Application and prospects of high-throughput screening for in vitro neurogenesis
2022-07-05ShuYuanZhangJuanZhaoJunJunNiHuiLiZhenZhenQuanHongQing
Shu-Yuan Zhang,Juan Zhao,Jun-Jun Ni,Hui Li,Zhen-Zhen Quan,Hong Qing
Shu-Yuan Zhang,Jun-Jun Ni,Hui Li,Zhen-Zhen Quan,Hong Qing,Key Laboratory of Molecular Medicine and Biotherapy in the Ministry of Industry and Information Technology,Department of Biology,School of Life Science,Beijing Institute of Technology,Beijing 100081,China
Juan Zhao,Aerospace Medical Center,Aerospace Center Hospital,Beijing 100049,China
Abstract Over the past few decades,high-throughput screening(HTS)has made great contributions to new drug discovery.HTS technology is equipped with higher throughput,minimized platforms,more automated and computerized operating systems,more efficient and sensitive detection devices,and rapid data processing systems.At the same time,in vitro neurogenesis is gradually becoming important in establishing models to investigate the mechanisms of neural disease or developmental processes.However,challenges remain in generating more mature and functional neurons with specific subtypes and in establishing robust and standardized three-dimensional(3D)in vitro models with neural cells cultured in 3D matrices or organoids representing specific brain regions.Here,we review the applications of HTS technologies on in vitro neurogenesis,especially aiming at identifying the essential genes,chemical small molecules and adaptive microenvironments that hold great prospects for generating functional neurons or more reproductive and homogeneous 3D organoids.We also discuss the developmental tendency of HTS technology,e.g.,so-called next-generation screening,which utilizes 3D organoid-based screening combined with microfluidic devices to narrow the gap between in vitro models and in vivo situations both physiologically and pathologically.
Key Words: High-throughput screening;Stem cells;Neurogenesis;Cell differentiation;Three-dimensional cell culture;Cellular microenvironments
INTRODUCTION
High-throughput screening(HTS),also called large-scale cluster screening,first appeared in the 1980s and utilized microplates as a platform,an automated handling system as an operator,and a variety of highly sensitive detection instruments to screen out “hits” from thousands of drug candidates.As an integrated and multidisciplinary technology,HTS combines diverse fields,such as molecular biology,medicinal chemistry,mathematics,computer science and microelectronic technology.With its rapid,efficient,economic,microscale,highly automatic and computerized features,HTS technology has made great contributions to biomedical research,such as identifying new drug candidates for pharmacological research[1,2],enzyme engineering,including the directed evolution of enzymes[3],and genetic research[4].
Based on ligand-target interactions,HTS can be performed between various candidates and targets,including substrates and enzymes,inhibitors and enzymes,ligands and receptors,proteins and proteins,and DNA and proteins[5].According to thein vitroscreening models,HTS can be mainly divided into cell-free assays,also called biochemical assays,and cell-based assays.While cell-free assays dominate in the early stage of HTS,cell-based assays are gradually gaining an essential role because some cellular processes,such as transmembrane transport,cytotoxicity effects or other off-target effects,can be tested in cellular models,and some screening targets are difficult to extract and purify from cells[6].Significantly,in recent studies,screening targets have been extended from biochemical compounds such as enzymes,receptors,antibodies,nucleotides and living cells to tissues and even organoids to investigate protein–protein/DNA/RNA interactions,cell-protein interactions,cell–cell interactions and even protein-tissue interactions.Therefore,categories of testing candidates are also developing from biochemicals aiming at diverse targets such as enzymes and receptors in intracellular signaling pathways to microenvironments that are suitable for various cellular behaviors.Since the exploration of cell-extracellular matrix(ECM)-interactions is growing and three-dimensional(3D)cell culture technologies are developing,the HTS platform is evolving from two-dimensional(2D)to 3D.In previous research,hydrogel droplets and synthetic scaffolds could be attached to HTS platforms[7].
In addition to extending the variety of screening targets and candidates,researchers have also been working on improving the miniaturization,integration and automation of the screening platform to meet the increasing need for HTS applications in biomedical research.Specifically,the screening platform has developed from comprising microtiter plates with 96 wells to those with 384 wells and then to those with 1536 wells[8].To further elevate the screening efficiency,microarrays are utilized to promote integration by immobilizing protein or DNA targets on glass chips.Then,to separate each spot,save reagents and create various cellular microenvironments,combinations of microwells and micropillars are applied for HTS[9,10].
Furthermore,microfluidic-based microarrays also play an important role in HTS because of their higher efficiency,improved automation,controlled microenvironments,adjustable flow parameters,achievement of microscale reaction volumes and the capacity for single-cell analysis.Methods based on microfluidic systems can be divided into two groups: Droplet-based microfluidics and array-based microfluidics[11].Assay-based microfluidic devices have also been successfully utilized in HTS for drug screening[12,13],cell heterogeneity analysis[14],cell–cell interactions[15]and even cell–ECM interactions[15].
Compared with array-based microfluidic devices,droplet-based microfluidic devices are well suited for analyzing single-cell activities because biomolecules,particles or even single cells can be encapsulated in water-in-oil droplets that are emulsified when they are flowing through the dropletproducing devices to form the droplet library[16].Every droplet contains a barcode that represents the elements encapsulated.The barcodes usually include nucleic acid sequences,which are capable of large screens,and fluorescent tags,which are suitable for real-time reading[17].The droplets pooled in the library are then reinjected into the microfluidic device,usually merging with other cells or biomolecules to start the incubation,followed by a screening based on various characteristics,such as cell density[16]and fluorescence intensity[18].In addition,droplets can also be sorted according to the variety of readouts.The strategies applied in droplet sorting are based on fluorescence-activated cell sorting(FACS),which is a mechanical actuation,also called reverse cell sorting,accomplished with the assistance of peristaltic pumps and valves[19]as well as dielectrophoresis[20].
Over the years,the requirements of neurogenesis methods have grown with the increase in neurodegenerative diseases,andin vitroneurogenesis has been playing an important role in disease modeling,tissue engineering,drug screening and regenerative medicine[21-25].However,the ways to generate mature and functional neural cells with high efficiency and cell purity remain a problem.Here,we discuss how HTS technology promotes the progression ofin vitroneurogenesis in three sections,including screening out functional genes regulating neurogenesis,small molecules inducing neural lineage cells,and suitable microenvironments that facilitatein vitroneurogenesis.Furthermore,we review the applications of these obtained neural lineage cells using HTS technologies.Finally,with this review,we strengthen the connections between this promising and fast-developing technology andin vitroneurogenesis to raise awareness of generating more functional,mature and specific neural cells,as well as reproducible and standardized organoids with HTS technologies,for the sake of establishing robust neural developmental or disease models to better serve drug screening and regenerative medicine.
FABRICATION OF THE HTS PLATFORM
For the sake of designing various patterns for microarrays and microfluidic chips,depositing targets on the substrate surface is an essential step,which has been performed using a variety of methods,such as direct contact printing and noncontact printing,also known as ink-jet printing,photolithography,soft lithography,electron beam lithography,nanoimprint lithography,dip pen nanolithography,and laserguided direct writing[26].Direct contact printing can place desired biomolecules as ink from a stamp or a pin,linked to a high-precision robotic arm,to substrates with a reactive surface,which is usually accomplished by click reactions[27].Noncontact printing can eject samples in the form of droplets to specific positions mainly by piezoelectric and thermal printers[26].Photolithographic techniques can immobilize biomolecules on a substrate with photosensitive groups,for example,self-assembled monolayers such as alkanethiol[28],as linkers[29].Patterns on the microarrays can be designed according to the patterns on the masks,which could selectively activate the photosensitive groups with UV light irradiation[29],and then the solubility of the photoresists will change,leaving the substrate in the development step.For soft lithographic techniques,the word “soft” can describe elastomeric stamps or channels,which are made of commonly used poly(dimethylsiloxane)(PDMS)[30].PDMS stamps are utilized in microcontact printing,while channels are required in microfluidic channel flow patterning;these are the main methods used in soft lithographic techniques.The PDMS stamp can be fabricated using photolithographic techniques as the master is patterned with UV light and photoresists on the substrate,and then the liquid prepolymer is cast on the prepared master to form elastomeric stamps[30].After that,these stamps can pattern the substrates through microcontact printing using molecules that can interact with biomolecules and cells.
To further enhance the resolution to the nanoscale for higher throughput,electron beam photolithography and dip pen nanolithography are applied for direct protein patterning on microarrays[31-35].Nanoimprint lithography is also a nanostructure patterning technique that has been used to manipulate multiarchitectural chips with fields of topographies in nanometer dimensions to carry out highthroughput analysis for the screening of topographical structures that could promote stem cell differentiation[36,37].In addition to using biomolecules as targets,cells can also be directly patterned into substrates,although attaching them to biomolecules that have been positioned to substrates is another matter.This technique is called laser-guided direct writing,in which the laser,focused by the lens,propels single cells with optical forces toward the substrate to form cell clusters[38].This technique has been applied in tissue engineering through the reconstruction of tissues by micropatterning cells on soft matrices such as collagen or Matrigel to build cell–cell interactions that resemble those in the native microenvironment[39].
CURRENT IN VITRO NEUROGENESIS METHODS
For neurobiological research,in vitroneurogenesis plays a significant role in conducting drug screening,establishing models for investigating mechanisms of neural development or diseases,and deepening research on regenerative medicine for cell therapy and tissue engineering[22,23,40].Consequently,exploring more efficient methods forin vitroneurogenesis,including obtaining pure and functional neurons,building neural circuits,and forming neural tissue and even cerebral organoids,is of vital importance.To date,various methods have been used to manipulatein vitroneurogenesis(Figure 1).Embryonic stem cells(ESCs),pluripotent stem cells(PSCs)and mesenchymal stem cells(MSCs)have been induced to differentiate into functional neuronal cells through growth factors such as epidermal growth factor(EGF),basic fibroblast growth factor(bFGF),and insulin-like growth factor 1(IGF-1)as well as neurotrophic factors such as neurotrophin-3(NT-3),brain-derived neurotrophic factor(BDNF),and glial cell line-derived neurotrophic factor(GDNF)(Supplementary Table 1)[41-43].Sometimes,to obtain specific subtype neurons,key factors participating in subtype neuronal development are added;for example,sonic hedgehog(Shh)and fibroblast growth factor 8(FGF8)are reported to be essential in the induction of dopaminergic neurons[44,45].
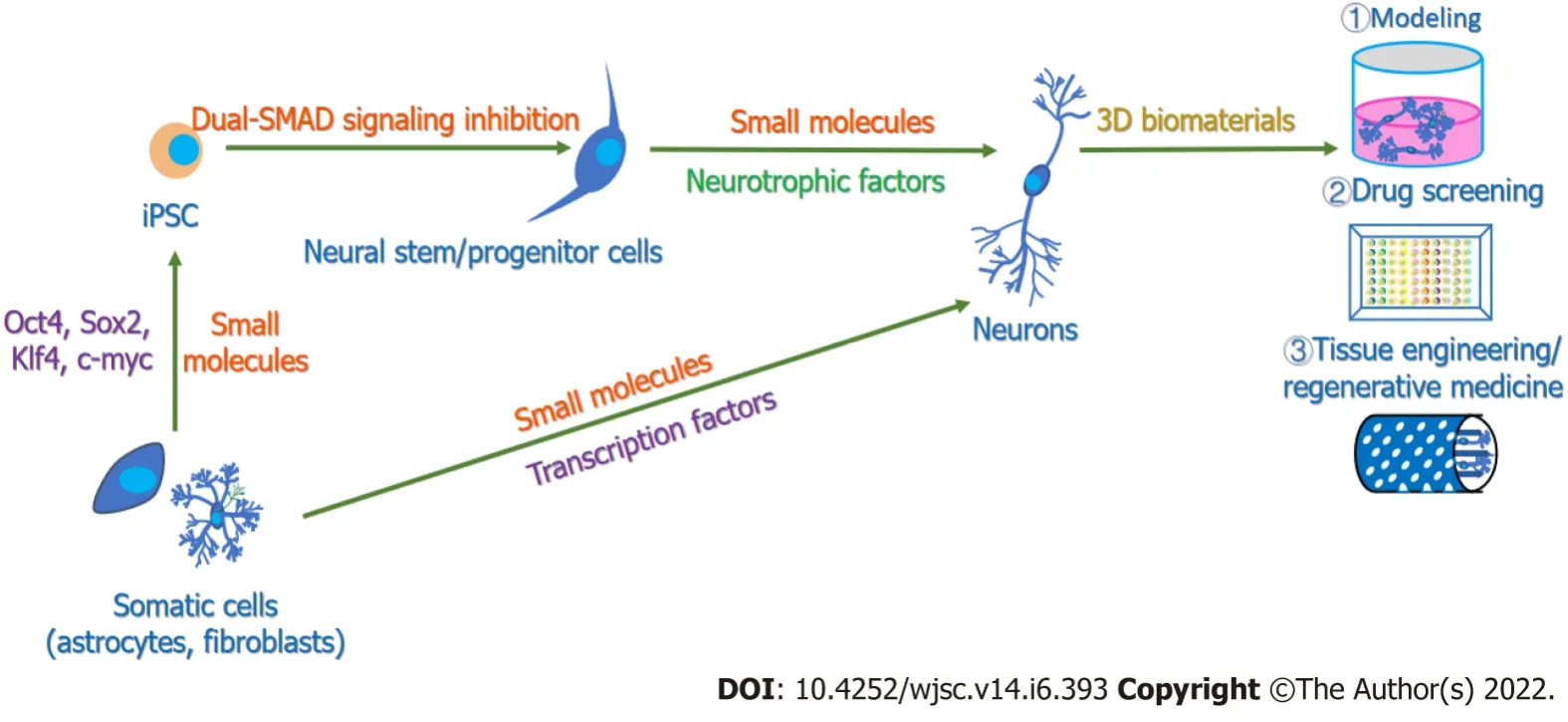
Figure 1 Main protocols and applications of in vitro neurogenesis.Neurons can be generated in vitro through the differentiation of induced pluripotent stem cells or the transdifferentiation of somatic cells with the aid of transcription factors,growth factors or small molecules[51,131].With the combination of 3D biomaterials,induced neurons can be applied in various fields,including modeling,drug screening,neural tissue engineering and regenerative medicine[213,232,276].iPSCs: Induced pluripotent stem cells.
Furthermore,it has been found that the forced expression of distinct neurogenesis fate determinants,as well as the help of growth factors,can support the differentiation of ESCs and PSCs toward neural lineage cells(Supplementary Table 1)[46-49].In fact,since induced PSCs(iPSCs)were developed by the integration of four transcription factors[50],neural transcription factors have been an efficient tool to reprogram fibroblasts[51,52],astrocytes[53,54]and other cell lines[55]to neuronsin vitro.Recently,with the assistance of HTS,researchers have successfully explored some small molecules and their combinations to promote neural differentiation[56,57]and cell reprogramming[58-61]to obtain functional neurons(Supplementary Table 1).In these ways,functional and pure neurons with different neural subtypes,including dopaminergic neurons,cholinergic neurons,GABAergic neurons and glutaminergic neurons,have been generated.
Over the years,3D culture systems have been developing rapidly,as they can provide cells with mechanical support,external stimuli and cell–ECM interactions to better mimic the architecture and functions of thein vivomicroenvironment in living tissues and organs compared with conventional 2D cultures[62].For many years,adding 3D matrices has become an attractive choice for the culture of neural cells and the generation of neural tissues or even neurospheresin vitro[63-65].For cell culture,the three most commonly used models include the “on-gel model”,the “sandwich gel model” and the “ingel model”.For even more complex 3D culture models,a study has cocultured predifferentiated human PSC(hPSC)-derived astrocytes with hPSC-derived,Ngn2-induced neurons on Matrigel-coated glass chips and observed more mature morphology of astrocytes,followed by the coculture of astrospheres and neurospheres on polytetrafluoroethylene membranes as organotypic-like cultures,and the astrocytes improved morphological complexity with fine leaflet-like structures[66].
There have also been other 3D culture systems containing spinner flasks and rotary bioreactors that can prevent cell attachment to the surface and aid in generating 3D spheroids[67],such as neural organoids.Over the past decade,organoid technologies have become a promising method to bridge the gap between cellular models and animal models and to better recapitulate the complexity of the cytoarchitecture at the tissue/organ level.The technology relies on the self-organization,self-renewal and differentiation capacity of ESCs or iPSCs and their potential to form cells from three germinal layers[68].The protocol for generating cerebral organoids from hPSCs mainly includes the generation of embryonic bodies,the neural induction period,the neural differentiation period embedded in Matrigel,and the procedure for brain tissue growth and expansion in spinning bioreactors to provide enough oxygen[69,70].Therefore,with the supplementation of 3D matrices and specific biomolecules,including growth factors and small molecules that precisely control the signaling pathways regulating neural lineage induction and neural differentiation,as well as devices providing sufficient oxygen for tissue growth,cerebral organoids containing discrete brain regions[69]and even organoids with specific brain regions,such as forebrain organoids[71,72],midbrain organoids[73,74],thalamic organoids[75]and cortical organoids[76],can also be generated.Specifically,to imitate morphogen concentration gradientsin vivo,an inducible Shh-expressing hPSC line was constructed and embedded in one pore of developing organoids to create a Shh signaling gradient[72].Treatment with LDN193189,SB431542 and XAV939 inhibited bone morphogenetic protein(BMP),transforming growth factor(TGF)-β,and Wnt signaling,respectively,to induce organoids toward forebrain identity[72,77].Midbrain organoids were also generated based on the mechanism of regional patterning,as the addition of FGF8 and activation of Shh signaling have been proven to be significant elements in inducing midbrain dopaminergic identity[73,74,77,78].Cortical organoids are induced through dual-SMAD signaling suppression following the induction of cortical identity patternsin vivo[76,77].
However,the self-organization ability of PSCs usually leads to an unpredictable arrangement of the internal structure of the organoids,which increases the uncertainty of their applications inin vitromodeling.Therefore,to improve the reproducibility between batches,a feasible method is to control the initial culture conditions,such as the starting cell types,cellular density,3D matrices,and size and geometry of aggregates,because minimal deviations from the initial conditions will generally result in batch-to-batch controlled organoids[79].To achieve this,trials have been performed to engineer 3D matrices and explore more suitable scaffolds,such as poly(lactic-co-glycolic acid)(PLGA)microfilaments,for guiding self-organization[71].
APPLICATIONS OF HTSS TO NEUROGENESIS
Identification of functional genes regulating neurogenesis
During neurogenesis,genes correlated with neural development are expressed in spatial and temporal order[80].In previous studies,key genes have been identified[81]and have recently been the focus of further trials seeking to generate functional neural cells[48].Significantly,basic helix loop helix transcription factors play important roles in neural specification and differentiation[82],and the forced expression of transcription factors has been manipulated to convert ESCs,iPSCs or nonpluripotent somatic cells into functional neurons even with specific subtypes,including midbrain dopaminergic neurons,spinal motor neurons,and forebrain GABAergic interneurons[21,51,83-86].In addition to transcription factors,miRNAs are also effective tools to induce neural cell types;for example,Yooet al[87]reported that miR-9/9* and miR-124,together with NeuroD2,could convert human fibroblasts into functional neurons by regulating SWI/SNF-like BAF chromatin-remodeling complexes.
For decades,functional gene identification has been carried out based on forward genetic approaches,mostly through whole-genome mutagenesis screening.Chemical mutagens,such as ethylmethanesulfonate and ethylnitrosourea,as well as polymerase chain reaction-based gene deletion strategies,are commonly applied to induce mutations,and phenotypes are evaluated to screen out functional genes[88-91].However,for neurogenetic research,this method usually requires model organisms,including transgenic mice,Drosophila or zebrafish,which makes it time-consuming and difficult to manipulate screening on high-throughput platforms.Reverse genetic approaches are also becoming popular strategies to perform HTS to identify functional genes controlling various cellular behaviors.Since the discovery of RNA interference(RNAi),RNAi-based HTS has shown superiority in the identification of functional genes,the dissection of signal transduction pathways,and target exploration for drug development[92,93].Compared with traditional forward genetic methods,RNAi screening fits more to cell-based screening,which is more suitable for conducting experiments at the cellular and molecular levels and is easier to utilize on HTS devices.Due to its advantages,this technique has been widely applied to identify genes that regulate neural development.Koizumiet al[94]injected double-stranded RNA into the preblastoderm embryos of Drosophila and found 22 genes that influence embryonic nervous system development,seven of which had unknown functions,nine of which had known functions,and the rest of which had known nervous system development phenotypes,such as dmt expressing a nuclear localization motif for peripheral nervous system development.Güneşet al[95]performed a selection-based screening and transduced CD34+hematopoietic stem and progenitor cells and neural stem cells(NSCs)with shRNA.After next-generation sequencing and the comparison of shRNA representation in two cell types,they determined SMARCA4 to be the stemness regulator that controls NSC self-renewal by upregulating RE-1,a silencing transcription factor,and downregulating BAF53,suggesting its function in the repression of cell differentiation toward the neural lineage[95].Other RNAi screening studies have also identified genes that play roles in neural outgrowth,axonal regeneration and neural cell death[88,96,97].
Over the years,with the discovery of the clustered regularly interspaced short palindromic repeats(CRISPR)system in prokaryotic organisms,CRISPR/Cas9 has opened new avenues toward HTS.The method of CRISPR-based HTS can be divided into arrayed/plate screens and pooled/barcode screens[4].In arrayed screen format,the process is conducted in multiwell plates,and sgRNAs are individually delivered through viral transduction or transfection,followed by screen readout through highthroughput imaging,while for pooled screens,sgRNAs are synthesized and delivered as a pool,followed by viability screening through next-generation sequencing to evaluate the difference in the abundance of sgRNAs between samples for the identification of target genes or phenotypic screening,such as through FACS[4].Compared with RNAi screening,CRISPR/Cas9-based screening has fewer off-target effects and is capable of investigating nontranscribed spacers and noncoding RNA,as RNAi can suppress gene expression only at the posttranscriptional level,whereas CRISPR/Cas9 can invalidate target genes at the genetic level[98].Thus,the applications of CRISPR/Cas9 screening have grown in recent years,and genes related to bacterial toxicity[99],DNA mismatch repair pathways[100],new drug targets[101],cell viability and proliferation[102]have been identified in human cancer cells.
Significantly,CRISPR activation(CRISPR/a)and CRISPR interference(CRISPR/i)also play important roles in HTS in the identification of functional genes.These methods have been reported to serve as an effective tool in developmental biology studies.For instance,Gengaet al[103]combined CRISPR/i screening and single-cell RNA-seq and identified FOXA2,the transcription factor that plays a significant role in endoderm development,as inhibition of FOXA2 impaired differentiation toward the foregut endoderm and the subsequent hepatic endoderm.In a neural differentiation study,Liuet al[104]utilized CRISPR/a screening through the establishment of a sgRNA library consisting of 55561 sgRNAs targeting all computationally predicted transcription factors and other DNA-binding factors and identified various transcription factors that could promote neural differentiation by sorting Tubb3-hCD8+cells.They also studied the interactions between these hits through a combination of two sgRNAs and found that Ngn1,along with Brn2,Ezh2 or Foxo1,significantly improved the conversion efficiency of mouse embryonic fibroblasts(MEFs)into neurons[104].The investigation of one of the hits,Ezh2,showed the downregulation of endodermal-and mesodermal-related genes in Ezh2-induced neurons,which indicates the possible mechanisms of enhanced neural differentiation of Ezh2[104].
Given that HTS possesses the ability to identify functional genes,particularly transcription factors related to neural development,future directions could still focus on the genetic networks of neurogenesis,especially the function of noncoding sequences,such as noncoding RNAs.For example,Zhuet al[105]designed a paired guide RNA CRISPR/Cas9 Library to delete approximately 700 long noncoding RNAs(lncRNAs)in the human liver cancer cell line Huh7.5OC.From a genome-scale lncRNA deletion screen,they found 9 lncRNAs positively or negatively correlated with the proliferation and survival of cancer cell lines[105].Furthermore,CRISPR/a and CRISPR/i have also been proven to be useful in lncRNA screening.Liuet al[106]performed a genome-wide CRISPR/i screen with dCas9-KRAB targeting 16401 lncRNA loci in 6 transformed cell lines and an iPSC cell line,and the results showed that 499 lncRNAs participated in cell growth,such as LINC00263,the knockdown of which downregulated the proliferation of the U87 cell line and upregulated endoplasmic reticulum stress-and apoptosis-related genes.A CRISPR/a screen was also applied to identify lncRNAs related to the drugresistance pathway in cancer cell lines[107].In addition,CRISPR screening has been reported to discover functional miRNAs.Panganibanet al[108]conducted a genome-wide CRISPR/Cas9 screen and recently demonstrated that knockout of miR-124-3 led to upregulation of C/EBP homologous protein 10,the transcription factor associated with ER stress-mediated apoptosis,by regulating the IRE1 branch of the ER stress pathway.Moreover,screening out noncoding genes that serve as endogenous regulatory elements could help to further deepen our understanding of gene expression regulation.Klannet al[109]performed CRISPR/Cas9-based epigenomic regulatory element screening through dCas9-KRAB and dCas9-p300 to repress or activate the activity of the DNase I hypersensitive site and identified previously uncharacterized regulatory elements controlling the expression of b-globin and HER2.Baumannet al[110]strongly induced master transcription factor Sox1,as well as a neuroepithelial marker,through dCas9-VP64 to target the promoter of Sox1 and restored the neuronal differentiation potential of NPCs.Then,they transfected dCas9-Tet1 into NPCs that stably expressed dCas9-VP64 after transducing gRNA and found that the neuronal differentiation potency was increased with dCas9-Tet1 decreased DNA methylation levels around the transcription start of Sox1[110].Thus,it is helpful to further understand the intrinsic interaction between activation of transcription factors and the regulation of epigenetic editing and even chromatin modification in neural development.Consequently,HTS could be a leading method to identify key genes and pathways,including transcription factors and noncoding regions related to neural development,which would provide novel targets forin vitroneurogenesis.
Identification of small molecules for neural lineage induction
Although the overexpression of transcription factors in initial cells has made great progress in the induction of functional terminal differentiated cells,it could lead to safety problems such as tumorigenesis when the viral vectors integrate into the genomes of receptor cells[111].Compared with the forced expression of transcription factors,utilizing cell-permeable chemical small molecules is safer,more cost-effective,less time-consuming,and easier to standardize.Thus,developing small molecules that could replace the effect of transcription factor overexpression has great prospects.This assumption first came into reality when Houet al[112]identified a combination of seven small molecules that could reprogram MEFs to ESC-like PSCs.Before that,when many studies focused on the identification of small molecules that are capable of replacing defined transcription factors,HTS technology made huge contributions to identifying those specific small molecules.
For HTS technology,cell-free assays have been of great support for the identification of small molecules based on their effects on activating or blocking signaling pathways that facilitate or inhibit neural differentiation,which usually act as agonists or antagonists of kinases belonging to those signaling pathways.For example,SB431542 is an inhibitor of the ALK5/TGF-β1 receptor,which was identified by a flashplate-based assay with the immobilization of GST-tagged Smad3,and GST-tagged ALK5 was used as a kinase[113].Except for using the interaction between biomolecules,especially between candidates and kinases or receptors affiliated with specific signaling pathways,cell-based methods are more commonly applied in screening compounds that could replace essential transcription factors,including in the period when the chemical induction method was the only method available for exploration.Takahashi and Yamanaka[50]screened out iPSCs by integrating a bgeo cassette,a fusion of the β-galactosidase and neomycin resistance genes,into Fbx15,a downstream gene of Oct4,to conduct iPSC screening through drug resistance to G418.However,iPSCs isolated in this manner were different from ESCs in their gene expression patterns,and screening was performed based on the activation of endogenous Nanog or Oct4 with a drug-resistance marker or the green fluorescent protein(GFP)reporter gene[114-116].Recently,optical screening methods using fluorescent proteins,luciferase or the lacZ gene were proven to be effective in cell-based screening.To screen small molecules that could replace Sox2 in reprogramming,an Oct4-GFP transgenic reporter was used to provide Oct4-GFP+colony numbers so that the reprogramming efficiency could be represented after retroviral transduction of MEFs with Oct4,Klf4 and c-Myc[117].Then,from screening 200 small molecules,it was found that Repsox could substitute Sox2 even without c-Myc or the histone deacetylase inhibitor valproic acid(VPA),which could greatly improve reprogramming efficiency[118]through the inhibition of the TGF-β signaling pathway[117].In addition,the G9a histone methyltransferase inhibitor BIX-01294 was also demonstrated to substitute Sox2 in the presence of Oct4 and Klf4(c-Myc is dispensable when generating iPSCs from mouse and human fibroblasts)[119,120].Other transcription factors,including Klf4,could also be replaced during reprogramming by VPA[121]or the GSK-3β or CDK inhibitor kenpaullone[122],and Oct4 could be replaced by the inhibitor of the ALK5/TGF-β1 receptor,SB431542 or Repsox[123].
In addition,HTS has been generally used to screen small molecules that could generate functional neurons to eliminate the ectopic expression of transcription factors.Liet al[59]screened approximately 5000 small molecules using Ascl1-infected mouse fibroblasts and found that forskolin,SB4315342,ISX9 and CHIR99021 could improve the number of TauEGFP-/Tuj1-positive neural cells.Subsequently,IBET151 was screened out from approximately 1500 candidates in the presence of the former four small molecules and in the absence of the transcription factor Ascl1[59].In subsequent tests,it was found that ISX9 was capable of activating the master neural genes NeuroD1 and Ngn2[59].In addition,different combinations of various small molecules can also be screened to obtain more efficient cocktails for neural differentiation.Chamberset al[124]screened approximately 400 different combinations according to the day the compounds were added,and they confirmed that CHIR99021,DAPT and SU5402,which were added at day two of differentiation,along with SB431542 and LDN193189,could direct the differentiation of human NPC(hNPC)into nociceptive sensory neurons.They evaluated the decrease in PAX6+ cells and the improvement of TUJ1+ cells as the screening phenotype[124].In another study,Zhanget al[61]performed a screen containing hundreds of combinations among 20 small compounds with different concentrations in a stepwise protocol and identified a cocktail including LDN193189,SB431542,TTNPB,thiazovivin,CHIR99021,VPA,DAPT,SAG,and purmorphamine that could convert human astrocytes into functional neurons.
To date,small molecules have been widely utilized in inducing neural cell lineages from PSCs or somatic cells,as a number of experiments have successfully generated functional neurons from ESCs,fibroblasts or astrocytes by chemical cocktails(Supplementary Table 2)[56,58-61,125,126].Among the chemical compounds,SB431542 and LDN193189 have been widely applied to induce neuroectodermal cell lines from PSCs since the inhibition of dual-SMAD signaling was proven to be capable of converting human ESCs(hESCs)to PAX6+ neuroepithelial cells by the SB431542/Noggin protocol[127].LDN193189 is an inhibitor of ALK2/ALK3 that was used to replace noggin,a soluble BMP antagonist.Dorsomorphin,also known as Compound C,was able to inhibit the BMP type 1 receptors ALK-2/ALK3/ALK6 and subsequently repress the phosphorylation of Smad1/5/8[128].However,dorsomorphin resulted in moderate inhibition and unstable metabolism;thus,LDN193189 was recently developed and exhibited a highly decreased IC50[129].In addition,CHIR99021,also named CT99021,is an inhibitor of GSK3β and can activate Wnt signaling,which has been demonstrated to play an important role in maintaining neural stem/progenitor cell proliferation and differentiation[130].Nevertheless,CHIR99021 has been reported to induce the neural crest lineage through activation of the Wnt signaling pathway[124],the inhibition of which by XAV939 could help to generate cortical neurons[131].Of note,CHIR99021 was added in the final differentiation step,as it can function in the promotion of axonal outgrowth and synapse formation[131].Significantly,Kirkebyet al[57]tested CHIR99021 in a dose-dependent manner and found that hESCs differentiated into neural progenitors with all regions from the telencephalon to the posterior hindbrain along the rostrocaudal axis following increasing concentrations of CHIR99021.Except for the rostrocaudal axis,dorsoventral axis patterning could also be induced by small molecules,as ventralization can be regulated by Shh,and dorsalization can be controlled by the Wnt canonical pathway and the BMP pathway[77].Purmorphamine is a Smoothened receptor agonist that can activate the Shh signaling pathway and has a similar effect as another small molecule called SAG,which is a potent Smoothened receptor agonist.Thus,neural subtypes could be enriched through the coordination of Shh signaling,Wnt canonical signaling and BMP signaling[132].Forskolin is also a commonly used small molecule that functions as a diterpene adenylate cyclase activator,and the addition of forskolin could increase the level of intracellular cyclic AMP(cAMP).Significantly,the activation of cAMP/PKA-cAMP-responsive element binding(CREB)signaling,for example,by treatment with dibutyryl-cAMP,can phosphorylate CREB protein,which is an essential transcription factor regulating many target genes related to the survival,proliferation and differentiation of neurogenic cells,such as Bcl-2,BDNF,tyrosine hydroxylase and somatostatin[133].Furthermore,SU5402 and PD0325901 are inhibitors of FGFR1 and mitogen-activated extracellular activated signal-regulated kinase(MEK),respectively,which all serve as inhibitors of FGF/MEK/ERK signaling.DAPT,a g-secretase inhibitor,is likewise a small molecule commonly applied as a Notch signaling inhibitor.There have been a number of studies using the inhibition of FGF and Notch signaling to suppress cell proliferation and thus lead to differentiation[124,126,131].
To date,with the support of the HTS method,many types of small molecules have been developed to convert pluripotent cells such as ESCs and NPCs or nonneural somatic cells such as fibroblasts into neural lineage cells.However,there is still great demand to generate mature neurons with specific neural subtypes and positional cues of different brain regions.To meet this requirement,more selective chemical compounds with optimum concentrations and combinations with different addition orders are desired.Mauryet al[134]utilized the automated 384-well plate format to treat hNPCs with various concentrations,durations,and combinations of small molecules and directed NPCs to spinal motor neurons and cranial motor neurons with specific regional identities.Additionally,developing diverse types of small molecules that regulate gene transcription through different mechanisms could be another perspective to expand the collection of small molecules for neural lineage conversion.In addition to small molecules that work as signaling pathway modulators,other chemical compounds that can repress epigenetic-related enzymes,such as histone-modifying enzymes,DNA methylationassociated enzymes,and modulate nuclear receptors remain to be further explored[135].For example,VPA,an inhibitor of histone deacetylase;RG108,an inhibitor of DNA methyltransferases;and TTNPB,a retinoic acid(RA)analog and a nuclear receptor RAR agonist,are applied in the conversion of neural cells through transdifferentiation methods[58,61,126,136].Therefore,further investigations of epigenetic mechanisms and the orchestrated signaling processes underlying neural lineage specification are required to develop targeted small molecules.
Screening for more suitable microenvironments for in vitro neurogenesis
The cell microenvironment consists of ECM,soluble molecules such as cytokines and hormones,and interactions with adjacent cells.To better facilitate neurogenesisin vitro,screening and reconstituting suitable microenvironments similar to thosein vivoare important.However,traditional culture systems cannot be used to assess various microenvironmental factors at the same time.With the HTS platform,it is much more convenient and efficient to evaluate various parameters,including the type,topography,and stiffness of 3D materials and the types and concentration gradients of soluble biomolecules on a highly integrated chip.
Screening for 3D scaffolds:ECM molecules,mainly containing laminin,fibronectin,collagen Ⅳ,entactin,elastin,heparan sulfate proteoglycans,hyaluronan,chondroitin sulfate proteoglycans and tenascin-R in the CNS[137,138],are essential components of the microenvironment.In the natural matrices above,the widely applied 3D matrices are collagens[63,64,139,140],hyaluronan[141],and another commonly applied biomaterial called Matrigel[22,142,143]due to their superior contributions to thein vitroneural proliferation,differentiation and outgrowth of NSCs,NPCs or ESCs,some of which also build neural circuits and recapitulate CNS neural development.
Since these natural matrices are largely extracted from animals or cultured cells,it is difficult to control the biochemical and mechanical cues of batches[62].Thus,to increase the reproducibility and reliability in further applications,synthetic scaffolds such as synthetic polymer hydrogels,which mainly contain self-assembling peptide hydrogels[144,145],poly(ethylene glycol)(PEG)[146-148],poly(lactic acid)[149,150],PLGA[151,152]and electrically conductive polymers including poly(pyrrole)[153-155],and carbon nanotubes[156,157]have become attractive tools for 3Din vitroneurogenesis(Supplementary Table 3).Although there are various kinds and different combinations of biomaterials to utilize[158],it would be helpful if we make the best use of HTS techniques to make comparisons of which kinds or combinations of biomaterials will be the priority for cell culture.Therefore,in future studies,it will still be quite important to explore recipes of biomaterials that work as effectively as possible for neurogenesisin vitrothrough 3D cell culture.
Microarrays based on glass slides on 2D platforms have been typically used for biomaterial screening,usually including polymer microarrays for screening synthetic polymer scaffolds and ECM or tissue microarrays for screening naturally sourced matrices.As mentioned before,biomaterials are patterned on slides through contact printing,injection printing and photolithography[159].For polymer microarray screening,Andersonet al[160]fabricated a nanoliter-scale polymer array on which there were 576 different acrylate-based polymers in triplicate,synthesized by diverse combinations of 25 kinds of monomers through a light-activated radical initiator and UV light,attached to a poly(hydroxyethyl methacrylate)-coated slide.This microarray was used to evaluate polymer-based biomaterials for hESC attachment,spreading,proliferation and differentiation into cytokeratin-positive cells[160].Using a similar method,this team subsequently constructed another polymer microarray with 1152 different combinations of 24 polymers mainly containing different forms of PLGA in triplicate[161].They seeded human MSCs(hMSCs)and found that biomaterials containing PLGA-PEG inhibited cell attachment and spreading,while PLGA containing L-lactide could relieve this inhibition[161].Recently,they carried out a screening with 22 acrylate monomers copolymerized in different combinations to generate 1488 arrays and tested their effects on hESC self-renewal[162].
In addition to polymer-based arrays,ECM microarrays are also commonly utilized for the investigation and dissection of functional elements for regulating cell behavior.Nakajimaet al[163]displayed ECM-based biomaterials,including collagen I,collagen IV,fibronectin and laminin,as well as artificial biomaterials containing acidic gelatins,basic gelatins,ProNectinTMF plus and ProNectinTM L,poly(Llysine),and poly(ethyleneimine)with weight-averaged molecular weights of 800,10000,25000,and 750000 on gold-coated glass plates.The results showed that fibronectin,laminin,Pro-F,Pro-L and PEI-0.8 could support NSC adhesion,while collagen and gelatins had no effect on NSC adhesion[163].The probable reason for these results could be that NSCs adhere fibronectin and laminin through b1 integrin,and an electrostatic interaction might have promoted NSC’s adhesion to PEI-0.8[163].Ahmedet al[164]screened 190 combinations of 19 ECM proteins that were selected according to their expression in the ventral midbrain during dopaminergic neurogenesis and identified that Sparc,Sparc-like(Sparcl1)and Nell2 could synergistically increase the number of TH+neurons differentiated from long-term neuroepithelial stem cells.
In addition,tissue matrix-based microarrays were fabricated by removing soluble components and mechanically fragmented matrices from 11 different porcine tissues and organs,which could preserve the natural diversity and complexity of biomaterials[165].In this way,studies could focus on naturally sourced ECM components from various tissues and organs and analyze the tissue/organ-specific differences that subsequently lead to cell lineage specification.While retaining the complexity of ECM proteins,disassembling functional domains could also be an effective approach.Linet al[166]conducted a peptide microarray and seeded normal murine mammary gland cells and demonstrated that the peptides LTGKNFPMFHRN and MHRMPSFLPTTL could induce epithelial-to-mesenchymal transition and decrease E-cadherin levels.
Screening for surface topography and morphology:In addition to the type of biomaterial,surface topography and morphology can also support neural induction.Studies have been performed to investigate the impact that continuous,discontinuous and random topographies that biomaterials have on the guidance and outgrowth of axons and dendrites[167].In particular,continuous topographies can impact the orientation and shape of NSCs through the regulation of cytoskeleton rearrangement and nucleus elongation,while discontinuous isotropic topographies are reported to induce NSCs to the glial lineage[168].Thus,to explore more suitable biomaterials expected to support neural growth,precise surface topologies were screened to search for the topologies that promote axon and dendritic growth.Large-scale screening showed that anisotropic grating patterns could best facilitate axon growth,while dendrites showed almost no sensitivity to surface topologies[169].Furthermore,matrix stiffness also plays an unignorable role in neural development,as distinct subtypes of neurons exhibit different neurite outgrowth rates when cultured in conditions of varying elasticity[170].To investigate the mechanical properties suitable for controlling specific cellular behavior,HTS has been utilized;for instance,Kumachevet al[171]constructed a droplet-based screening platform by encapsulating mESCs into agarose microgels with different elastic moduli.Kouroukliset al[172]arrayed various combinations of five ECM proteins on a poly(acrylamide)hydrogel substrate with three different elastic moduli to assess the effect of substrate stiffness on the differentiation of bipotential mouse embryonic liver progenitor cells.
Screening for combinations of growth factors:Soluble bioactive molecules such as growth factors,including bFGF and EGF;members of the neurotrophin family,including BDNF and GDNF;and members of the TGF family are important parts of the microenvironment during neurogenesis,regulating neural proliferation and differentiation[173-175].The HTS method could be used to evaluate the best candidate or the best combination to promotein vitroneurogenesis.Konagayaet al[176]immobilized five growth factors,including bFGF,EGF,IGF-1,BDNF,and ciliary neurotrophic factor(CNTF),on a chip and displayed them either as a single component or as the combination of any of two factors to explore their function on NSCs.They found that either bFGF or EGF alone could facilitate the proliferation of NSCs,and that the combination of these two factors showed a synergistic effect.Both IGF-1 and BDNF could facilitate NSC differentiation toward the neural lineage,but CNTF promoted glial lineage differentiation[176].Nakajimaet al[163]used a cell-based assay to coimmobilize growth factors and natural or synthetic matrices,and they found that EGF promoted the maintenance of NSCs and that two nerve growth factor(NGF)family members,NGF and NT-3,could facilitate NSCs toward neuronal differentiation.In addition to for cell-based microarrays requiring immobilization of biomolecules,Muckomet al[177]applied a high-throughput microculture system consisting of complementary micropillars and microwells that could hold 532 independent microenvironments for cell culture.They seeded adult rodent NSCs and provided 6 soluble factors,BMP4,TGF-β,FGF-2,shh,Wnt-3a and Ephin-B2,and evaluated the extent to which their individual signals and double,tertiary and quaternary signal combinations could influence neural differentiation[177].Their results indicated that Wnt-3a and Ephin-B2 synergistically facilitated neural differentiation and maturation,while TGF-β,FGF-2 and Wnt-3a affected NSC proliferation and differentiation antagonistically[177].
Screening for 3D microenvironments:Beyond exploring single variants such as the abovementioned ECM proteins,surface topography,matrix stiffness and soluble factors,it could be more effective to combine various elements together and screen the whole microenvironment(Supplementary Table 4).A typically used approach is to premix ECM proteins and soluble signaling molecules in different combinations within multiwell plates,such as 384-well plates,and then to codispense the mixtures on substrate slides of the microarray.Later,cells are seeded on each spot.Linet al[178]designed a microarray capable of screening microenvironments,including substrate stiffness,ECM matrices,various growth factors and cytokines.The elastic modulus could be adjusted by altering the base/cure ratio of PDMS to mimic hard tissues such as cartilage,cornea,and arterial walls,while regulating the acrylamide/bisacrylamide ratio of PA could mimic soft tissues,including the brain,liver,and prostate[178].Soenet al[179]also constructed a microarray for screening out microenvironments from 44 combinations of ECM proteins and signaling factors that promote the neural differentiation and specification of primary human NSCs(hNSCs).Moreover,Brafmanet al[180]designed a 3D microarray screening method called arrayed cellular microenvironments,which could hold 8000 spots to screen microenvironments containing ECM proteins,growth factors and small molecules for evaluating cell attachment,growth and proliferation of hPSCs.To avoid interference between each spot on microarrays and to make the microculture system more suitable for culturing nonadherent cells,Gobaaet al[181]constructed arrays of PEG hydrogel microwells that could hold 2016 microenvironments to evaluate the effects of modular stiffness,bioactive molecules and ECM proteins on stem cells.The microwells were fabricated by stamping a silicon substrate with previously spotted biomolecules on a hydrogel substrate with different PEG concentrations to alter stiffness[181].From screening these artificial niches,they studied the impact of laminin-1 and Jagged-1,the Notch ligand,on NSC fate[181].
However,when the microenvironment-based arrays are carried on a 2D platform,the cell–ECM interactions are weakened.For the sake of recapitulating the microenvironments and simulating the cellular statesin vivo,3D-based HTS platforms are recommended to evaluate the microenvironments proper to activate the specific cellular activities.Various microscale 3D culture screening systems mainly contain hanging drop plates,cellular microarrays and microwell plates(Supplementary Table 4)[67].Because hanging drop plates are not suitable for long-term culture,these platforms might be more suitable for neural differentiation or organoid formation.Cellular microarrays,as well as microwell plates,are more broadly used for 3D HTS protocols,as these two methods allow for longer culture periods and more stability than hanging drop plates.For cellular microarrays,cells could be premixed with gels and coprinted on the substrates with robotic arrayers(Figure 2).For instance,Fernandeset al[182]designed a dual-slide incubation method that included a methyltrimethoxysilane-coated glass slide with preprinted all-trans-RA and FGF-4 and another poly(styrene-co-maleic anhydride)-and a poly(L-lysine)-coated glass slide containing mESCs embedded in alginate spotted with a robotic spotter.Another common platform contains two complementary chips,a microwell and a microchip.The procedure involves spotting the mixture of cells and 3D matrices on the top of the micropillar and adding culture medium with screening candidates,such as small molecules and soluble factors,into the microwell.Then,by stamping and incubating two complementary slides,the small molecules and growth factors can diffuse into the cell spots and trigger biological reactions(Figure 2).In this way,multiple elements in microenvironments can be screened to study the regulation of stem cell fate[183-185].Given that microarray-based screening can result in interference between spots,microwell plates are also commonly used in 3D screening methods.Rangaet al[186]performed a 3D niche microarray on 1536-well plates that could control five characteristics: Matrix mechanical properties,ECM proteins,cell–cell interaction proteins,soluble factors and proteolytic degradability(matrix metalloproteinase sensitivity).Researchers cross-linked branched PEG-based macromers with specific peptide sequences susceptible to cell-secreted matrix metalloproteinases[186].Then,they encapsulated mESCs in 3D PEG gels to investigate mESC proliferation and self-renewal properties in different combinations of microenvironments.From the completed studies,we can learn that HTS platforms,especially 3D platforms,are tools with great potential for constructing microenvironments to discover combinations of elements that could well facilitatein vitroneurogenesis,such as NSC proliferation and differentiation,and even the internal signaling in charge of those cell behaviors.
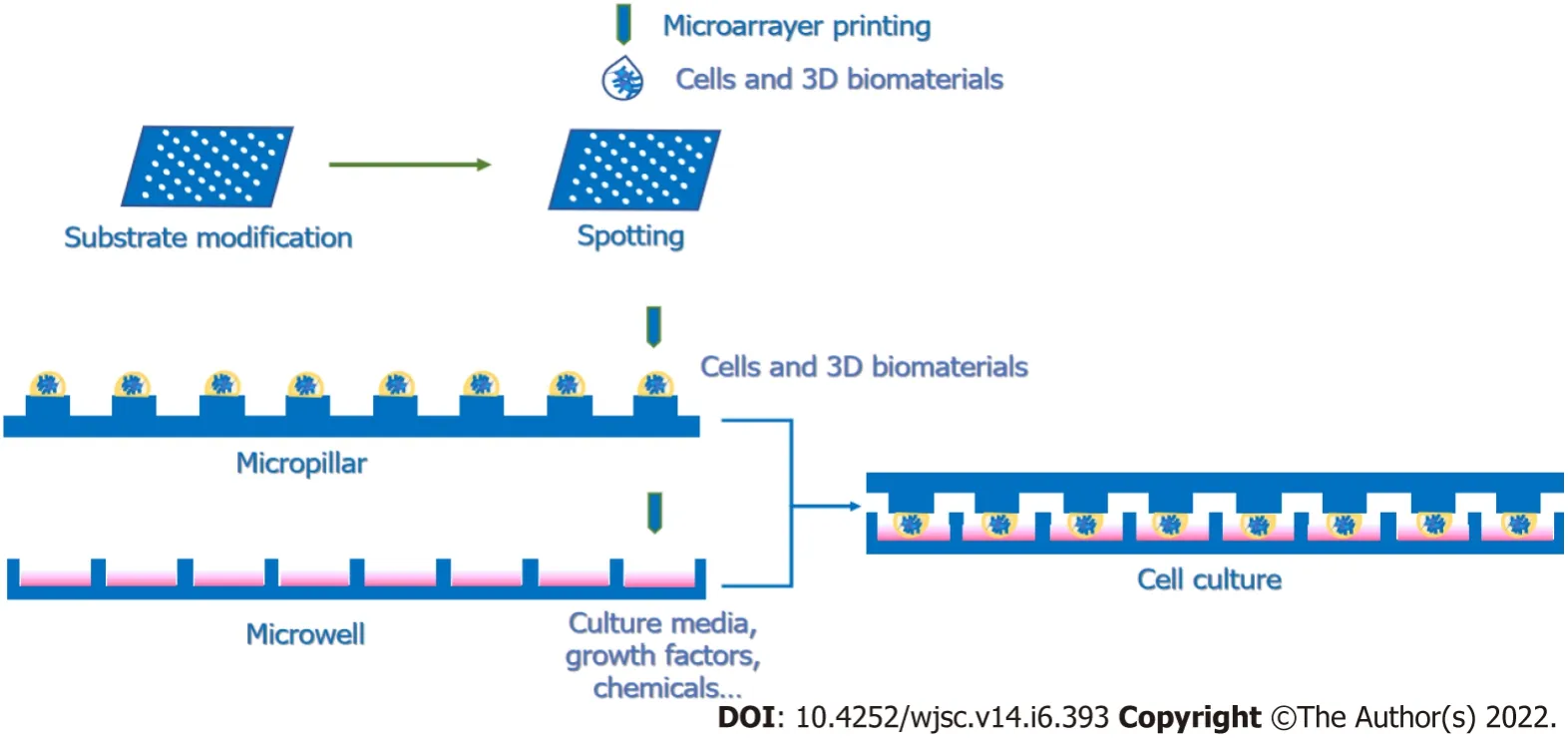
Figure 2 Procedures for 3D microenvironment screening.The mixture of neural cells and 3D biomaterials is printed through a microarrayer and subsequently spotted on the substrate of microarrays or micropillars.Then,culture medium is added to the microarray,and the micropillar is stamped into the microwell containing culture medium,including various combinations of growth factors and chemicals.
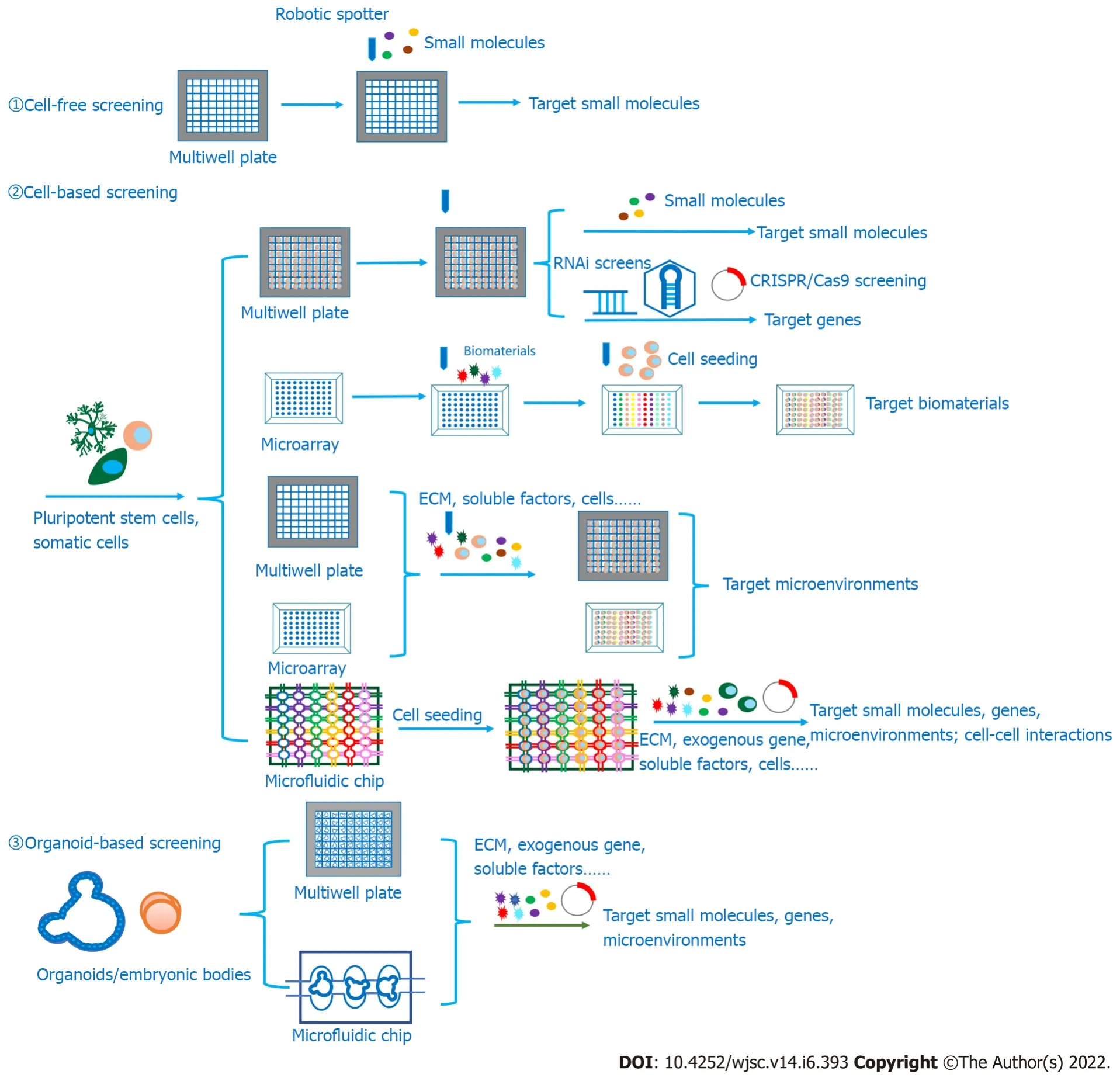
Figure 3 Summary of high-throughput screening platforms and their functions.Cell-free,cell-based and organoid-based HTS platforms can screen out target small molecules,genes,biomaterials and microenvironments to promote the survival,proliferation,differentiation and maturation of pluripotent stem cells,induce the transdifferentiation of somatic cells to other cell types,and even generate specific organoids for further applications,including disease modeling,development modeling,drug screening,tissue engineering and regenerative medicine.High-throughput screening(HTS)platforms include multiwell plates,microarrays and microfluidic chips,indicating the great potential in HTS in future studies.HTS: High-throughput screening;ECM: Extracellular matrix.
APPLICATIONS OF INDUCED NEURONAL CELLS
Since HTS technology can identify small molecules,specific genes and physiological microenvironments that contribute to neurogenesisin vitro,it is also important to focus on the clinical backgrounds and applications of the generated functional neurons or neural lineage cells using this technology.
Over the past 20 years,stem cell therapy and regenerative medicine have received considerable attention and have been expected to be applied in the treatment of CNS diseases,especially neurodegenerative diseases.The potential of stem cell therapy for CNS disease treatment lies in the capacity of NSCs to compensate for lost neurons with differentiated functional neurons,rebuild neural networks,secrete neurotrophic factors and reduce neuroinflammation to increase the survival rates of transplanted cells and healthy neurons[187,188].Before clinical trials,studies of stem cell transplantation were conducted on animal disease models to examine the safety and efficiency of stem cell therapy.Recently,various studies have been carried out using primary NS/PCs,neural precursors,immortalized neural stem cell lines,iPSC-derived NS/PCs,and directly reprogrammed neural precursor cells to treat spinal cord injury(SCI)[189,190],AD[191,192],PD[193-195],Huntington’s disease(HD)[196,197]and ALS[198]in animal models,and the symptoms of those diseases were relieved to some extent.For instance,the long-term selective stimulation of hM3Dq-expressing human iPSCs(hiPSCs)-derived NS/PCs enhanced their BDNF secretion,neuron-to-neuron interactions and synaptic activity in the surrounding host tissue in mouse SCI models,suggesting that enhancing neural activity and interactions with other cells in the neural circuit can be an effective way to improve the therapeutic effect of stem cell transplantation[199].Furthermore,the cografting of MSCs and dopaminergic precursors at the appropriate dose can help to enhance the survival of dopaminergic neurons in PD rat models[200].To better improve the survival,proliferation,differentiation and integrity of transplanted stem cells,growth factors[201,202],neurotrophic factors[203-206],hormones[207]and pharmacological treatments[208-211]have been applied with stem cells as methods of combination therapy.Additionally,genetic modification of engrafted stem cells can help to maintain long-term persistence,integrate into host tissues,and facilitate differentiation and maturation.The overexpression of growth factors[212],neurotrophic factors[213-218],morphogens[219]and transcription factors[220]in NS/NPCs is a widely applied method to further improve the therapeutic effect on CNS diseases and injuries.
For clinical studies,NPCs were transplanted into the dorsal putamina of patients with moderate PD,and a four-year evaluation was performed,which found this transplantation surgery to be safe and lacking in immune response or adverse effects[221].Motor improvement and enhanced midbrain dopaminergic activity were shown,although they decreased somewhat over four years[221].A longterm phase Ⅰ clinical trial also proved the safety of hNSCs and found a transitory decrease in the progression of the ALS Functional Rating Scale Revised up to four months post-transplantation[222].These results are promising,and in future studies,neural precursors derived from iPSCs or somatic cells,especially fibroblasts,can be applied to clinical studies to show safety and efficiency.For an allogeneic approach,developing an iPSC bank based on the human leukocyte antigen haplotype could provide more possibilities for stem cell therapy[223],and the genetic editing of patient-derived iPSCs is also a feasible approach for autologous stem cell transplantation[224].Meanwhile,exploring other cell sources for stem cell therapy could help broaden the field for clinical studies.Recently,a research group used NSCs isolated from midbrain organoids,which are generated from hPSCs,and transplanted them into rat PD models[225].The results showed midbrain dopaminergic neuron engraftment and reproducible behavioral restoration in those PD models[225].
Neural lineage cells generatedin vitroare also cell sources for neural tissue engineering,as they are becoming an attractive option for CNS disease treatment,considering that the support of 3D scaffolds can mimic the microenvironments that help the engrafted cells survive,integrate and differentiate.For brain injury repair,the injection of NSCs with a hyaluronate collagen scaffold loaded with controlled release of bFGF can recover cognitive function through the promotion of survival,differentiation and synaptic formation of NSCs in traumatic brain injury(TBI)rats[226].Chitosan scaffolds are also common options for the neural tissue engineering treatment of TBI in animal models[227-229].In addition to hydrogel materials,researchers also use porous scaffolds to prevent the collapse of scaffolds and provide enough space for neural differentiation,metabolic exchange,and neurite extension of grafted cells[230,231].For SCI repair,collagen microchannel scaffolds and gelatin sponge scaffolds carrying NSCs with drugs or neurotrophic factors have enhanced tissue repair efficiency in SCI animal models[232,233].Importantly,with the capacity to construct complex 3D microstructures,3D bioprinting techniques have been applied to create CNS architectures for regenerative medicine.For instance,a microscale continuous projection printing method can print 3D hydrogel scaffolds within 1.6 s to fit the size of injured spinal cords in rodent models;this technology is also scalable to human size[234].The 3D-printed neural tissues loaded with NS/PCs can integrate into host tissues,promote axon regeneration,and improve spinal cord functions in animal models[234,235].In clinical trials,the collagen scaffold(NeuroRegen Scaffold;NRS)has been used with MSCs to treat SCI patients,and the recovery of sensory and motor functions was observed[236].Another study transplanted NRS loaded with autologous bone marrow mononuclear cells into SCI patients,and in some patients,partial shallow sensory and autonomic nervous functional improvements could be observed,but the recovery of motor functions was not observed[237].Therefore,with the development of various 3D scaffolds and 3D bioprinting techniques,neural tissue engineering has great potential to contribute to CNS disease treatment in further studies.
Functional neurons can also be used forin vitromodeling to achieve a better comprehension of CNS diseases and neural development mechanisms.For neural disease modeling,AD models have been constructed utilizing hNPCs with familial AD(FAD)genes in Matrigel-based 3D culture systems,and aggregated p-tau proteins and amyloid-β deposits resembling AD pathology were observed[22].Furthermore,they showed that a high amyloid-β42/40 ratio could drive Aβ accumulation and phosphorylated tau protein accumulation in this 3D AD model[238].To recapitulate neuroinflammation in AD,they also conducted a 3D AD triculture model containing hNPC-derived AD neurons/astrocytes and subsequently plated microglia in the microfluidic platform[239].The results showed that migrating microglia,the upregulation of AD-related proinflammatory factors and the toxic effects of microglia on neurons and astrocytes could be observed in the 3D AD triculture model[239].In addition to AD models,PD models have also been established using 3D culture within vitroneural differentiation.Taylor-Whiteleyet al[240]first constructed a 3D PD model by differentiating human SH-SY5Y neuroblastoma cells into dopaminergic cells with RA and BDNF cultured in Matrigel[240].Next,they treated cells with preformed a-synuclein(a-syn)oligomers and observed a-syn-positive inclusions that resemblein vivoLewy bodies in morphology[240].Organoids have also been proven to be effective tools for 3D modeling.Kimet al[241]utilized hiPSCs with leucine-rich repeat kinase 2 G2019S mutation,which is a well-known trigger of late-onset familial and sporadic PD,to generate 3D midbrain organoids.From the 3D organoid model,they identified the TXNIP gene,which can contribute to the generation of α-syn in LRRK2-associated PD[241].Another research group built a 3D sporadic AD model by treating brain organoids with human serum to mimic the serum exposure caused by a blood–brain barrier breakdown in AD[242].AD-like pathologies could be observed in serum-exposed brain organoids,with increases in Aβ aggregates,phosphorylated microtubule-associated tau protein(p-Tau)levels,synaptic loss,apoptosis,and impaired neural networks[242].In addition,3D models aimed at other neural system diseases,including HD[243],hypoxic brain injury of prematurity[244]and brain tumors[245],have also been established.These 3D disease models provide us with feasible and valid platforms for future studies of disease pathogenesis and drug screening.
In addition to disease modeling,in vitro3D models can also be constructed to recapitulate neural development.To study neural tube morphogenesisin vitrothrough 3D culture,Rangaet al[23]first performed combinational HTS to screen out appropriate parameters of 3D matrices,based on which they investigated the effects of early developmental signaling molecules,including RA,Shh,Wnt-3a,BMP4 and FGF8,on dorsal-ventral(D-V)patterning with their 3D neural tube model.Another study cultured mESCs in Matrigel or defined 3D scaffolds containing laminin and entactin or PEG and induced floor plate formation and D-V pattering with RA[246].Marianiet al[247]induced human iPSCs to serum-free,floating embryoid body-like,quick aggregates with embryonic dorsal telencephalon properties,which could be used as anin vitro3D model for human cortical development.With the use of cerebral organoids,gene expression programs and epigenetic signatures during human brain development were recapitulated,as well as the interaction patterns between different brain regions[248-250].These models mimicking neural development could be applied to explore mechanisms underlying organogenesis and cell–cell interactions during neurogenesis[66,251]and could also be an option for studying neural genetic disorders[252],as well as a platform for drug screening.
LIMITATIONS AND PROSPECTS
Improving the conversion efficiency using small molecules and activating endogenous loci
Conversion efficiency is often discussed in articles focusing on cellular reprogramming and neural differentiation,which refers to the ratio of the target cell types to the initial cell types.The improvement of neural conversion efficiency is an important subject to address to increase the purity and efficiency of generated neurons for future clinical use.To overcome these difficulties,small molecules have been screened out to replace transcription factors,as small molecules can improve the conversion efficiency of cellular reprogramming compared to the overexpression of transcription factors[253].For instance,VPA can enhance the reprogramming efficiency of somatic cells to iPSCs[118,121],and CHIR99021,LDN193189,and A83-01 can further improve the neural induction rate[136].Significantly,the application of CRISPRa is capable of greatly enhancing reprogramming efficiency by targeting the human embryo genome activation-enriched Alu motif,leading to more efficient activation of Nanog and Rex1[254].Therefore,activating endogenous loci controlling cellular reprogramming and neural lineage induction can be an effective way to increase neural conversion efficiency.An alternative way is to selectively ablate proliferative cells and keep functional neurons for the sake of guaranteeing the safety and efficiency of stem cell transplantation.This research was performedviapharmacological activation of the suicide gene within weeks after transplantation,and the yield of dopaminergic neurons and the recovery of motor functions were not affected by diminishing the graft size in the PD rat model[255].Thus,in future studies,HTS technology can still play an important role in screening small molecules and endogenous genes,which can aid in improving the conversion efficiency and generate more functional neurons.
Developing an organoid/spheroid-based HTS system
Over the decades,testing probes of HTS methods have been developed from molecules to cells and even to tissues/microenvironments and organoids.Currently,molecular,cell-and tissue-based screening systems have come into use,and it is also quite important to investigate organoid-based HTS devices with increased producibility and reduced heterogeneity between batches,allowing for large-scale screening[256].Jorfiet al[257]screened FAD-mutated hNSCs or iPSC-derived neurospheroids with a 96-well cell culture plate with 1536 microwells.They embedded the neurospheroids in Matrigel and screened several chemical compounds to assess their impact on neural differentiation[257].The establishment of a high-throughput bioengineered human cardiac organoid in the 96-well format was also reported[258].After that,105 hit compounds from approximately 5000 candidates,which were screened from iPSC-derived cardiomyocytes in the 2D platform,were screened over a 3-log scale concentration range that requires approximately 1000 human cardiac organoids to develop compounds with the capacity for cardiomyocyte proliferation[259].Another study generated kidney organoids from hiPSCs utilizing multiwell plates,and this HTS-compatible platform was used to screen out an inhibitor of nonmuscle myosin II ATPase activity as a specific activator of polycystic kidney disease cystogenesis in organoids[260].Renneret al[261]also developed an automated workflow that could integrate midbrain organoid culture,immunostaining and high-content imaging for high-throughput chemical screening using a 96-well format,which could save manual operation and improve the compatibility of organoid culture and HTS.Although high-content imaging analysis has been a powerful tool to evaluate organoid generation,for brain organoids,it is probable that the evaluation of neural circuit dynamics,such as that through 3D microelectrode arrays,could become a standard in upcoming studies[262,263].Furthermore,combinations with the automated workflow of organoid culture and artificial intelligence can shed light on CNS disease modeling and drug discovery for clinical trials[264].
Improving microfluidic-based HTS systems
Over the years,HTS devices have been developed,ranging from multiwell plates to microarrays;notably,microfluidic devices are gradually showing their features in HTS technology.The lab-on-chip method has contributed to this development.Schudelet al[265]designed a microfluidic chip to separate cell clusters by dividing the chip into one part for siRNA patterning and another for target screening to study virus–host interactions.Furthermore,to improve the screening efficiency after cell transduction,Wanget al[266]designed a droplet-based microfluidic platform compatible with single-cell screening to identify the yeast Saccharomyces cerevisiae with elevated protein production through RNAi screening and searched for genetic targets capable of improving protein secretion.Hanet al[267]first utilized a CRISPR/Cas9 screen on a microfluidic platform,also called a microfluidic separation chip,on which cells transduced with the lenti-CRISPR kinase library were sorted to examine transport distances to evaluate cell deformability[267].For chemical screening,Titmarshet al[268]constructed a high-density microbioreactor array that could provide 8100 chambers for the proliferation of hPSCs or hPSC-derived cardiomyocytes.They found that CHIR99021 showed the best effect on human cardiomyocyte proliferation among purmorphamine,IGF-1 and FGF-2[268].Although the microfluidic array could provide thousands of chambers as reactors,the numbers of candidates allowed for one screening are usually limited.Thus,exploiting microfluidic devices that are able to hold more isolated channels for screening more candidates at a time has great potential.
However,the HTS platforms currently available for cell/organoid-based screening are mainly well plates that lack automation and integration and commonly cause reagent waste.Therefore,in further research,the microfluidic platform shows great promise to achieve a higher throughput and autocontrolled and integrated properties.For instance,Schusteret al[269]designed an automated microfluidic 3D cellular and organoid culture platform for the culture of pancreatic ductal adenocarcinoma organoids generated from single cells from patients.The platform could contain 20 independent experimental conditions and 200 individual chambers that are large enough to hold growing organoids[269].The researchers performed dynamic and combinational drug screening and recorded the incidences of cellular apoptosis and death to evaluate the treatment effect of the temporal drug combinations.
In addition,microfluidic devices could also be utilized to perform cell coculture using droplet-based microfluidic systems,which could function in studying the microenvironments of cell–cell interactions under high-throughput conditions[270].Other researchers have also designed high-throughput 3D coculture systems on microfluidic chips[271,272].With these methods,HTS could be performed on these platforms to screen out 3D microenvironments containing cellular interactions,such as synaptic connections between neurons and astrocytes.In addition,microfluidic chips have been applied in generating concentration gradients of biomolecules to study steepness-dependent neural chemotaxis on high-throughput 3D platforms[273].Rifeset al[274]constructed a microfluidic platform to generate gradients of CHIR99021 to activate Wnt signaling,and they modeled neural tube development in this 3D microfluidic system.Therefore,in future studies,HTS will be performed on microfluidic systems due to their capacity to better recapitulate the microenvironmentsin vivo,which is a strategy that shows great promise.
CONCLUSION
HTS technologies are playing increasingly important roles in neurogenesisin vitrodue to their ability to screen out crucial genes controlling neural lineage determination,small chemical molecules regulating cell fate,and microenvironments,including 3D matrices,soluble factors,physical parameters and interactions with other cell types(Figure 3).After screening out suitable microenvironments,these culture conditions could be applied in generating mature and functional neurons,neural tissues and organoidsin vitrofor further applications,such as 3D modeling and drug screening,to investigate neural diseases or developmental mechanisms and explore medical solutions.With the requirements of 3D models,3D-based screening with tissues or organoids is developing to better evaluate screening outcomes from an overall perspective than molecular or cell-based screening can.Meanwhile,the screening devices are trending toward minimization,automation and integration,from multiwell plates to microarrays and microfluidic devices,to conduct the screening process in a high-throughput manner that requires less time and consumes fewer reagents.Today,the need for combinational screening is growing,as investigations of the interactions between different drugs or environmental factors are vital to developing combined therapies and novel culture conditions.In addition,it is notable that microfluidics makes it easier to perform high-throughput combinational screening with nanodroplets and microwell array plates that can hold only two nanodroplets in a well[275].Overall,from past studies and due to the fast development of HTS devices,we anticipate that HTS technologies will be able to make great contributions toin vitroneurogenesis and solve other problems in regenerative medicine in future studies.
To conclude,HTS technology could help to dissect the mechanisms of genetic regulation during neurodevelopment,identify niche-targeted small molecules and secreted factors to promote endogenous NSC activation for clinical treatment,and screen out biomaterials and other microenvironment elements to generate more functional and mature neurons with specific subtypes and improved purity,which could be used to establish 3D neural disease or developmental models.Although some 3Din vitromicroenvironments cannot be reconstructed based on HTS at the present time,we still predict that HTS will be a promising tool for defining microenvironments for higher efficiency modelling.
ACKNOWLEDGEMENTS
We thank the Biological and Medical Engineering Core Facilities of Beijing Institute of Technology.
FOOTNOTES
Author contributions:The review was conceived and designed by Qing H,Zhao J and Zhang SY;The study was drafted by Zhang SY;The review was revised by Qing H,Zhao J and Zhang SY;The review was discussed by Qing H,Zhao J,Ni JJ,Quan ZZ,Li H and Zhang SY.
Supported byNational Natural Science Foundation of China,No.81870844,No.82001167 and No.82101394.
Conflict-of-interest statement:All the authors report no relevant conflicts of interest for this article.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers.It is distributed in accordance with the Creative Commons Attribution NonCommercial(CC BYNC 4.0)license,which permits others to distribute,remix,adapt,build upon this work non-commercially,and license their derivative works on different terms,provided the original work is properly cited and the use is noncommercial.See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORCID number:Shu-Yuan Zhang 0000-0001-6925-2140;Juan Zhao 0000-0002-9719-751X;Jun-Jun Ni 0000-0001-8486-8146;Hui Li 0000-0002-7392-0233;Zhen-Zhen Quan 0000-0002-2557-0371;Hong Qing 0000-0003-2820-450X.
S-Editor:Fan JR
L-Editor:A
P-Editor:Fan JR
杂志排行

World Journal of Stem Cells的其它文章
- Disagreements in the therapeutic use of mesenchymal stem cellderived secretome
- Adipose tissue in bone regeneration-stem cell source and beyond
- Role of stem cells-based in facial nerve reanimation:A meta-analysis of histological and neurophysiological outcomes
- Long noncoding RNAs in mesenchymal stromal/stem cells osteogenic differentiation:Implications in osteoarthritis pathogenesis