中蜂形态鉴定与分子生物学遗传特性研究进展(1)
2022-07-20徐祖荫
徐祖荫
(贵州徐祖荫养蜂工作室,贵州 贵阳 550001)
中蜂是我国主要蜂种之一,约占全国蜂群总数的50%。20世纪70~80年代,国内学者对中蜂进行形态测定,并据此将分布于我国境内的中蜂划分为9个生态类型。
长期以来,蜜蜂的分类和系统进化一直是蜜蜂生物学的研究重点之一。从20世纪60年代开始,德国的鲁特涅(Ruttner)、Koenige等,美国的Peng、Smith等就开始对蜜蜂的分类、系统进化等方面进行了广泛的研究。1988年,鲁特涅经过长期研究、提出测定蜜蜂的38个形态指标,是迄今为止国内外比较公认也最常用的形态指标测定标准。
大概在20世纪70年代后期和80年代前期,我国学者开始对分布于我国的东方蜜蜂(中蜂)开展大规模分类、分布、进化等方面的研究,这个时间点恰好也是国际上测定蜜蜂形态指标逐渐成熟和完善的时期(如Ruttner)。受当时工作条件(如经费、检测手段等)的限制,所选择的形态指标有限(主要突出经济性状及主要的形态特性,如吻长、3+4背板长、第四背板突间距、第三腹板后缘宽、翅长、翅宽、小盾片颜色、体色色型等),测定数据未能细加整理并完整、充分地反映在代表性的论文中,未能引起国外同行的重视,以致国际上蜜蜂分类权威如鲁特涅、Peng等为此感到缺憾。尽管如此,众多科技工作者(以杨冠煌、匡邦郁等为代表)对东方蜜蜂在我国的分布、分类、进化等方面做了大量工作,基本研究得出了我国中蜂种下分类的具体框架,经我国畜禽资源委员会确认,载入《中国畜禽遗传资源志——蜜蜂志》中,并广泛用于指导生产实际。
20世纪与21世纪交替之际,国际上蜜蜂分类应用研究,已经上升到分子鉴定水平。尽管如此,形态分析仍然是研究蜜蜂不可或缺的一个重要手段,是蜂种划分的一个重要指标。蜜蜂在长期进化过程中形成了与环境相适应的形态结构,了解蜜蜂的外部形态、内部构造及其功能,对深入掌握蜜蜂的生物学特性、分布活动规律、分类鉴定、遗传育种、提高饲养管理水平等方面都十分必要。随着现代科技进步,一些微小生物形态测定仪的出现,大大提高了形态测定的速度和精度,减轻了测定的工作量。近20年来,我国学者也将分子生物学、几何形态测量学运用到中蜂种群分布、演化规律的研究中,取样范围几乎囊括了我国中蜂分布的全部地区,取得了一系列重要成果,进一步印证了第一次中蜂资源普查结果,从多个角度深化了人们对中蜂遗传、分化规律的认识,现归纳如下。
1 我国境内的中蜂已分化为不同的地理生态类型,不但不同的地理、生态类型之间,即使在同一类型之间,从形态特征到基因检测,均不完全相同
物种分类的基础理论是生物进化论。世界著名的昆虫学家、进化分类学家、中国科学院院士、原中国科学院昆虫研究所、动物研究所所长陈世骧提出了“居群”、“又变又不变,又连续又间断”的物种概念。陈老指出:每一物种都有一定的生活习性,占有一定的分布区域。在它们的分布区域内,有可能生存的场所与不能生存的场所彼此相互交替着。因此,每一物种在其分布的空间内,形成分散的、不连续的、大小不同的群体单元,称之为居群或种群。同群的个体因经常相遇,所以成为实际的繁殖单元。群与群之间也可以通过迁移杂交而实现基因交流。距离越远,交流的机会也越少。相距遥远的居群(种群)不可能直接交流,因而分化为不同的地理类型或生态类型。居群(种群)分化是物种分化的前奏,居群是分散的,又是联系的,物种是由既连续又间断的居群所组成(图1)。居群(种群)是变异的,正如个体变异一样,同种间的居群(种群)没有两个完全相同的。
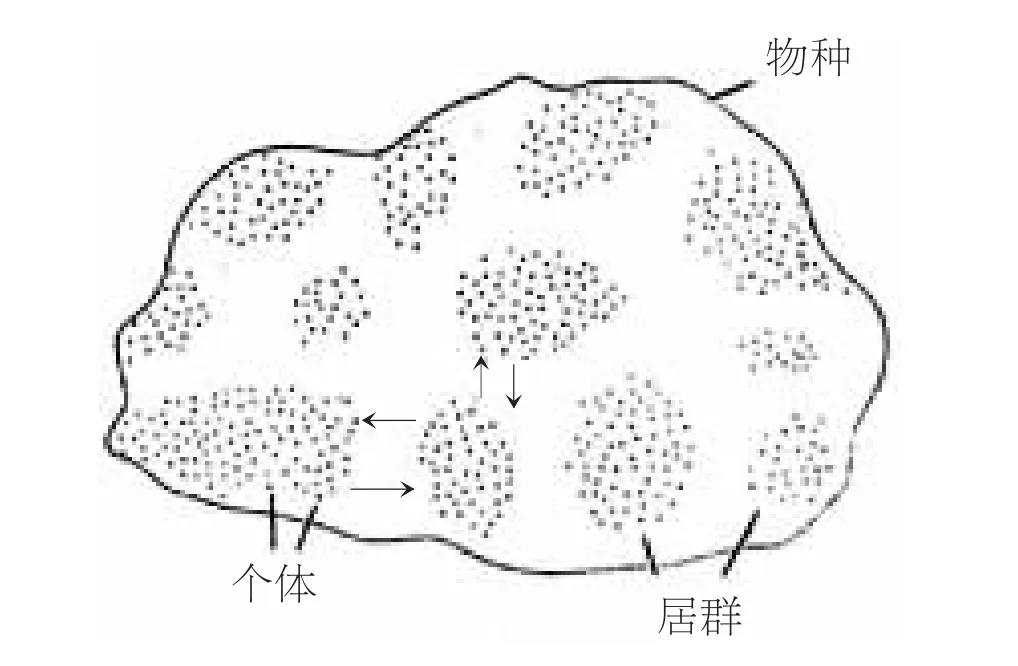
图1 物种的群体结构示意图(箭头表示群间交流)(引自陈世骧)
自首次公布全国中蜂资源调查结果以来,不断有学者继续对其进行深入研究。尤其近20年来,采用改进后的形态鉴定(几何形态测量学)方法和分子生物学手段,对国内不同地区的中蜂开展了大量的相关研究,结果表明,不同地区的中蜂,即使在已知的同一生态类型之间,都具有其相对独立的形态特征和分子遗传标记(蒋滢,1997;王瑞武,1998;董霞等,2001;谭垦等,2002;石巍等,2004;苏松坤等,2004;姜玉锁,2007;吉挺等,2008、2009;陈晶等,20 08;刘之光等,2008;沈飞等,2008;丁桂玲等,2008;朱翔杰、周冰峰等,2011;周姝婧等,2012;徐新建等,2012;于增源等,2012;陈伟文等,2013;张祖芸等,2017;曹联飞等,2017)。
曹联飞等(2017)通过对浙江省丽水市中蜂样本基于线粒体DNA tRNA leu—COⅡ进行序列测定,共发现20个单倍型。其主体单倍型占41.5%,与已报道的福建中蜂主体单倍型及广州中蜂序列相同,支持将丽水中蜂划为华南生态型,但丽水中蜂第二大单倍型分布频率与福建中蜂明显不同,并同时报道了2个新发现的单倍型,提示丽水中蜂还有其独特的遗传背景。
邱勇等(2021)在云南省保山市昌宁县4个乡镇(翁堵、鸡飞、漭水、田园)采样,以线粒体DNA上的细胞色素氧化酶亚基1基因(cox-1)片段为分子标记,利用生物信息学软件对上述同县不同乡镇的中蜂(应为同一生态类型)进行遗传学特性分析,发现它们既有共有的单倍型,也有一定比例独立的单倍型,其独有单倍型率为28.6%~50%,各自具有相对独特的遗传结构。核苷酸多样度(pi)监测表明,4个地理种群在0.00150~0.00174之间波动,表明其内部遗传差异化程度虽然不大,但仍存在着一定的遗传分化(弱分化)。
谭垦等(2002)对来自国内外东方蜜蜂mt DNA COⅡ基因420个碱基对测序分析,即使来自同一个地区2个蜂群之间,如北京的China 1和China 2、云南河口的China 6和China 8,云南撒营盘的China 3和China 4,其碱基序列位点变异分别为3个、2个和2个,差异率分别为0.7%、0.4%和0.4%。上述研究结果,完全印证了“同种间的居群,没有两个完全相同”的观点(陈世骧,1977)。
2 分布于我国境内的中蜂,随纬度北移,海拔升高、年平均气温降低,普遍规律是中蜂体型变大、体色变深
谭垦等(2001,2005)先后对采自云南省14个不同地区、不同生态环境样点,以及对采自北京、吉林、甘肃、湖南、湖北、云南、四川、安徽等省市634个样点的样本,参照鲁特涅1988年提出的标准进行测定和统计分析。结果显示,相对于低纬度、低海拔、气温较高地区的蜜蜂来说,高海拔、高纬度、气候比较寒冷的地区的蜜蜂体型较大,颜色也较深。这种形态上的变化,对蜜蜂适应其所生活区域内的环境,具有重要的生物学和生态学意义。在高寒湿冷的地区,蜜蜂为了御寒,需要增大摄食量和采集更多的食物,反映体型大小的指标如足、翅、背板、腹板、蜡镜间距等的大小也随之增大。体色变深有利于吸收热量、保温防寒。这一研究结果与Verma(1996)在研究印度东方蜜蜂地理形态趋化,Rutther(1988)和Stefen fuch(2000)研究北纬36°西方蜜蜂的地理形态趋化,Hepburm研究高山和平原非洲蜜蜂形态变化地区的结论是一致的。此后,谭垦等(2004)对甘肃中蜂,王桂芝等(2008)分别对山西、河南两省中蜂的测定,也都得出相同的结论。甚至在地理范围不太大的区域内,如湖北神农架(谭垦等,2005)都具有相同的趋势。
李华等(2008)对采自青海民和县的中蜂检测时还发现,青海中蜂不但个体较大,且覆毛和绒毛都比较长,这与其高纬度、高海拔、低气温的生境有关。蜜蜂体表密生的绒毛,除能黏附大量花粉粒外,还具有护体和保温的作用。
王桂芝(2009)对采自黄河中下游地区(包括北京、山东、山西、河南、河北)14个样点中蜂8个与色型相关的指标分析,认为工蜂的体色除与纬度、海拔、年平均气温等因素有关外,还与日照时数、太阳辐射以及大气水分有关。例如处于黄土高原东部、位于太行山系两侧、海拔1 000 m、紫外线辐射强的亚干旱地区,蜜蜂体色较深,多为黑色;而在山东平原及近海的亚湿润地区,季风活跃,光、温、水资源适宜,蜂体体色就较亚干旱地区偏浅,一般呈浅棕或黄色。
