 g·mL-1),分别取醇提液制成质量浓度为0.1、0.2 g·mL-1的溶液待用。
1.6.2 提取液的生物测定 将1.6.1中所述的溶液各3 mL置于内垫双层滤纸的培养皿(Φ9 cm)中,待有机溶液完全挥发后,每个培养皿中再加入3 mL蒸馏水和50 粒白菜种子。以同体积蒸馏水作为对照,每个处理3个重复。将各培养皿均匀的置于25 ℃培养箱内,用40 ℃浸泡20 min后的白菜籽进行不同提取液种类及浓度的发芽影响测定,以鉴定抑制物活性。以胚根长至白菜籽的2倍为发芽标准,48 h后统计白菜籽的发芽率,72 h后用游标卡尺测量白菜的苗高和根长。
1.7 种皮各溶剂相生物测定
按系统溶剂法(李庆梅等, 2013),将种皮甲醇浸提液分离为石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相,取各分离相制取质量浓度为0.5、1 g·mL-1的溶液待用。按1.6.2方法对各分离相进行生物活性测定,生物测定。
1.8 数据处理
所得数据采用Excel 2010软件处理,利用 SPSS 软件进行单因素方差分析,并用LSD 法检验其差异显著性,结果以平均数±标准误(Mean ± SE) 表示,用Origin 2018软件作图。
2 结果与分析
2.1 池杉种子基本形态
池杉种子外部轮廓呈不规则三角形或多边形,微扁; 表面光滑,有光泽,合点区(即发育成熟的种子与种鳞的分离点)表面粗糙; 有明显尖锐的种脊,皮厚,坚硬,具有瘤和凸缘(图1A); 种子红褐色,极少数深褐色。去除种皮的种仁呈长椭圆形,胚乳及胚呈乳白色(图1B),胚乳表面被覆淡褐色膜质珠心膜(图1C),子叶4~9片(图4D); 种子长(11.72±0.19) mm,宽(6.04±0.12) mm,厚(6.00±0.11) mm,千粒质量为(88.5±0.09)g; 种皮质量约占种子质量的71.61%。
2.2 种子生活力和发芽率
池杉种子的生活力测定结果为: 胚呈红色,具有生活力的种子占80.95%,未完全染色不具备生活力的种子占8.38%,无胚或胚已皱缩的空粒种子占10.67%。池杉种子发芽试验的萌发率为0,说明存在极显著的休眠现象。
2.3 种皮的超微结构
包被池杉种胚的组织主要包括种皮、残余珠心(珠心膜和珠心帽)和胚乳(即雌配子体)。其中,种皮的光滑表面是外表皮(图2A1),它由1层纵向伸长的薄壁细胞组成(图2A2),纹饰清晰,外表面覆盖着蜡质。外表皮在处理和储藏过程中极易脱落,图2A3为外表皮自然脱落的种皮横切面,显示外表皮下面是种皮层,池杉种皮分为外种皮、中种皮和内种皮3层。外种皮由1或2层形状较规则的薄壁细胞组成,细胞排列紧密,无明显间隙; 中种皮是种皮主要的坚硬部分,由数层排列紧密且强烈木质化的厚壁细胞组成,这些细胞的细胞壁分布大量纹孔; 内种皮是种皮的最内层,在成熟的干种子中,内种皮由来自薄壁细胞的压缩死细胞组成,这些细胞在种子发育过程中逐渐退化(Tillman-Sutelaetal., 1995a; 1995b),从种皮的横切面来看,池杉的内种皮以鳞片层的形式出现在中种皮的内表面(图2A3)。

图1 池杉种子的解剖结构Fig. 1 The anatomical structures of Taxodium distichum var. imbricatumA:种子正面; B:种子横切面; C:种仁; D:胚。ch:合点区; mi: 珠孔; mg: 雌配子体; em:种胚; nc: 珠心帽; nr: 珠心残余, 珠心膜; co: 子叶. A: Front of the seed; B: Cross section of seed; C: Kerne; D: Embryo. ch: Chalazal region; mi: Micropyle; em: Embryo; nc: Nucellar cap; nr: Nucellar remnants; co: Cotyledons.
与对照相比,热水处理过程中外表皮全部脱落,外种皮裸露出来,与外表皮细胞相比,外种皮细胞较短,表面纹饰清晰,细胞排列紧密,未出现纹孔和明显细胞缝隙(图2B1,B2); 热水处理后的种皮横切面显示,池杉种皮结构无明显改变,细胞层仍然排列紧密,与对照组无明显区别(图2B3)。经98%酸蚀15 min 后,与对照相比,种皮表面纹理被破坏,并出现许多腐蚀裂痕和腐蚀坑,有纹孔出现(图2C1、C2); 从横切面来看,种子外表皮和外种皮已被完全腐蚀,种皮结构受损,中种皮裸露并变薄(图2C3),酸蚀15 min增强了种皮透性。酸蚀30 min 后种皮结构种皮表面出现大量腐蚀裂痕和腐蚀坑,细胞结构被严重破坏(图2D1、D2); 中种皮细胞内侧细胞壁上的纹孔清晰可见,种皮结构进一步被破坏,中种皮进一步变薄,种皮透性进一步增加(图2D3)。

图2 池杉种皮的超微结构Fig. 2 The ultrastructure of seed coat of Taxodium distichum var. imbricatumA1-A2: 对照,未处理种子表面; A3: 外表皮自然脱落的种皮横切面; B1-B2: 60 ℃处理15 min后的种皮表面; B3: 60 ℃处理15 min后的种皮横切面; C1-C2: 酸蚀15 min后种皮表面; C3: 酸蚀15 min后种皮横切面; D1-D2: 酸蚀30 min后种皮表面; D3: 酸蚀30 min后种皮横切面. ep:外表皮; sa:外种皮; scl:中种皮; en: 内种皮; mg: 雌配子体; pi: 纹孔; nr: 珠心残余, 珠心膜. A1- A2: Control, untreated seed surface; A3: Cross section of seed coat in which the epidermis has fallen naturally; B1-B2: Surface of seed coat treated at 60 ℃ for 15 min; B3: Cross section of seed coat treated at 60 ℃ for 15 min; C1-C2: Seed coat surface after acid-scarified 15 min; C3: Cross section of seed coat after acid-scarified 15 min; D1-D2: Seed coat surface after acid-scarified 30 min; D3: Cross section of seed coat after acid-scarified 30 min; ep: epicuticle; sa: sarcotesta; scl: sclerotesta; en: endotesta; mg: megagametophyte; pi: pits; nr: nucellar remnants.
2.4 种皮的透水性
池杉种子的吸水曲线见图3,可知酸蚀处理、刻伤处理与完整种子吸水曲线变化趋势相似,吸水率随时间延长而增加。在开始吸水的24 h内,各处理吸水速度都很快,酸蚀处理的最快,吸水率达到46.39%; 吸水24 h后各处理种子的吸水速度均有所放缓。刻伤处理和完整种子经过96 h吸水后,吸水率趋向稳定,维持在38%左右。酸蚀处理种子经168 h吸水后,种子吸水率稳定在65%左右。完整种子和刻伤处理种子的吸水基本一致,差异不显著(P>0.05),但酸蚀处理种子吸水率显著增大,达到极显著差异(P<0.01)。由此可见,池杉种子自然吸水过程大致分为3个时期,即快速吸水期(0~24 h)、缓慢吸水期(24~96 h)、近饱和期(96~240 h)。
将完全吸水饱和的池杉种子进行解剖,分离出种皮和种仁两部分,测定含水量,以未经吸水性测定的干种子作为对照,结果见图4。结果表明,经不同处理的池杉种子种皮含水量均高于胚乳,在吸水达到完全饱和后,完整种子种皮含水量(42.58%)与刻伤处理种皮含水量(42.08%)无显著差异(P>0.05),而酸蚀处理(53.66%)的种皮含水量与完整种子差异显著(P<0.05),这可能是由于酸蚀增强了种皮透性,促进了种子吸水。与对照组相比,完整种子、酸蚀种子和刻伤处理种子的种仁含水量分别比对照高出25.84%、35.03%和28.69%,酸蚀处理的种仁含水量最高(41.65%),与完整种子差异极显著(P<0.01); 完整种子含水量(32.46%)与刻伤后的种仁含水量(35.30%)无显著差异(P>0.05)。
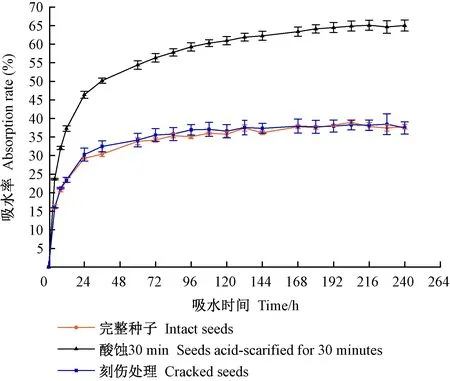
图3 不同种皮处理对池杉种子吸水率的影响Fig.3 Effects of different coat treatments on water absorption rate of T. distichum var. imbricatum seeds
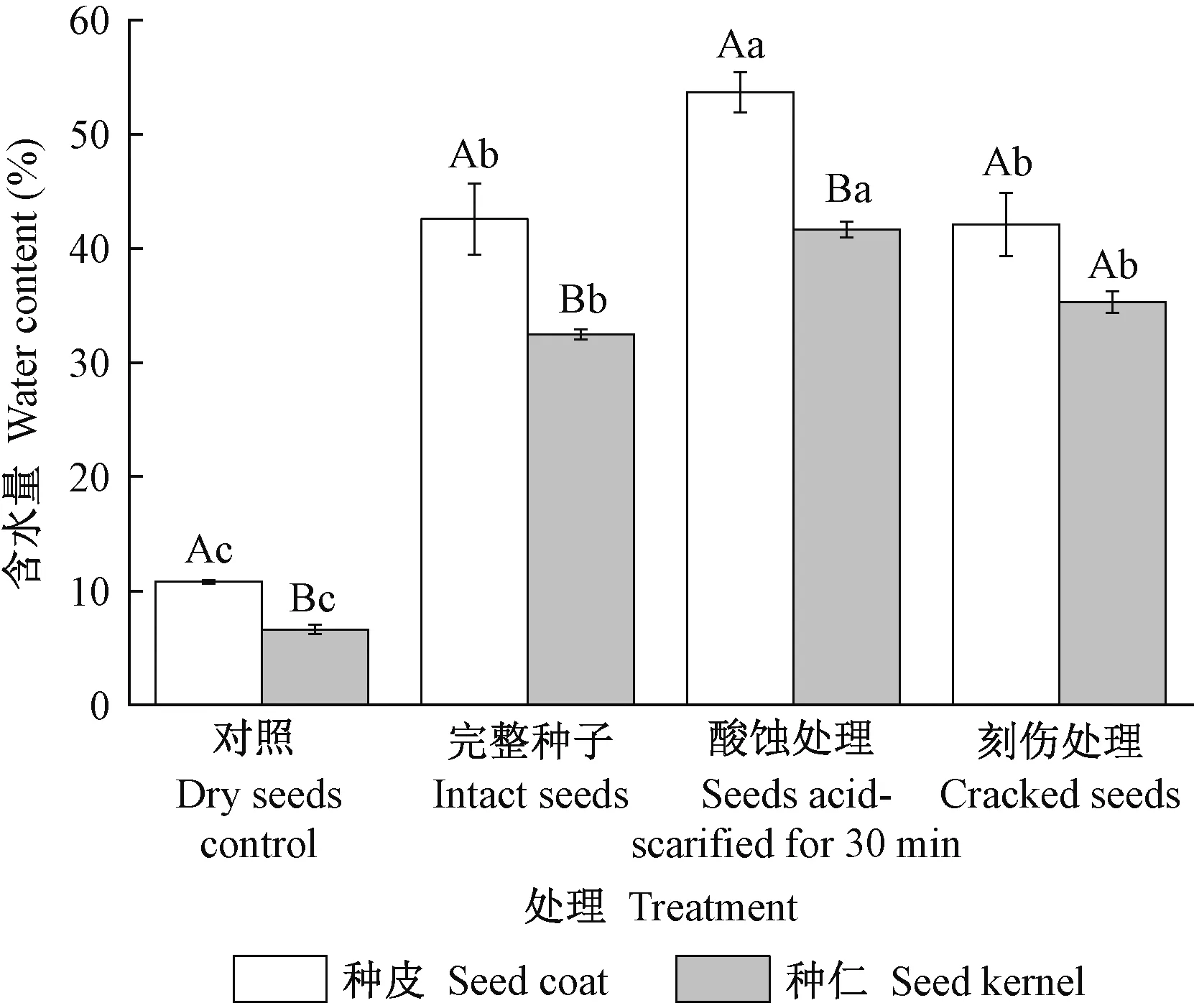
图4 不同处理种子的种皮、种仁含水量差异Fig. 4 The effects of different treatments on water contents of seed coat and kernel不同大写字母表示相同处理不同部位有显著性差异,P<0.05; 不同小写字母表示相同部位不同处理有显著性差异(P<0.05)。下同。The different capital letters indicate significant differences in different parts of the same treatment, P<0.05; the different lowercase letters indicate significant differences in the same parts under different treatments (P<0.05). The same below.
2.5 池杉种子各部分甲醇浸提液对白菜幼苗苗高和根长的影响
由图5可知,在浓度0.2和0.4 g·mL-1的种子浸提液中,白菜苗高均值分别为对照(12.49 mm)的79.77%和 76.40%。说明池杉种子的浸提液对白菜幼苗生长有抑制作用,且抑制作用随浓度增大而增强。在浓度0.2和0.4 g·mL-1的种皮浸提液浓度中的白菜苗高分别为分别为对照(12.49 mm)的81.88%和77.76%,与种子浸提液无显著差异(P>0.05); 但在浓度0.2和0.4 g·mL-1的种仁浸提液下,白菜苗高分别为对照的90.83%和87.65%。由此说明,池杉种子不同部位的甲醇浸提液对白菜幼苗生长的抑制作用不同,表现为种皮的抑制作用高于种仁,抑制作用随提取液浓度增大而增强。
在0.2和0.4 g·mL-1的种皮浸提液下的根长分别为分别为对照(33.67 mm)的86.21%和62.51%; 在0.2和0.4 g·mL-1的种仁浸提液下的根长分别为对照的134.60%和97.26%。方差分析和多重比较结果表明: 0.4 g·mL-1的种子浸提液抑制作用与种皮浸提液有极显著差异(P<0.01),与种仁浸提液无显著差异(P>0.05)。以上结果说明,种皮对白菜根长抑制作用最强,且抑制效应随浓度增大而增加,种仁未表现出明显抑制作用。由此可初步证实池杉种子含有内源抑制物质,且基本存在于种皮。
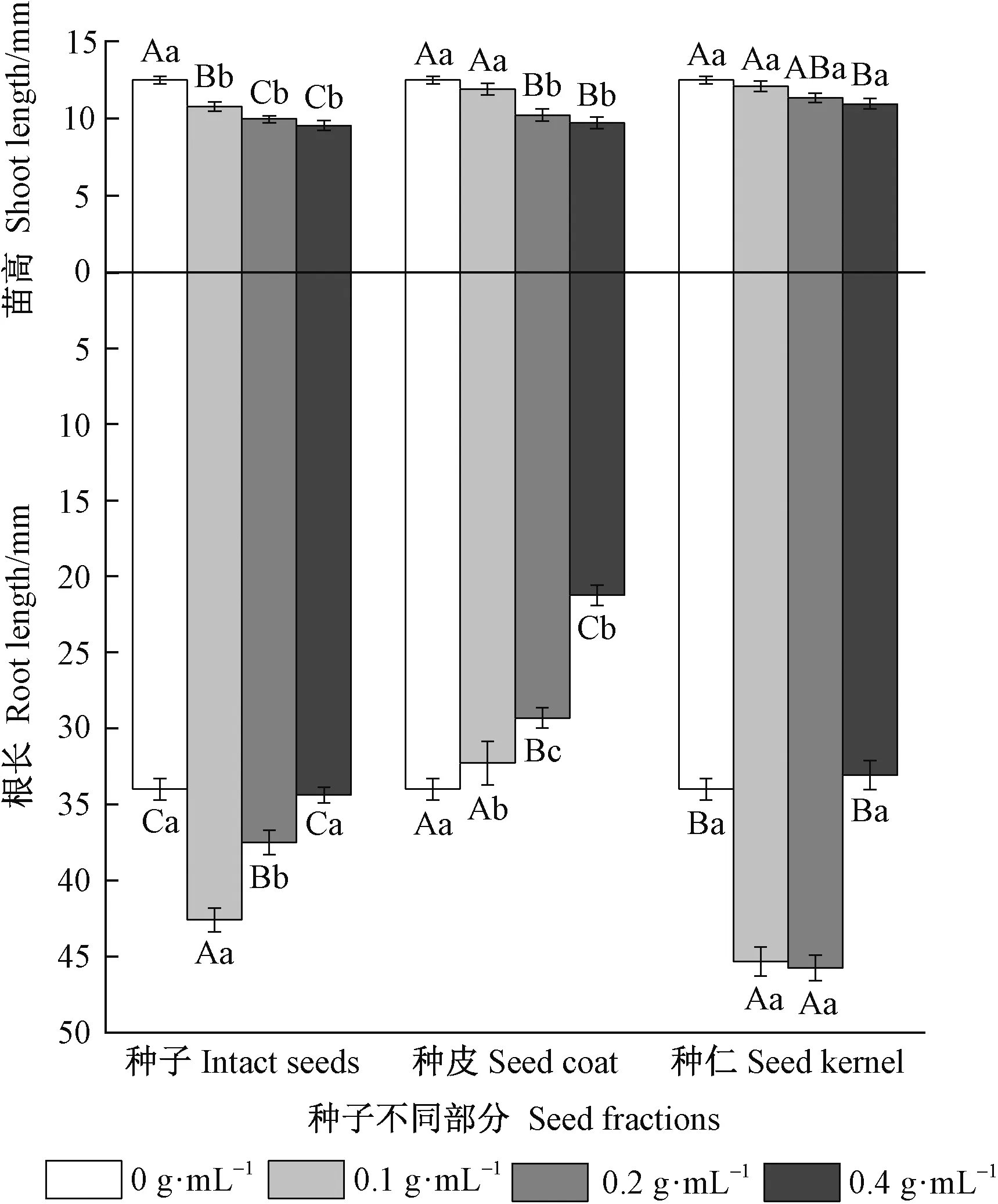
图5 池杉种子不同部分甲醇浸提液对白菜苗高和根长的影响Fig.5 The effects of methanol extracts from different fractions of T. distichum var. imbricatum seeds on shoot length and root length of Brassica pekinensis
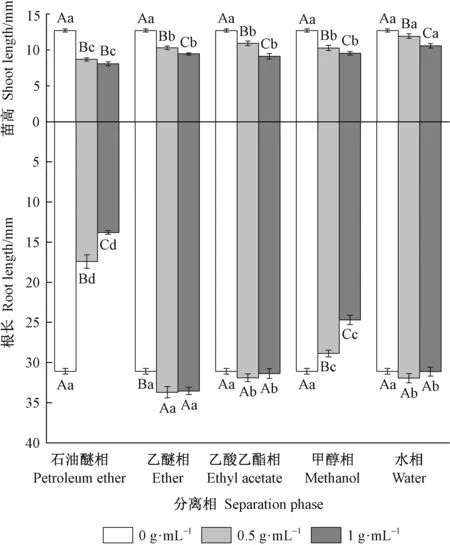
图6 池杉种皮各溶剂相对白菜幼苗苗高和根长的影响Fig.6 The effects of different separation phase of Taxodium distichum var. imbricatum seeds coat on shoot length and root length of Brassica pekinensis
2.6 池杉种皮各溶剂相对白菜幼苗苗高和根长的影响
由图6可知,在0.5 g·mL-1的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相下的白菜平均苗高分别为对照(12.70 mm)的68.39%、81.05%、85.92%、80.91%和93.81%,各分离相抑制强度表现为石油醚相>甲醇相>乙醚相>乙酸乙酯相>水相。1 g·mL-1的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相下的白菜平均苗高分别为对照的63.54%、74.37%、71.76%、75.06%和83.33%,各分离相抑制强度表现为石油醚相>乙酸乙酯相>乙醚相>甲醇相>水相。方差分析和多重比较表明,石油醚相与其他各相呈极显著差异(0.5 g·mL-1时F=14.692,P<0.01; 1 g·mL-1时F=10.329,P<0.01)。以上结果说明,池杉种皮甲醇浸提液分离出的石油醚相、乙醚相、乙酸乙酯相、甲醇相以及水相对白菜幼苗苗高生长均表现出不同程度的抑制作用,抑制作用随浓度提高而增强,且石油醚相中的抑制物质抑制活性最强。
在0.5 g·mL-1的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相下的白菜平均根长分别为对照(31.09 mm)的56.00%、108.40%、102.60%、92.89%和102.78%,各分离相抑制强度表现为石油醚相>甲醇相>乙酸乙酯相>水相>乙醚相。在1 g·mL-1的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相下的白菜平均根长分别为对照的44.36%、107.90%、100.89%、79.40%和100.16%,各分离相抑制强度表现为石油醚相>甲醇相>水相>乙酸乙酯相>乙醚相。方差分析和多重比较表明,石油醚相与其他各相呈极显著差异(0.5 g·mL-1时F=105.284,P<0.01; 1 g·mL-1时F=250.853,P<0.01)。由此说明,乙酸乙酯相、水相和乙醚相未表现出明显抑制作用,但幼苗根长随浓度增大有下降趋势。对白菜幼苗根生长表现出抑制作用的主要为石油醚相和甲醇相,且抑制作用随浓度增大而增强。池杉种皮中抑制活性强的抑制物质主要集中于石油醚相。
4 讨论
4.1 种皮结构和种子休眠关系
种子萌发的第一步是吸水,只有充分吸水,才能引起相关酶的活化,进而增强种子的呼吸作用和同化速率(赵群金等, 1998; 颜升等, 2014)。种皮是最先接触到水的结构,种子的外源休眠往往与种皮有关,了解种皮结构、测定种皮透性有利于了解休眠原因(尹黎燕等, 2002)。池杉种皮厚且坚硬,占据了种子大部分质量,且被覆蜡质,外种皮细胞排列紧密,内种皮含有木质素、蜡和黄褐色色素(Renetal., 1999),但由于外表皮易脱落,中种皮分布大量纹孔等原因,推测池杉种皮对水分吸收具有一定阻碍,但不会完全抑制种仁的吸水。热水处理和酸蚀处理是增强种子透性的有效方法,不仅可以使种皮龟裂、软化,还能去除种皮表面的蜡质和油脂,改善种皮的透性,促进种子萌发(杨期和等, 2006)。但本试验的热水处理未对池杉种皮的微型态造成影响,细胞排列和种皮结构无明显变化,说明热水处理对促进池杉种皮透性无明显作用。在酸蚀处理后,种皮表面出现的裂痕和腐蚀坑有利于增强种子的透性,且酸蚀处理会使种皮变薄,胚根萌出的机械束缚减弱,有利于种子吸涨和萌发。但浓硫酸具有强腐蚀性,处理时间过长易对种胚造成伤害,本试验中酸蚀30 min未对种胚造成损伤,但是否可以进一步延长处理时间还有待进一步研究。
池杉种皮存在一定透水性,无论种皮是否破损,种仁均能吸收到部分水分。在吸水试验结束后的解剖过程中发现,与完整种子和刻伤处理种子相比,酸蚀处理的种子更易沿种脊分为两半,且与完整种子相比,酸蚀处理的种仁含水量高,这可能是由于酸蚀处理使种皮变薄,削弱了种子的机械束缚,从而促进了种子进一步吸水膨胀。吸水96 h时,完整种子已经接近饱和,而酸蚀种子还在缓慢吸水的现象。由此推测,池杉种皮的透水性可能不是影响种子萌发的主要因素,但厚且坚硬的种皮可能对种胚吸涨产生了严重的机械阻力,进而可能抑制胚根的生长和伸长,造成种子萌发迟缓。这与青钱柳(Cyclocaryapaliurus)(杨万霞等, 2005)、华西小石枳(Osteomelesschwerinae)(李佛莲等, 2016)、欧洲冬青(Ilexaquifolium)(贾有伟等, 2017)和白皮松(Pinusbungeana)(Guoetal., 2018)等的研究结果相似,其种皮均有一定透水性,但坚硬种皮的机械障碍对种子萌发产生了一定抑制作用但与枳椇(Hoveniaacerba)(李英等, 2014)、东京野茉莉(Styraxtonkinensis)(赵婕等, 2019)和合欢(Albiziajulibrissin)(陈丽等, 2019)等的研究结果不同,这些物种种子的种皮结构致密,存在透水障碍、透气性差等问题,是导致种子休眠的主要原因。本试验表明池杉种皮所产生的机械束缚使种子可能具有物理休眠特性。
4.2 内源抑制物与种子休眠关系
内源抑制物质是随着植物生长发育过程而产生的一系列天然化合物,影响细胞的分化、分裂、伸长和发育,最终推迟或抑制同种或异种植物种子发芽。抑制物的存在是引起植物种子休眠的主要原因之一(Kentzer, 2015)。喻梅等(2012)研究发现野生鸭儿芹(Cryptotaeniajaponica)种子中存在抑制物,其粗提液在较低浓度下即可抑制其种子萌发; 李庆梅等(2013)发现栎类(Quercus)种子中含有抑制物质,其主要抑制种子萌芽和生长; 闫芳等(2016)认为黄瑞香(Daphnegiraldii)种子有内源抑制物,尤其是种胚是引起种子休眠的主要原因。在本研究中,对照条件下白菜种子萌发率高且生长迅速,而池杉种子浸提液对白菜苗高和根长有一定抑制作用,说明池杉种子含有内源抑制物质,能抑制异种植物种子萌发。将池杉种子分为种皮和种仁两部分,分别进行甲醇浸提和白菜苗高和根长的生物测定后表明,抑制物质存在于种皮中。
种子内部存在的抑制物质种类多样,常见的有氰化氧、酸类、香豆素、ABA、乙烯、酯类、芥子油、生物碱、芳香油等(Thomasetal., 2000; 刘艳, 2007),其种类不同,极性也有可能存在差异,因此通过系统溶剂法进行分离后,抑制物质就分别集中在各有机相中。黄耀阁等(1998)、张艳杰(2007)采用系统溶剂分离法分别对西洋参(Panaxquinquefolius)果实、南方红豆杉(Taxuswallichianavar.mairei)种子的浸提液进行分离,发现西洋参果实抑制物质主要存在于乙醚相和甲醇相中,南方红豆杉的抑制物质则主要集中在种皮的甲醇相中。尚旭岚等(2011)采用系统分离法对青钱柳(Cydocaryapaliurus)果皮的浸提液进行分离,发现果皮的乙醚相中含有的香草酸等抑制物质是导致青钱柳休眠的主要原因。本研究中的池杉种皮各分离相的生物测定结果表明,种皮甲醇浸提液的各分离相对白菜幼苗苗高和根长表现出不同程度的抑制作用,其中无论苗高还是根长,石油醚相的抑制作用最强,这说明池杉种皮中抑制作用较强的物质主要集中于石油醚相中,即池杉种皮内的抑制物质可能是一些极性较小的物质。这些物质可能是导致池杉出苗不整齐的主要原因之一,由此说明池杉种子具有生理休眠特性。但内源抑制物质的成分鉴定还有待通过高效液相色谱和质谱联用等方法进行分析,今后研究中可对池杉种皮浸提液各分离相对种子自身的抑制作用进行探索,以期揭示池杉种子的休眠机制。
5 结论
池杉种子具有显著休眠现象。池杉种皮的透水性不是抑制种子萌发的因素,种皮的机械障碍和抑制物质可能是造成池杉种子休眠的主要原因,且这些抑制物质可能是极性较小的物质,因此池杉种子是兼具物理休眠和生理休眠的复合休眠。
