模拟氮沉降对毛竹林林下植被多样性及养分特征影响
2022-07-10梁金鹏蒋小凡
崔 珺,梁金鹏,蒋小凡,马 涛
(黄山学院,安徽黄山 245041)
全球多样性丧失的第三大驱动因素就是大气氮沉降[1].相较于森林群落中的乔木层而言,林下植被的多样性更易受到氮沉降的影响.长期氮沉降会导致林下层物种组成发生改变,但改变的程度取决于森林的类型[2-3]、功能类群[4]、氮素状态[5]以及效应、土地利用方式[6-7]等因素.鲁显楷[3]等在氮添加实验中首次提出了植被多样性受到土壤酸化机制的影响,其他的研究成果也支持该理论[4-5].除了林下植被多样性外,植物的元素平衡也会受到大气氮沉降的影响.氮沉降对植物的影响主要是元素失衡,这些影响有的会直接作用于植物本身,有的则通过改变土壤化学元素的组成,间接影响植物[8-9].李德军[10]总结前人研究指出氮沉降会造成植物体内养分元素比例的失衡.不同生态系统植物对氮沉降的响应不同,一方面植物对氮沉降的敏感程度因种而异[11],另一方也会受到生存环境的影响.
毛竹是我国重要经济竹种,毛竹林也是我国栽种面积最广的笋、材用竹林,而林下植被是竹林生态系统最重要的组成成分之一,对竹林的水土保持、养分循环、生物栖境供给和竹林生产力等有重要作用[12].鉴于林下植被的重要性,专家学者对于不同类型[13]、不同结构[14]和不同经营措施[15]的毛竹林林下植被进行了多样性调查,由于近年来气候变迁和营林措施的改变,许多竹木混交林逐渐转变为纯林的集约化经营导致了一段时间后生产力的下降,因此在竹林经营中常采用施肥的方式来改变衰退的林地,施肥措施则会直接对林下植被及土壤造成干扰[16].
氮沉降对毛竹林分但影响的研究主要集中在氮沉降对毛竹实生苗生长[17]、光合作用[18-19]、生物量[20]及土壤养分状况[21]等方面,但是关于毛竹林林下植被多样性及养分影响的报道较少.因此,本研究通过设置不同梯度的氮添加实验模拟氮沉降,来研究氮沉降对毛竹林林下植被多样性及养分特征的影响,了解不同施肥梯度毛竹林林下植物的物种组成与多样性及养分分析,为实现竹林的可持续经营提供数据支撑.
1 研究区概况和研究方法
1.1 研究区概况
研究区域位于皖南泾县东部蔡村镇(117°57'-118°41'E,30°21'-30°50'N),属于亚热带湿润季风气候,四季分明,春温多变,夏雨集中,秋高气爽,冬季寒冷.雨量充沛,年降水量达1 100-2 500 mm,分配不均;年平均气温为16 ℃[22].调查区域植被丰富,森林覆盖率达76.4%,林中灌木种类主要有檵木(Loropetalum chinensis)、乌药(Lindera aggregata)、紫金牛(Ardisia japonica)、枸骨(Ilex cornuta)和薄叶山矾(Symplocos anomala)等.竹林面积达7 000 hm2,毛竹年产量1 400余万根,被誉为“华夏毛竹第一镇”.土壤主要以黄棕壤和红黄壤为主,呈酸性[23].
在实验区的毛竹林设置固定3 块面积为20×50 m2的标准样地.模拟氮沉降采用施肥的方式进行,2013 年开始每年7-9 月,分2 次在处理样地内进行施肥(每次施肥量为全年的1/2,干洒),连续两年.施肥设2 个处理,分别是高氮(HN,100N kg·hm-2·a-1)和低氮(LN,50N kg·hm-2·a-1),用尿素作为氮源,不施肥作为对照(CK,0N kg·hm-2·a-1),样地土壤的基本情况如表1所示.

表1 实验毛竹林土壤特征
1.2 研究方法
1.2.1 多样性调查
毛竹林下植物多样性采用样方调查法,在不同处理样地中随机设置3 块面积为5 m×5 m 的小样方,对植物的种类[24]进行辨识并记录,主要包括灌木的树高(m),株数,3 cm处基径(cm)和冠幅(cm)用以分析灌木层的物种多样性;草本层(高度低于50 cm)记录草本植物的盖度和株数,用以分析草本层的物种多样性[25].
1.2.2 比叶面积测定
通过多样性指数计算,选取重要值靠前的优势灌木,对其进行比叶面积和养分特征分析.收集优势灌木叶片,按树种将叶片样品混合,挑选出无病虫害、叶形完整的成熟叶片,数量以30 枚为宜,可根据实际情况酌量减少,用湿毛巾擦干净后,使用叶面积仪测量叶面积并做好记录,将叶片装入信封后写上编号后放入烘箱,于105℃杀青后,在65℃烘干至恒重,称量样品干质量,计算比叶面积(specific leaf area,SLA)[26]:SLA=A*N/G.式中,A 为平均单叶面积,N 为叶片数,G 为N个叶片的干物质重量.
1.2.3 养分分析
将叶片处理成粉末状装入保鲜袋,用以测量全碳、全氮及全磷元素指标.用粉碎机将烘干至恒重的叶片磨碎,过筛,放入自封袋用于化学分析[27],使用EA 3000 元素分析仪(EuroVector,Italy)测定全C及全N,植物样品采用湿化消煮法[28]处理后使用FIAStar 5000流动注射仪(FOSS Analytical,Denmark)测定全P.
1.2.4 多样性数据处理
对样地的多样性数据进行整理,使用Excel 2007 对数据进行处理分析.计算植物的相对多度(RA)、相对频度(RF)、相对盖度(RC)、重要值(IV)和相关指数[29]:重要值(IV)=(相对密度+相对优势度+相对频度)/3;密度(D)=个体数目/样地面积;相对密度(RD)=100×(一个种的密度/所有种的密度);优势度(盖度C)=底面积(覆盖面积总值)/样地面积;相对优势度(RC)=100×(一个种的优势度/所有种的总优势度);频度(F)=包括该种的样地数/样地总数;相对频度(RF)=100×(一个种的频度/所有种的总频度).
物种丰富度指数:R=S,式中S为样地内所有物种数.
Margalef 指数:Dmg=(S-1)/lnN,式中S为样地内所有物种数,N 为样地中所有物种个体数之和.
均匀度指数采用Pielou指数:J=H'/lnS,式中S为样地内所有物种数.
2 结果与分析
2.1 毛竹林下植物物种结构特征
泾县蔡村毛竹林下植被以灌木、草本和蕨类植物为主.如表2 所示林下植被共计29 科35 属37 种,其中草本层13 科14 属14 种,含草质藤本1 科1 属1 种,蕨类植物3 科3 属3 种;灌木层16 科21属23种,含木质藤本2科2属2种,攀援灌木2 科2 属3 种.草本层中最具代表性的是禾本科,在不同处理样地中重要值均为最大;而灌木中以樟科、蔷薇科、茜草科、豆科、大戟科和百合科为主.

表2 实验毛竹林下植物物种组成及重要值
不同处理的样地中,同种植物的重要值不同.在不同处理的样地草本层中,均出现的植物有红根草、海金沙、荩草、爵床和黑足鳞毛蕨,其中荩草在不同处理样地中重要值均最大,且重要值随着氮添加增加而增大.而红根草的重要值随着氮添加而减小,除此以外,海金沙和爵床的重要值在低氮处理中呈上升趋势,在高氮处理中呈下降趋势;与之相反的是黑足鳞毛蕨,其重要值在低氮处理中下降,在高氮处理中上升.
灌木层植物有三种类型:攀援灌木、灌木和木质藤本.不同处理样地中均出现的有攀援灌木拔葜和木质藤本南蛇藤.氮处理样地中的菝葜和南蛇藤的重要值均高于空白对照样地.从表2可知,不同处理样地灌木层中均出现的13种植物包括满山红、檵木、六月雪、蓬蘽、野山楂、野蔷薇、拓树、野鸦椿、山胡椒、乌药、紫金牛、薄叶山矾和野桐,在不同处理中重要值不同,其中山胡椒、乌药、紫金牛和薄叶山矾的重要值在不同处理样地灌木重要值均较大.进一步分析可知,山胡椒和紫金牛的重要值在氮添加样地中均高于空白对照,而乌药的重要值与之相反,在氮添加样地中低于空白对照.薄叶山矾的重要值在高氮处理中呈下降趋势,而在低氮处理中呈上升趋势.
2.2 毛竹林下植物群落的主要指数特征
如表3所示,在灌木层中,对照样地的所有多样性指数的平均值和最大值均高于施肥处理的样地.在N 添加处理样地中,多样性指数中的物种丰富度和Margalef 指数,在HN 处理样地中的数值大于LN 处理样地;而Simpson指数、Shannon-Wiener指数和Pielou 指数在HN 处理样地中的数值小于LN处理样地.其中Simpson指数在空白对照样地显著高于N添加处理样地(P<0.05).

表3 不同施肥梯度灌木、草本的物种多样性
草本层中,各个多样指数变化规律不一致.与对照样地进行比较,物种丰富度指数数值在HN处理中呈上升趋势,在LN 处理中呈下降趋势.Margalef 指数在高氮与低氮处理中均呈下降趋势,Shannon-Wiener 指数则与之相反,在高氮与低氮处理中均呈上升趋势.而Simpson 指数和Pielou指数在高氮处理样地中呈现下降趋势,在低氮处理样地中呈现上升趋势.
不同处理样地中,灌木层的物种丰富度指数、Margalef 指数和Simpson 指数均大于草本层;灌木层的Shannon-Wiener 指数在HN 处理样地中低于草本层,但在LN 处理和对照样地中灌木层该指数大于草本层;Pielou指数在不同处理样地中均呈现草本层高于灌木层的特点.
2.3 林下灌木重要值及养分分析
通过重要值的计算,在林下植被中选取了紫金牛(Ardisia japonica)、山胡椒(Lindera glauca)、乌药(Lindera aggregata)、檵木(Loropetalum chinense)和薄叶山矾(Symplocos anomala)5 种常绿灌木的叶片为研究对象,测定了叶面积和碳氮磷养分元素并计算了比叶面积等指标.

表4 5种灌木养分分析
5 种不同的灌木的比叶面积在HN 和LN 处理样地中的数值均比对照样地中的数值大,即HN>LN>CK.比叶面积最大值出现在LN 处理的檵木,为17.30 m2·kg-1;最小值出现在CK 的薄叶山矾,为7.36 m2·kg-1.5种灌木的比叶面积氮含量为12.33~18.29 g·kg-1,最高的是低氮处理的紫金牛,最低的为对照样地的薄叶山矾.3 种不同处理梯度样地的碳含量均在369~525 g·kg-1.不同处理样地中P元素的变幅在0.37~1.13 g·kg-1.3种不同施肥梯度样地的灌木的碳氮比均值大于25,氮磷比均值大于14.对照样地中除乌药(14.69)和薄叶山矾(15.34)的氮磷比小于16,其余3种灌木叶片氮磷比均大于16,且乌药在HN(14.27)处理中氮磷比值与CK处理相比呈下降趋势.
对灌木层5 种灌木重要值、叶片的比叶面积和养分进行LSD 分析发现不同施肥梯度样地的灌木相关指标差异不显著,对于不同施肥梯度样地的分别进行相关性分析,结果见表5.
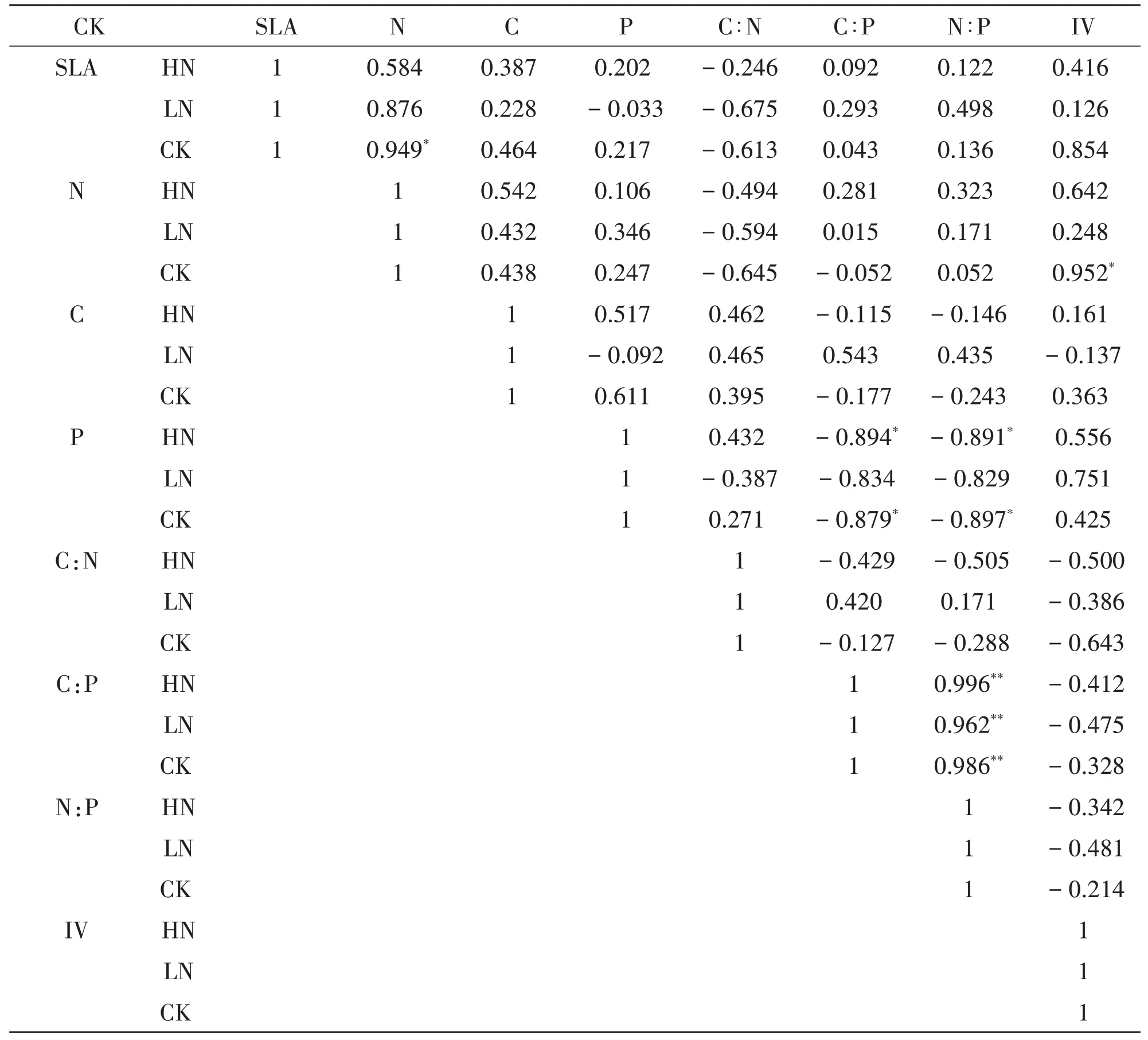
表5 灌木相关指标的相关性分析
从相关性分析可以看出对照样地中植物叶片氮含量(N)和重要值(IV)与比叶面积(SLA)呈正相关(P<0.05),高氮处理样地和空白对照样地中磷元素含量(P)均与碳磷比(C∶N)、氮磷比(N∶P)呈显著负相关(P<0.05).碳磷比(C∶P)与氮磷比(N∶P)在不同施肥梯度样地中均呈极显著正相关(P<0.01).
3 讨论与结论
毛竹林林下植被主要分为两层,草本层和灌木层,其中草本层中最具代表性的是禾本科植物,而灌木中以樟科、蔷薇科、茜草科、豆科、大戟科和百合科为主.而彭志等[30]在安徽的霍山县、广德县和黄山区三个地区的研究发现毛竹林下灌木主要以豆科、蔷薇科、大戟科、壳斗科、樟科为主,草本主要以禾本科、菊科、金星蕨科、百合科、唇形科、豆科等为主.对比发现草本层均有禾本科植物,而灌木层均有蔷薇科、豆科、大戟科和百合科植物,说明安徽地区竹林林下植被分布的植物种类较为相近.
就毛竹林下植物群落的主要指数特征而言,对照样地的灌木丰富度指数(R)、Margalef 指数(Dmg)、Simpson 指数(D)和Pielou 指数(J,均匀度指数)平均值均高于草本丰富度指数,这与彭志[30]等有关于安徽其他地区毛竹林林下植被多样性研究结论相一致,说明泾县毛竹林林下植被中灌木的种类较为丰富,且分布也较为均匀.对照样地中灌木的Simpson指数均高于草本,这与李艳霞[31]研究湖南省炎陵地区毛竹林植物多样性的结论相反.Simpson 指数(D)对外界环境扰动敏感,且受生境异质性和群落内物种个体数的影响较大,这也从侧面反映了安徽与湖南两个地区毛竹林的生境差异性较大.
除此以外,对照样地中所有多样性指数均高于其他施肥处理的样地.这与吴建平[32]等关于亚热带杉木人工林下植物多样性对氮沉降的响应研究结论一致,说明随着氮沉降的增加,植物多样性存在下降的趋势.
植物的氮磷比反映植物的生长状况[33].当植物叶片N∶P>16时,表示植物生长主要受P 元素的限制,而叶片N∶P<14 时,则主要受N 素限制,当N∶P 介于14 到16 之间时,植物生长受到N、P二者的共同限制[34].CK 样地中乌药的氮磷比介于14 到16 之间,说明乌药的生长受到氮、磷两种元素的限制,HN处理样地中的乌药亦是如此.但是在LN处理中乌药的氮磷比增大,说明LN处理中可能抑制了乌药对P 的吸收.李德军[10]在研究中指出氮沉降使土壤有效氮增加,导致土壤pH下降,趋于酸化.植物体内氮增加的同时,会抑制植物对磷元素的吸收,使氮磷比增加.由此可以看出LN 的添加导致磷元素吸收受限,氮磷比增大,但是进一步增加N 输入,会打破了P 元素限制,在HN处理中氮磷比下降.本实验中磷含量在不同处理中呈现不同变化规律,还需要进一步进行探究.
对照样地(CK)植物叶片氮含量(N)和比叶面积(SLA)呈正相关(P<0.05),这与崔珺[35]等在石台研究常绿阔叶林林下灌木叶片养分的研究结果一致,说明皖南地区灌木植被叶片N 的可利用性对植物的SLA 会产生一定的影响.前人研究指出,环境中的资源相对丰富,植物的比叶面积(SLA)更大,植物个体对资源的利用、光合和生长能力更强[36-37].本实验中氮添加样地中的5种灌木叶片的比叶面积均呈增大趋势,说明氮添加对植物的生长策略产生了影响.这5种灌木在氮添加环境有更好的资源利用能力,更能适应氮沉降量相对较多的环境.
综上所述,不论是从林下植被物种多样性角度还是从林下灌木的SLA 和化学计量的研究结果均说明氮添加对毛竹林林下植被多样性和植被的生长策略均产生了影响,但相关变化规律还需长期观测来探究.
