海南清澜港红树植物海莲种群结构与分布格局研究
2022-07-12李茂金叶储民陈玉凯
李茂金,叶储民,陈玉凯
1.海南省林业集团有限公司,海南海口 570205;
2.热带岛屿生态学教育部重点实验室,海南师范大学生命科学学院,海南海口 571158
种群结构不仅可以反映种群个体属性特征的数量分布状况,也可以反映种群数量动态及种群的发展趋势,同时可以在很大程度上反映种群与生境间的相互作用关系,以及在群落中的生态作用和生态位。种群的分布格局与种群自身的特性有极大的关系,同时也受种群间效应、生境条件的直接影响。通过对群落中种群分布格局的调查,可以揭示群落空间结构的特征,有助于了解种群的某些生物学和生态学特性,以及解释一定环境因子对种群行为或者种群生存的作用。调查得出的分布格局结论和数量特征指标,可以作为确定抽样方式、数据处理提供理论基础。在生产和开发利用上,对于红树林的经营管理以及混交林的配置具有重要参考价值[1-2]。
海莲隶属红树科木榄属,常绿乔木,是红树林东方类群树种,在中国境内,海莲仅分布在海南省,近年来引种于广东省深圳海滩;在世界上它分布于印度、越南、泰国、缅甸、菲律宾、印度尼西亚等国家的湿热海滩。
目前对海莲的研究已有较多的报道,如,从生理生态方面的研究有:郑文教[3]等研究了盐度对海莲幼苗的生长和某些生理生态特性的影响;马丽[4]等研究了铝胁迫对海莲幼苗保护酶系统及脯氨酸含量的影响。从种群和群落方面的研究有:王佳燕[5]等对海南东寨港海莲群落林隙结构及其更新特征的研究,结果表明:海南东寨港河港海莲群落最主要的干扰类型是树木的大量枯死;林隙幼苗密度低于林下幼苗的密度,而林下幼苗的高度低于林隙幼苗的高度,由此可知,适当的林隙可以促进林分更新;廖宝文[6]等对海南岛东寨港几种红树植物种间生态位的研究表明:作为红树科的植物种之一,海莲有其特殊的生长环境要求,只适生于底质较为硬实的半泥沙质的高潮滩涂,海莲生态位宽度值最小,高潮滩滩涂植物海莲、角果木与其它树种的生态位重叠值均相对较低(其他树种的生境可能与海莲、角果木的生境比较相似的原因),与实际情况相符。海莲这一红树植物为嗜热窄布种,一般生于潮水较浅、土质比较硬实、阳光充足的泥沙滩涂生境,主要分布在热带红树林林区的高潮滩涂带,与中低潮滩的红树植物占据相近或相似生态空间的程度不大,所隐含的竞争现象也比较小;郑松发[7]等对海莲群落和木榄群落主要种群分布格局的研究表明:海莲种群在某种程度上具有集群性,但是强度较低,可视作随机分布和集群分布的过渡类型,分布格局类型主要与各自的群落内生境的微地形变化有关。
海莲种群是海南省文昌市清澜港红树林林区内的主要优势种群之一,该研究的目的,是通过探讨清澜港红树林林区内优势植物海莲种群的结构特征、分布状况及其发展趋势,阐明海莲种群空间分布格局与环境关系,以期为海莲及其他优势红树种群的人工经营及种群的合理保护提供参考依据。
1 研究区概况
海南清澜港红树林自然保护区是中国最大的红树林区之一,保护区总面积2948hm2,其中红树林面积1223.3hm2,位于海南省东北部文昌市界内,地理坐 标 为19°15′N ~20°09′N,110°30′E ~110°02′E,属热带季风海洋性气候[8]。区域内的清澜港港湾深入内陆,形成了内宽口窄的漏斗状,在文昌江和文教河汇入湾内,沿岸积累的淤泥深厚,风浪微弱,为典型的泻湖-河口湿地生境,很适宜红树林的生长。
文昌清澜港红树林保护区主要分布在八门湾的沿岸滩涂带,并且断断续续地分布到港湾外,港湾沿岸陆地为沿海台地地貌,原生的植被类型是热带雨林,但受人类经济活动干扰,绝大部分已开发改成为虾塘、椰树、桉树林、木麻黄林及其他果树林[8]。
2 研究方法
2.1 取样方法与野外调查
2020 年8 月到9 月,根据海南文昌清澜港红树林自然保护区海莲种群的结构现状及其生境特点,共选择了3 个样地进行调查研究,编号分别为P1、P2、P3,样地概况见(表1)。其中,样地1 位于八门湾内与清澜镇交界处,样地2 位于八门湾内与东郊镇交界的沿海台地,样地3 位于文昌北部的铺前镇珠溪河沿岸。该研究以海莲种群为目标物种进行调查,在调查区域内,由于红树林多为带状分布,植物种类也较少,样方面积不好做大,因此三个样地取的样面积为900m2,共2700m2,将每个样地分成50m2的棋盘格子样方,各18 个,并采用每木调查法,测量每个样地内树种的个体的相对坐标、胸径、冠幅、高度等特征。

表1 样地概况Tab.1 Plot Characteristics
2.2 种群结构的研究方法
根据海莲种群的生长特点,对各样地中海莲种群每木调查所得的数据进行分组统计,其中冠幅直径的组间距为1.0m,高度的组间距为2.0m[9-10],由于种群径级结构的相对稳定性类似于年龄结构的稳定,所以许多学者都采用胸径结构来代替年龄结构进行分析乔木种群的结构及其动态特征[11]。该文对海莲的胸径结构进行3 个年龄级的分析,即(幼龄)0<胸径≤7.5cm、(中龄)7.5<胸径≤22.5cm、(老龄)胸径>22.5cm 以上。
2.3 种群树高、胸径和冠幅的相关性分析
通过对样地进行每木调查,得到清澜港红树林海莲种群冠幅直径、胸径、树高的基本实验数据,对数据进行处理,首先通过利用excel 分别作出冠幅直径与胸径、树高与胸径的散点图,然后根据所得各散点图结合5 种研究模型[12-13],对冠幅直径与胸径、树高与胸径进行相关性分析;利用SAS 软件对各组调查所获得的数据进行模型拟合后,得到各方程的待定参数及各回归方程及其方程决定系数R2,以决定系数R2大小来评价其拟合程度大小,R2小,说明拟合程度差,模型则不够可靠。反之,R2大说明拟合程度好,模型可靠。
2.4 种群分布格局分析
以各样地内海莲个体数据为依据,对三个样地调查得到的数据进行方差均值 (亦称扩散系数)的t 检验,以二项分布的聚块指数(m*/m)、平均拥挤度(m*),Morisita 指数的F 检验,判定海莲种群不同样地和不同年龄级的空间分布格局[14-15]。
3 结果与分析
3.1 海莲种群结构
由表2 可知,海莲种群的个体高度主要集中在0m~6m,占67%,其中各样地的优势高度以及个体比例分析结果如下:在样地P1 中,高度在6m~8m 范围的个体占65%;在样地P2 中,高度在2m 以下的个体占58%;在样地P3 中,高度在2m 以下的个体占26%,2m<高度≤4m、4m<高度≤6m 和6m<高度≤8m 的个体所占比例也较多,分别是24%、22%和22%。这三个样地的自然环境不同,各样地的整体外观有所差异,样地P1 处于中龄的植株较多,样地P2 处于幼龄的植株较多,所以两地的高度有一定的差异,样地P3 各个年龄的植株都有一定的数量,高度差异介于样地P1 和P2 两者之间。

表2 各树高级分布Tab.2 Distribution of Different Tree Height Grades
由表3 可知,海莲种群的个体胸径主要集中在2.5cm~7.5cm,占63%,经过分析,各样地的优势胸径及个体比例如下:在样地P1 中,胸径大小在7.5cm~22.5cm 范围内的个体占54%;在样地P2 中,胸径大小在7.5cm 以下的个体占84%;样地P3 中,胸径大小在7.5cm 以下的个体占59%。由此可知,在三个调查样地中,样地P1 的中龄海莲植株所占比例是最大的,说明在过去一段时间内,P1 样地林分发生变化,P1 样地中海莲种群呈现了增长趋势,老龄和幼龄植株相对较少,种群的年龄结构格局趋向于中间大两头小,P2 样地中海莲种群的自然更新能力最强,样地中存在大量的幼龄植株,老龄和中龄植株数量很少,说明样地P2 的环境相对较适合海莲的更新和发育,样地P3 的中老龄植株和幼龄植株比例相差不大,种群表现出较稳定的发展趋势。

表3 各胸径级分布Tab.3 Distribution of Different DBH Grades
由表4 可知,海莲种群的个体冠幅直径主要集中在0m~3m,占66%,其中各样地的优势冠幅直径及个体比例如下:样地P1,4<冠幅直径≤5m 和冠幅直径>5m 以上的个体较多,均占30%;样地P2,1m 以下的个体占54%;样地P3,1m~2m 的个体占52%。样地P1、P2、P3 的冠幅直径最大值的个体分别是6.5m、7m、5m;样地P1 和P3 分别缺失0m~1m 和4m~5m 的个体。

表4 各冠幅直径级分布Tab.4 Distribution of Different Crown Diameter Grades
由此可知,样地P2 的自然环境条件更有利于海莲种群的自然更新,而样地P1 的自然环境条件对海莲种群发展不利,有趋向中间大两头小的格局,这也说明了不同生境条件和不同立地条件对海莲种群的作用强度以及种群适应特征有所不同,海莲种群结构的差异与区域环境特点有较大的相关关系,由此反映出生境差异对种群结构的影响作用。
3.2 海莲种群树高、冠幅直径和胸径的相关性分析
3.2.1 种群树高与胸径的相关性分析
由图1 分析可以看出,海莲的树高具有随着胸径增大而增大的现象,观察数据可知,树高高度主要集中在3m~6m 的范围内,相应的胸径大小范围在10cm 以内,说明所调查的海莲林分年龄偏低;经相关数据分析,海莲树高与胸径之间有显著相关性,相关系数为0.6829,而由表5 中海莲树高与胸径各模型拟合结果可知,五种模型中y=2.4202Ln (x) +0.1578、y=0.2013x+3.2385 和y=2.8341+0.2722x-0.0020x2模型的决定系数相差不大,但模型y=2.8341+0.2722x-0.0020x2拟合效果最好,决定系数最高,利用此模型对海莲胸径估算其树高结果可靠,此模型可为调查海莲种群提供一定依据。由于红树林潮滩通行困难,此模型也可以减轻野外测高工作量,并可了解和进一步掌握海莲种群的生长发育过程。

图1 树高与胸径散点Fig.1 Scatter of Height and DBH
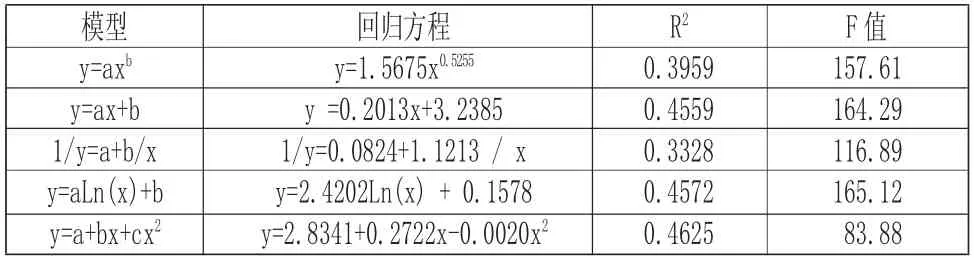
表5 树高与胸径关系数学模型及显著性检验Tab.5 Mathematical Model and Test of Significance of Height and DBH
3.2.2 种群冠幅直径与胸径的相关性分析
由图2 分析可以看出,海莲种群的冠幅直径具有随胸径的增大而增大的现象,观察数据可知,冠幅直径以1m~3m 的范围内最为密集,相应的胸径在10cm 范围内,经相关分析,海莲胸径与冠幅直径有显著相关性,相关系数为0.5492。根据表6 中海莲冠幅直径与胸径各模型拟合结果可知,五种模型中w=0.7218x0.5936和1/w=0.0997+2.7214/x 模 型 的 决定系数相差不大,但模型1/w=0.0997+2.7214/x 拟合效果最好,决定系数最高,利用此模型对海莲胸径故算其冠幅直径结果可靠。根据海莲种群的胸径与冠幅直径大小的相关关系,可以编制林分经营密度表,并且可以适时合理的对海莲林分进行抚育间伐。并通过对海莲种群的冠幅直径与胸径生长因子之间的相互关系分析,为进一步研究海莲林种群结构、自然演替和生长发育规律等生态学规律提供可靠的依据。

图2 冠幅直径与胸径散点Fig.2 Scatter of Crown Diameter and DBH
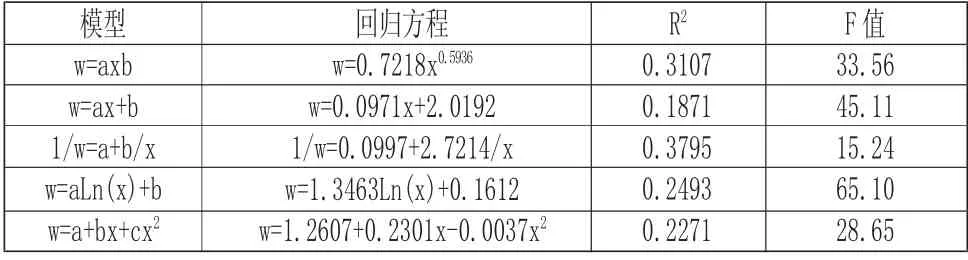
表6 冠幅直径与胸径关系数学模型及显著性检验Tab.6 Mathematical Model and Test of Significance of Crown Diameter and DBH
3.3 海莲种群分布格局
种群的空间格局也是种群结构的重要特征之一,它反映的是种群在水平空间上的配置格局或者是分布状况,其形成原因是由种群特性、种间关系以及种群所在的环境条件共同起作用所决定。
由表7 可知,所调查研究的三个样地中,海莲的种群都呈现出集群分布的状况,其中样地P2 的平均拥挤度以及聚块指数都达到了最高,说明样地P2 的这种分布格局在一定程度上有利于种群形成幼苗生长的小环境,同时这种分布格局也可以增强种群抵抗其它植物入侵的抗性,增加种群在群落中的竞争力以及种群对营养资源和生存空间资源的占据与巩固。

表7 不同样地海莲种群分布格局的测定结果Tab.7 Result of Spatial Distribution Patterns of Bruguiera sexangula Population in Different Plots
由表8 可知,海莲种群在不同发育阶段,种群的分布格局大体上呈现出集群型向随机型转变的趋势,并且各集群强度由大变小,趋向于减弱,种群中幼龄树集群分布,且聚集程度很高;中龄树为集群分布,聚集程度有所下降,老龄树逐渐趋于随机分布。但样地P3 的幼龄聚集强度较弱,明显低于P1 和P2,这是因为样地P3 已受到人为的干扰,而幼龄个体更易受人类活动的影响,因此有趋向随机分布的可能,而P2 的幼龄聚集强度非常高,明显高于P1 和P3,这种格局的形成是因为样地P2 的幼龄植株个体数量比较多,样地P2 中可以经常见到十几株簇拥生长。

表8 不同发育阶段海莲种群分布格局的测定结果Tab.8 Result of Spatial Distribution Patterns of Bruguiera Sexangula Population at Different Age Stages
4 结论与讨论
4.1 结论
海南清澜港红树林林区内的海莲种群各级树高的个体数分布较均匀,但不同样地间的各级高度个体数分布仍有一定的差异;海莲最大冠幅直径的个体是7m,66%个体的冠幅直径大小在0.5m~3m 范围内;63%的海莲个体的胸径大小在2.5cm~7.5cm 范围内,种群整体的自然更新能力较强。运用5 种研究模型对海莲胸径与树高和冠幅直径之间的关系进行拟合,均达到了极显著水平;目前海莲种群都基本属于聚集型分布,不同发育阶段种群的分布格局大体上表现为集群型向随机型转变;但人为活动干扰对海莲种群幼龄树的分布格局影响较大。
4.2 讨论
通过比较不同环境的海莲种群树高、胸径和冠幅直径结构特征,可以反映不同环境和立地条件对海莲种群的作用强度和种群适应特征,不同样地的海莲种群结构的差异很可能与局部气候、地形、土壤条件等区域环境特点有相关关系,反映出生境差异对种群结构的影响[16]。
海莲种群的树高与胸径的关系及冠幅直径与胸径的关系用不同的数学模型方程来拟合,均呈极显著相关关系,这与其他一些报道中的结果一致[13]。该文通过对海莲种群的树高、冠幅直径与胸径生长因子之间的相互作用关系分析,为进一步研究海南省文昌市清澜港红树林保护区海莲林种群结构特征、生长发育规律和自然演替规律等提供可靠的参考依据。
通过对胸径和冠幅直径的研究,不仅可以利用胸径和冠幅直径的相关方程来进行预测,在给定胸径的某一林分的人工造林密度和郁闭度时间,而且也可以在适当的重叠情况下,充分的利用林分空间,使整个林分获得最高的生产量,从而达到速生、优质、丰产的目的[12]。对红树主要树种的树高、胸径(或地径)的相关关系进行研究,也同样可以为进一步研究人工引种,或自然的红树林生长发育规律、群落特征规律等提供可靠参考依据。同时,冠幅直径的大小与胸径的大小均能体现出植物个体在种群或群落中的竞争力,二者存在显著的相关关系[17]。该研究中海莲种群的胸径和冠幅直径呈现较高的正相关,而个体数量随胸径和冠幅直径级的增加而逐渐减少,呈现较高的负相关,说明海莲种群胸径和冠幅直径越大,种间竞争越强烈,进而使植物个体生长速率下降,甚至出现死亡,这个过程体现了海莲种群的自疏过程。
在红树林群落中,由于其具有胎生繁殖的特性,成熟后的胚轴从母体上掉落,散落在母株周围的泥土中,然后由胚轴发芽长成幼苗。海莲的胚轴较长且林下有很多气生根,尽管潮水涨落,也很难将掉落的胚轴带到其他地方,因而容易形成其种群的集群分布格局。一旦幼苗长成成年个体,其树冠的高郁闭度很难让其他种类入侵成活,因此这种既定的分布格局比较难以改变。通过分析表明,清澜港的红树林优势海莲种群均趋于集群分布(表7),说明海南清澜港的整体环境很适宜海莲种群的繁衍生长。
许多研究表明,从植物种群在不同发育阶段的空间格局变化规律,可以推测成年个体的死亡、新生个体的产生以及种群遭受人为干扰的概况[18-19]。一般情况下,植物以母株为中心的繁殖方式是使低龄树呈现聚集分布的原因;随着树龄的增长,植物对营养条件、光因子的需求会加大,以及在自疏和它疏等生物作用下,种群会趋于均匀分布;随着树龄的继续增加,种群的分布格局类型也将趋于随机分布[20-21]。该研究中由于海莲种群的胎生现象使海莲幼苗成片生长在母树的周围,而且因为海莲种内幼苗之间的竞争比较小,容易形成集聚格局。在海莲种群的继续发育的过程中,个体对环境条件的要求会加剧,这便导致种内、种间的竞争加强,在竞争过程中,海莲种群会因为自疏和它疏的作用,使种群密度剧烈下降,其结果势必会使同一集群内的个体间出现分化,从而导致海莲的种群密度显著下降,集群将会解体,扩散趋势将会越来越明显;当这种变化持续到中树阶段,集群强度明显比小树的集群强度低,甚至在部分样地出现了随机分布现象。处于幼龄阶段的海莲种群呈现高程度的集群现象,有利于其存活和发挥群体效应;而成年后由于个体增大,集群的强度降低有利于个体获得更多的环境资源。由此可知,海莲种群的集群强度从幼龄到成年的变化规律,是种群的一种生存策略或者适应机制。该研究结果(表8)进一步表明,在自然条件下,海莲种群的幼苗,幼树呈现较高的集群分布,中龄树也是集群分布,但集群分布明显下降;而老龄树为随机分布;但在人为活动的干扰下,海莲种群幼龄树的分布格局较容易受到影响,因此未来要特别关注人为活动的类型和强度对清澜港海莲幼龄种群分布格局的影响。
