枝角类亚化石揭示的湖北太白湖过去百余年环境演变*
2022-06-30林涌权董旭辉廖粤军羊向东JeppesenErik
林涌权,董旭辉,2,廖粤军,李 燕,2,羊向东,Jeppesen Erik
(1:广州大学地理科学与遥感学院,广州 510006)(2:广州大学气候与环境变化研究中心,广州 510006)(3:广东省韶关市田家炳中学,韶关 512026)(4:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京 210008)(5:丹麦奥尔胡斯大学生物科学系,斯尔克堡 8600)
近百年来,在全球生态环境问题突出、人为干扰强烈和气候变暖的大背景下[1],湖泊环境亦遭受到了不同程度的破坏[2],引发系列环境后果,导致水环境质量不断下降、资源枯竭、功能退化[3],而且环境恶化驱动因素复杂多样,治理难度大[4].例如在人类活动强烈的东部平原湖区普遍出现的湖泊富营养化问题成因上,一方面与气候变暖[5]、风速变化[6]及流域(或湖泊内部)水动力条件[7]有关,另一方面也受到人类围垦、流域工农业营养盐污染和水产养殖等活动有关,引起部分藻类和水生生物过度繁殖,造成水体的富营养化[8].这些要素不仅会产生叠加交互效应,而且在不同的时间段内存在强度差异,加剧了湖泊生态系统修复的难度[9].厘清湖泊复杂系统状态转变过程中不同驱动要素的贡献是生态系统治理的重要前提[10],而且必须从生态系统的整体、长时间序列的角度上去予以科学的评估[11].
现代湖泊监测记录通常较短(<50年),无法提供完整、有效的长期生态系统演变信息,而湖泊沉积物的连续记录可以揭示湖泊及其流域长期生态环境变化过程,揭示其演化的规律和原因[12].自1980s以来,基于湖泊沉积物的湖泊古生态学日益受到重视,诸多湖泊水生生物如摇蚊、硅藻等残体内由于有难以降解的几丁质、硅质等成分,可以长期保存在湖泊沉积物中,因此可以提供丰富的历史水生生态系统不同组分的生物群落信息[13].随着科学界对生物群落演化规律的认识,特别是生物非线性响应规律、复杂系统论等的兴起,当前古生态学的研究已经由前期的单一指标、线性响应、特定指标的定量重建过渡到多指标比对、临界突变、系统动力学的研究趋势[11,14-15].无疑,挖掘新的生物指标、加强多生物指标的综合分析应该是今后古生态学研究的一个必然趋势,这不仅可以对不同指标独立的逻辑解释起到相互验证后补充的作用,而且可以提供生态系统层面的信息,包括系统各要素相互作用、食物网结构和功能、生境和物种多样性、以及系统的物质和能量循环等信息[16].
湖泊水体中的枝角类既是浮游植物的主要牧食者,又是浮游生物如食性鱼和一些无脊椎动物的主要食物,从而在水生态系统的营养传递和能量流动中起着极为重要的作用[17].因此,湖泊中枝角类的种群结构变化信息及其与环境因子之间的关系不仅是淡水生态学研究中的重要内容,对于湖泊生态修复也具有重要的参考意义[18].枝角类死亡后,其几丁质残体能很好地保存在湖泊沉积物中,使得枝角类有望成为古湖沼学研究中的重要生物指标[19].国际上已经开展了众多湖泊枝角类种群的现代环境监测及沉积物中亚化石的环境重建工作,但主要侧重于高山、北方寒冷区的湖泊[20-22];国内目前开展的枝角类亚化石研究也主要集中在云南地区[23-25],对东部平原湖区仅有Dong等对60余个湖泊表层沉积物[26]、刘桂民在太湖[27]、Cheng等在升金湖[28]有相关研究报道.我国东部洪泛平原湖泊众多,长期受到多重人类活动与自然要素的共同影响,湖泊生态系统类型独特,进一步明晰枝角类亚化石的环境指示意义、揭示其群落演化的长期趋势与演化机制对湖泊环境治理具有重要的意义.
本文拟通过古湖沼学的研究方法,选择东部典型浅水湖泊太白湖开展枝角类群落的长期生态学研究.近150年来,长江中下游的太白湖经受了严重的水文条件改变和湖泊富营养化问题,导致水质恶化,生态系统发生草-藻型稳态转换.目前一些研究工作对转换机制已有一定认识,特别是,对古生态指标开展了大量的相关研究.例如羊向东、董旭辉等提取该湖沉积物中记录的硅藻信息,重建了历史的营养演化过程[29-30]; 仝秀芳等利用孢粉组合与碳屑指标分析恢复了近1500年以来流域植被的变化及其驱动因子[31]; 张恩楼等通过北部湖区沉积摇蚊亚化石分析,再现了1950年前后摇蚊群落对富营养化水平增加的响应[32]; 赵雁捷等[33]和Xu等[34]则从不同的角度对太白湖历史环境演化机理进行了深入剖析.但直到目前尚未开展该湖沉积物中枝角类亚化石的研究,特别是枝角类群落与其余古生态指标所揭示的湖泊环境变化特征是否相一致仍然知之甚少[30,32,35].本文拟补充同一钻孔中的枝角类属种丰度信息,来识别太白湖近150年来枝角类群落变化的主要特征,结合环境监测数据和社会经济数据,探讨枝角类群落的演化机制; 对比其他常用的古生态指标硅藻、摇蚊等的响应差异,籍此评估亚热带湖泊中枝角类亚化石指标在重建历史环境变化中的应用前景,丰富湖泊古生态指标体系,并为定量古生态学提供更多的科学依据.
1 材料与方法
1.1 研究区概况
太白湖(29°56′~30°01′N,115°46′~115°50′E)是长江中下游洪泛平原湖泊群中的一个小型浅水湖泊,位于中国湖北省黄冈市黄梅、武穴两县市交界处,现今湖泊总面积约为25 km2,平均深度3.2 m.太白湖流域地处亚热带季风区,年均温度18.4℃,多年平均降水1273 mm,蒸发量1041 mm,水热条件好,湖水依靠地表径流和湖面降水补给,承雨面积960 km2[36].太白湖流域经济类型以农业为主,主要种植水稻、小麦和油菜等; 1950s中期至1978年太白湖流域内掀起了大规模的围湖造田潮,直到1970s末期才逐渐放缓.太白湖水产养殖开始于1950s,1980s后期发展为网箱养殖,渔场调整养殖结构,增加了滤食性鱼种并投放大量草鱼、蟹去除水草,从自然放养发展到集约式养殖模式围网养鱼并投放饵料(包括化肥),进一步导致营养富集,近年来的水体主要水质指标参数如表1所示.
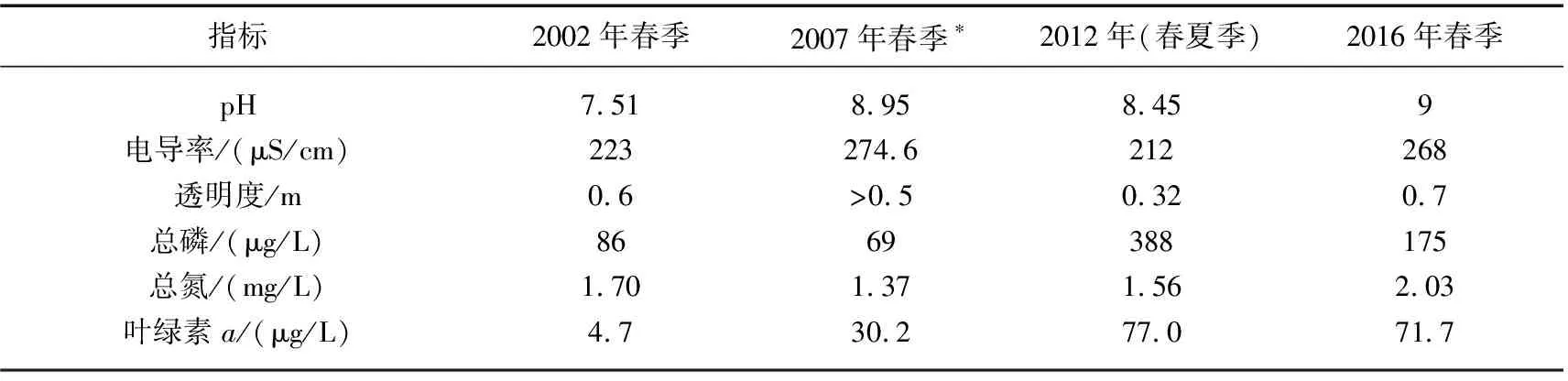
表1 太白湖主要水质参数多年变化
1.2 样品采集与实验室分析
本研究基于2007年5月在太白湖北部湖区所采集获得的柱状沉积岩芯(TN1孔,采样点位置为29°59′44.3″N,115°48′26.5″E,采样水深2.5 m)的上部42 cm样品,沉积柱上部20 cm按0.5 cm间隔,20~40 cm按1 cm间隔,40 cm以下按照2 cm间隔分样.该沉积柱年代的计算参照刘恩峰等已发表结果[38],上部42 cm涵盖了自1860s以来的湖泊沉积物.沉积物多指标包括硅藻、摇蚊、元素等的分析方法及结果参考文献[30-33,38-39].
枝角类前处理参考Frey的方法[40]进行: 称取约5 g且经充分混匀的样品,首先将样品置入备有200 mL 10% KOH溶液的烧杯中,然后水浴锅对烧杯加热(60℃,1 h)以对样品解絮凝,随后样品用 80 μm的筛网过滤,残余在筛网上的物质用蒸馏水进行清洗收集,定容至10 mL并加入数滴甲醛防止腐烂.
枝角类的鉴别在光学显微镜下进行.先将样品充分摇匀,取100 μL悬浮液置于计数框,在100~400倍显微镜下鉴别、计数.枝角类的鉴别主要依据当前国内外的枝角类鉴定图版[41-42]进行.由于枝角类微化石多以不同部位的形式存在,因此对某一种/属的计数要根据其不同的残体部位分别计数,最终以此种/属数量最丰富的残体部位的数量作为计数结果,每个样品总的个体计数至少要达到100以上.枝角类丰度换算成绝对丰度(每克干重沉积物中的枝角类数量)和相对丰度(某一种枝角类占所有枝角类总数的比例).枝角类百分比图谱使用Tilia Graph程序绘制,属种组合带的划分基于CONISS聚类分析结果[43].
1.3 数理统计分析
非度量多维尺度分析(Non-metric multidimensional scaling,NMDS)是一种将多维空间的研究对象简化到低维空间进行定位,分析和归类,同时又保留对象间原始关系的数据分析方法,可用于揭示样品间的相对差异大小及生物群落组合变化的总体趋势(用各个轴的得分来衡量,第一轴归纳了最主要的变化趋势)[44-45].对枝角类数据,选择在两个以上的样品中出现,含量至少在一个样品中超过2%的常见属种进行数值分析.最后数据包括了62个层位样品和30个主要枝角类属种类型.同样,对同一沉积柱中的硅藻和摇蚊群落数据进行NMDS分析,分别计算第一轴的得分,用以表示各个生物群落的总体变化特征.NMDS分析在软件PAST[46]中完成.
对于不同生物门类群落状态发生显著变化时间点的检测,本研究采用Rodionov等提出的检测平均水平显著变化的突变检测分析方法(sequential t-test algorithm for analyzing,STARS)[47].通过STARS计算得到枝角类、硅藻及摇蚊3个生物门类时间序列上的突变点.在Excel中加载STARS模块进行分析,本次研究参数设置显著度0.05,步长为10,通过计算状态均值变化指数(regime shift index,RSI)来表示状态间变幅大小.
枝角类与鱼类关系密切,Jeppesen等研究揭示出溞属(Daphnia)占溞属(Daphnia)和象鼻溞属(Bosmina)总量的比例(palaeolimnological predation index,PPI)与单位浮游生物食性鱼类捕获量呈显著负相关关系[48].因此,我们利用PPI来指示浮游生物食性鱼类丰度变化.建立该指数的长期变化有望与实际的渔业产量数据进行对比,进而评估利用枝角类群落来定量重建历史时期渔业产量方法的有效性.
2 结果
2.1 枝角类组合
太白湖钻孔中共鉴定出30个枝角类属种,尽管所有层位浮游种所占比例较高(接近80%),其中象鼻溞属Bosmina为岩芯柱的绝对优势属(平均百分含量达到75%),Bosminacoregoni为岩芯柱的绝对优势种,平均百分含量达到32%.然而沿岸/底栖/附生种的属种种类数目更多,以小型底栖种Alona的各种类型为主,其余的附生属种如Chydorussphaericus和Sidacrystallina亦有较高出现率.整个沉积柱中溞属(Daphnia)的相对丰度均较低(<2%).根据CONISS分析把枝角类群落划分为4个组合带,各个组合带变化特征如下:
组合带1(42~35 cm,对应1860-1910年).此时期以B.coregoni、Bosminasp.、小型Alonasp.和Bosminalongirostris为主,在1905年B.coregoni相对丰度最大,可达42.9%,总体上呈增加趋势.组合带1中沿岸种如小型Alonasp.、Acroperusharpae和Camprotercusreciirostris等相对丰度呈现降低趋势.
组合带2(35~25 cm,对应1910-1944年).此时期沿岸种Alonarectangulatype、小型Alonasp.、C.sphaericus和S.crystallina丰度变化趋势并不明显,浮游种变化不一,B.longirostris和Bosminasp.丰度增加,B.coregoni丰度减少.
组合带3(25~17 cm,对应1944-1965年).此时期沿岸种小型Alonasp.、C.sphaericus和S.crystallina的丰度总体增加,浮游种B.longirostris、Bosminasp.和B.coregoni丰度整体上相比第二阶段,有减少趋势.
组合带4(17 cm以上,对应1965-2006年).此时期沿岸种A.rectangulatype、小型Alonasp.、C.sphaericus和S.crystallina的丰度在第三阶段增加后保持稳定高值,Ilyocryptussp.首次出现.但浮游种仍占绝对优势,浮游种B.longirostris相比第三阶段丰度减少,而Bosminasp.和B.coregoni丰度保持稳定(图1).
2.2 枝角类种群NDMS分析
枝角类数据的NDMS结果中,前两个轴的特征值分别为0.99和0.14,解释属种数据的方差值分别为73.68%和10.13%.在属种分布图上,富营养种B.coregoni、B.longirostris等出现在图的右边,代表了轴一的主要变化方向; 而沿岸种如小型Alonasp.、A.harpae、C.reciirostris等则出现在图的左边.样点的排列主要受第一轴的控制,同图1中的聚类分析结果一致,从左到右4个组合带的样品依次排列; 第二轴上样品的排列主要反映了带内的差异(图2).
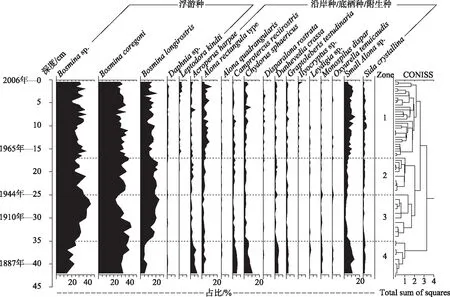
图1 太白湖沉积岩芯枝角类主要属种分布
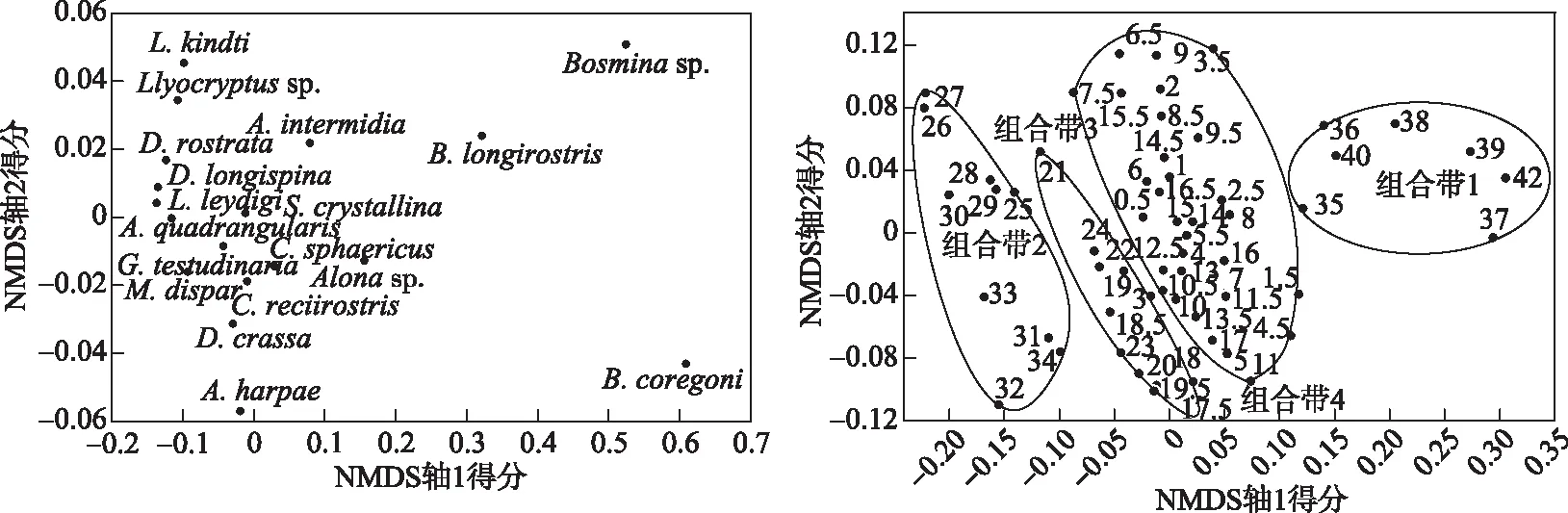
图2 太白湖沉积岩芯中枝角类数据NDMS排序分析结果(右图中数字为样品深度, cm)
2.3 枝角类对湖泊鱼类产量的指示意义
如图3所示,过去50年内,太白湖渔业经历了3个显著的增长时期: 1980年前和1985-2000年间的渔业产量呈缓慢上升趋势; 2000年后出现较大增幅(年产量从300 t增长至3600 t).古湖沼捕食指数(PPI)在0.54~0.62区间内波动变化,推导的鱼总产量波动整体上升趋势明显,特别是1985年后的两次增产过程均有反映,在渔业产量增长的初期(1965-1974年)PPI亦能较好地指示该过程.尽管1975-1982年间两者相关系数较差,但总体上PPI能一定程度上较好的刻画历史时期鱼类产量的变化趋势.
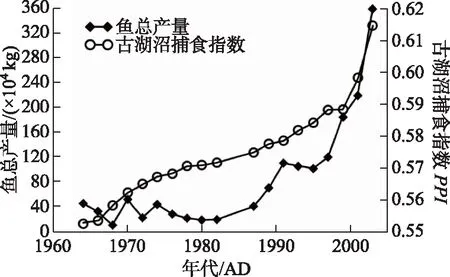
图3 1960s以来太白湖鱼类产量和古湖沼捕食指数PPI
2.4 多种古生态指标的对比
枝角类群落演替的STARS分析结果表明,在1910s前后枝角类发生了一次显著的属种演替(突变显著度RSI=3.1),随后直至1960s前后发生第二次显著的群落演替(RSI=-0.8较小),分别对应了浮游种Bosmina属的增加-减少及底栖属种Alona、C.sphaericus等的减少-增加的过程(图1,图4).
摇蚊和硅藻群落演替阶段则体现出不同的阶段性.摇蚊群落在1940s前始终保持相对稳定的结构,此后开始缓慢变化,年间变率相对稳定,直至1990s开始逐渐稳定,转变至富营养化群落状态,累计变率RSI=2.7.而硅藻群落的第一次显著变化发生于1900s,稍早于枝角类群落,随后群落变率较大,呈现波动状态,直至1985年群落变率回落(稍早于摇蚊群落的1990年),维持较为稳定的群落结构(图4).
总体上枝角类、摇蚊及硅藻3种生物门类的总体变化体现一定的差异性.从群落变率范围来看,3个生物门类大致相当,如第一阶段(1910s前)变率范围(NMDS得分)分别为0.10~0.25(均值0.18)、0.10~0.16(均值0.12)、0.15~0.22(均值0.19); 从变化的时间点来看,3种生物门类不同的演化阶段尽管存在5~10年的差异但大体重合(图4),例如枝角类和硅藻在1900s前后的第一次显著变化的时间点大体一致,而摇蚊和硅藻则在富营养化的加剧过程中体现出较好的一致性(如1990s前后的第二次群落转换).
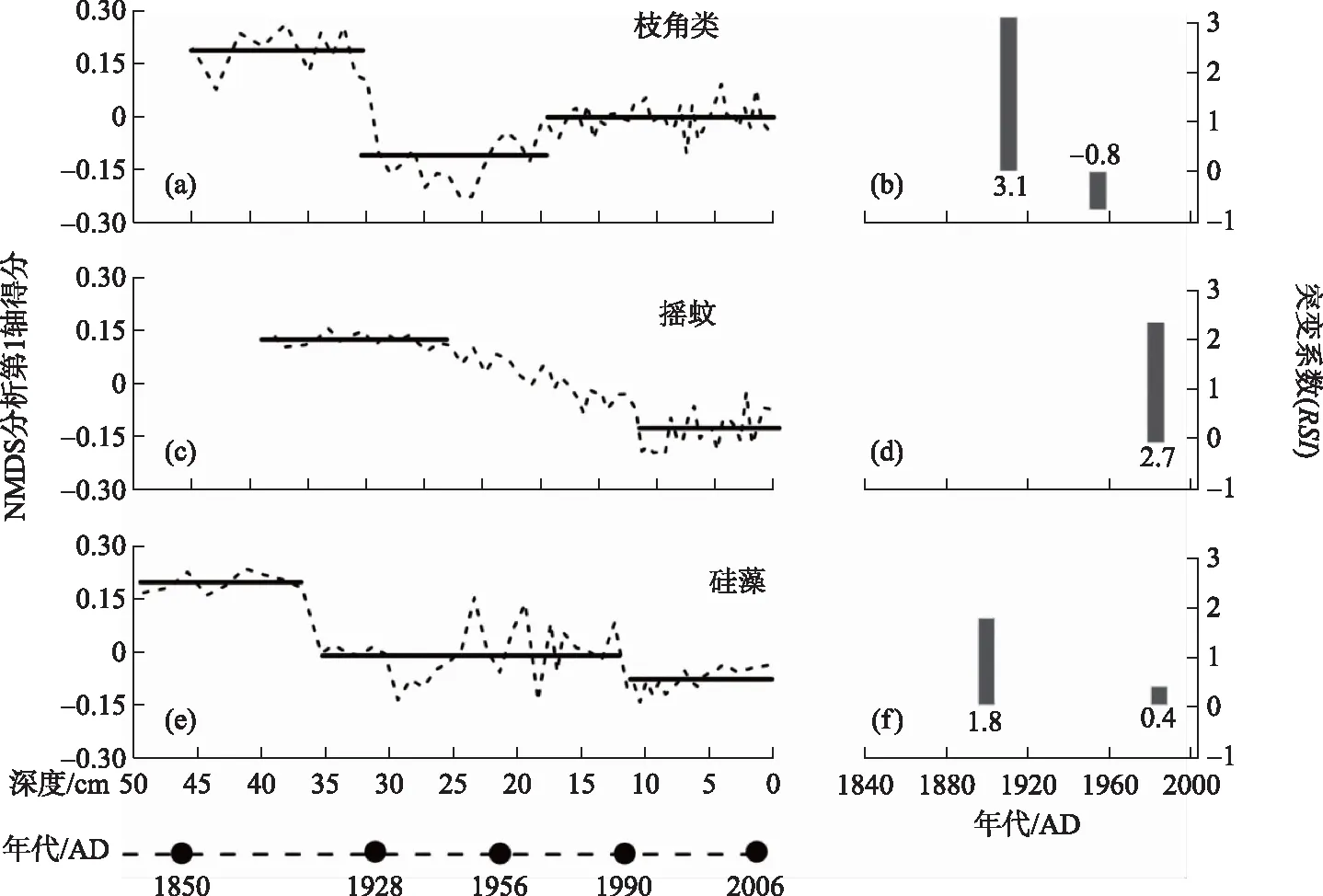
图4 太白湖各生物门类(a~b: 枝角类,c~d: 摇蚊,e~f: 硅藻)NMDS分析第一轴得分的状态转换过程检测结果(其中黑色实线代表种群平均状态,黑色棒状图的高度及下方数值代表状态转变系数RSI大小)
3 讨论
3.1 太白湖枝角类基本特征
太白湖枝角类以浮游种Bosmina为主,同时出现属种种类繁多但含量均较低的底栖、附生属种,这些属种均广泛出现于长江中下游其他湖泊中.作者团队曾于2012年前后对长江中下游60个湖泊的现生枝角类(表层沉积物中的群落)进行系统调查,共发现35个属种,其中28个属种为附生/底栖类型[26],97%的湖泊中均出现了浮游种Bosmina及底栖属种Alona、C.sphaericus,反映了亚热带浅水湖泊中枝角类群落的相似性[28,49].
沉积序列的NMDS排序分析能有效地揭示样品间的群落差异,进而揭示潜在的影响要素.图2可见第1轴的特征值远高于第2轴,表明太白湖沉积柱中枝角类属种的分布主要受第1轴环境因子的影响,根据以上主要属种生态特征,B.coregoni、B.longirostris和A.rectangulartype是偏好富营养水体的枝角类[49],从左到右,枝角类属逐渐从贫营养化指示性组合到富营养化指示性组合转变,因此第一轴大致反映营养梯度的变化.相应地,从左到右,样品点中的排列指示了各层为沉积时期湖泊环境状态的变化.枝角类组合的NMDS排序结果与CONISS聚类分析结果一致,响应了近150年来太白湖4个主要环境演化阶段.
3.2 枝角类群落揭示的湖泊环境变化
结合太白湖环境监测数据、社会经济数据以及多沉积指标数据,重建太白湖的环境主要演化阶段如下:
1860-1910年,此时期江湖自然联通,水动力条件强,存在季节性水位波动和周期性冲刷,太白湖水位较高且变化明显.湖水营养水平低,但入湖营养物质增加.枝角类组合以浮游类型Bosminasp.占优势,且呈现先轻微减少而后不断增加的趋势,表明在湖泊与长江自由联通的背景下[2],太白湖水动力条件强,水体更新快,水质洁净,与此时期太白湖频率磁化率高、中值粒径大和硅藻类组合以浮游类硅藻占优势等证据吻合(图5).枝角类附生种(如小型Alonasp.、A.harpae和C.reciirostris)的减少反映了当时江湖联通引起的季节性水位波动和周期性冲刷及旱涝指数先减后增,暗示该阶段水位先降低后升高,并一直保持较高水位,与枝角类响应的水位波动结论一致.指示富营养化的B.coregoni丰度增加则揭示了当时随着入湖营养物质的增加,根据记载[31],虽然此时期湖水总磷继续维持中营养水平,但因流域森林植被发育,湖泊生产力缓慢提高,因此喜好营养的枝角类丰度一度上升.
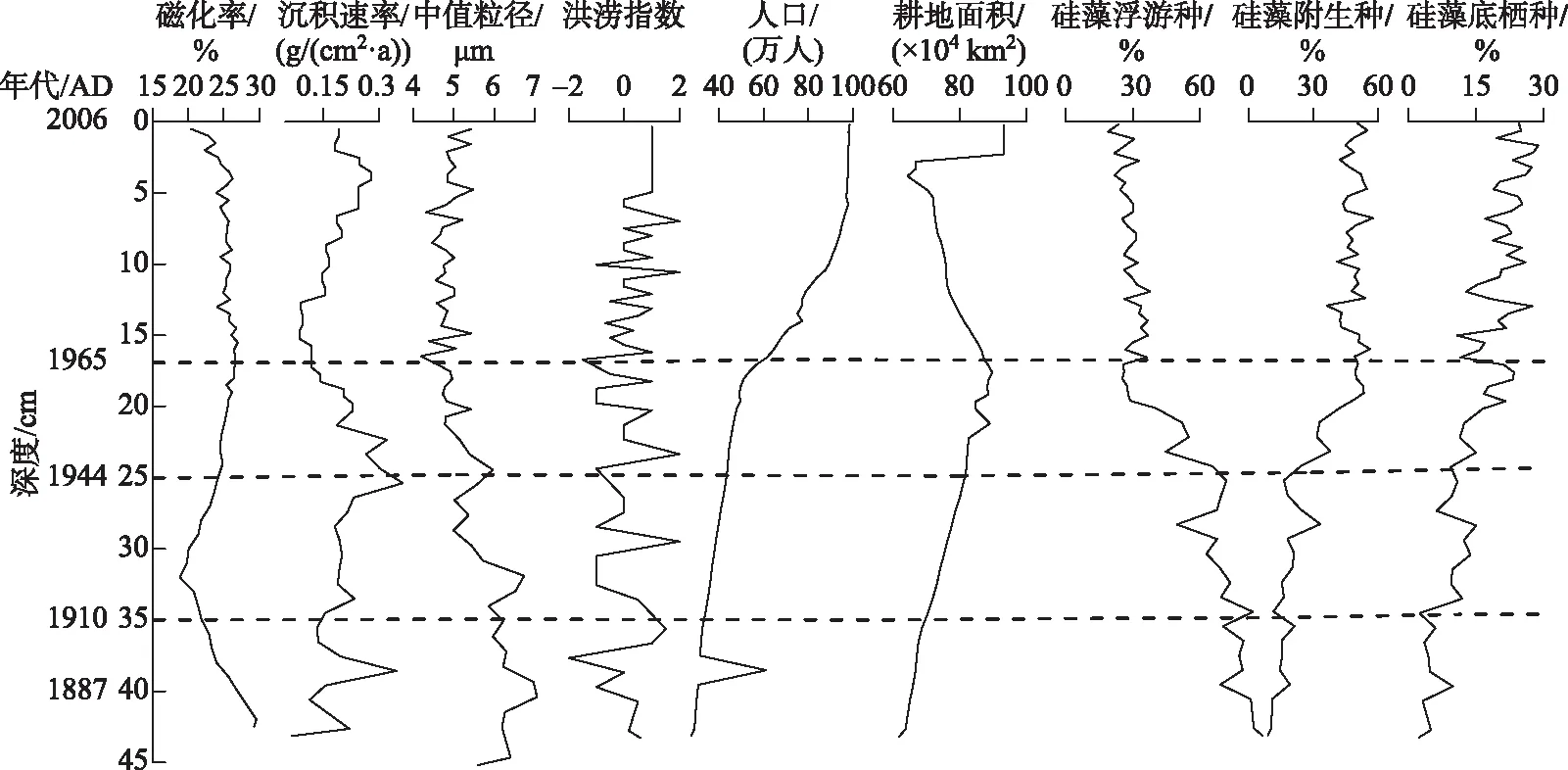
图5 太白湖钻沉积物指标、流域内旱涝情况、人口、耕地面积情况及不同生态类型的硅藻属种百分比(组合带对应于枝角类的组合划分)
1910-1944年,此时期江湖仍保持较高的连通性,捕食压力增强,人类活动开始加强但尚不剧烈,太白湖面积缩小,水位下降.根据记载,此时期太白湖已经成为当地水产农业的重要产地[29],鱼类喜好的枝角类浮游种B.coregoni丰度减少,指示了食物网中捕食压力的加大.而指示富营养化的枝角类浮游种B.longirostris和Bosminasp.丰度相比上一时期增加,则表明此时期人类活动的持续加强,从而引起入湖营养物质的增多,太白湖流域沉积通量和人口数量增加也能印证此时期人类活动的加强.后期枝角类沿岸种小型Alonasp.的增加和中值粒径减小则反映了湖水水位有所上升.该时期内的枝角类沿岸种数量上变化幅度小,整体上处于低谷平缓期.
1944-1965年,此时期人类活动明显加强,修闸建坝后江湖连通性开始变差,水生植被快速扩张.此时期枝角类沿岸种丰度的总体增加响应了太白湖的修闸建坝事件.1955年太白湖下游建成梅济新闸,修闸后湖水通过梅济港向东排入龙感湖,而后进入长江,水位直接由梅济堤闸人工调控,减小了江水倒灌对太白湖的影响,江湖连通性开始变差.1958-1963年间太白湖上游陆续建成了仙人坝水库、考田水库、荆竹水库[50],湖泊水动力减弱,换水周期变长,流速变缓,水生植物能够扎根生长并快速扩张,有利于沿岸/附生种枝角类的生长,这与硅藻类型由浮游类型迅速向附生类型组合演替表明水生植被快速扩张的研究结果高度一致[33].但枝角类增加的幅度并不大,反映了枝角类指标响应的滞后性,由于营养级的传递等原因,可能导致了浮游生产力增加的滞后.
1965-2006年,此时期随着人口数量的急剧增加,人类干扰进一步加剧,快速扩张的水产养殖和农业发展加大了湖泊的环境压力,江湖连通性差,水动力减弱.枝角类沿岸种的增加更明显地响应太白湖修闸建坝所引起的水动力下降事件,并与浮游种硅藻的减少、中值粒径和频率磁化率快速下降相互印证,水动力减弱,水位变化减小,入湖的泥沙减少,换水周期拉长,加上太白湖流域在1960s起开始广施化肥[18],太白湖过度的养殖和化肥的大量投入使得水质持续下降,1987年以来在该湖西面的武山湖周围相继建立了数个小型化工厂[32],入湖磷及污染负荷明显增加[29],因此指示富营养化的枝角类浮游种数量保持着动态平衡的高值.
3.3 多门类古生态指标对环境变化的响应
不同的生物门类对外界环境胁迫响应的敏感性和弹性存在较大差异.本研究揭示出在最初的环境变化情况下,硅藻群落最先发生显著变化(37 cm处,约为1900 AD),随后枝角类和摇蚊群落相继发生显著的变化(分别为35 cm处和27 cm处),均滞后于硅藻的响应.从第一次显著的群落演变幅度来看,枝角类变幅最大(RSI=3.1),而硅藻和摇蚊变幅则相对较小(RSI分别为2.7和1.8).这三者中,硅藻系初级生产者,生命周期较短(1~2周),直接受到湖泊水体状况(如pH值、水体营养、气候条件等)的影响[8],因此对环境变化响应最为敏感.而摇蚊大多是底栖类型动物,主要受底泥中的食物供给、溶解氧浓度、温度、水深等要素的影响[50],而且可以通过迁移、新陈代谢改变等来适应恶劣的环境[51],因此体现出一定的生态弹性(如图4c中12~27 cm间缓慢的变化),而且在重富营养化环境下,仍然稍稍滞后于硅藻对营养富集的响应(1985年硅藻群落变率回落,稍早于摇蚊群落的1990年,图4).
枝角类位处食物网的中间位置,在水生态系统的营养传递和能量流动中起着极为重要的作用[48].相应的,枝角类群落演替不仅会受到外界理化环境变化(如温度、水位、酸碱度等)的影响,还会受到食物网“营养级联效应”(如水生植被的多寡、鱼类数量)的影响[26,52]; 此外,枝角类在水深-光照度适应、逃避捕食者及物种进化等方面亦具备较高的灵活性[42],因此它相对于硅藻及摇蚊一定程度上存在更为复杂的影响机制.本研究中枝角类于1960s前后的系统状态转变过程,在硅藻及摇蚊群落中均未体现(图4),事实上,1960s以来太白湖TP和TOC含量整体呈上升趋势[33],但喜富营养环境的B.longirostris(较B.coregoni更偏好富营养环境)在1960s以来有较明显下降,因此枝角类的群落变迁应该还受到营养要素之外的作用力,例如湖泊水文条件改变的影响.历史文献记载[33],1955年太白湖下游建闸,修闸后湖泊水位直接受梅济堤闸人工调控,减小了江水倒灌对太白湖的影响.1958-1963年期间太白湖上游陆续建成了水库,使湖泊水动力进一步减弱[33,38,50].因此,枝角类亚化石能捕捉到历史系统演化轨迹中一些独特的生态系统信息.无疑,如何去解释枝角类这些不同的响应差异,是今后开展基于古生态学指标的环境重建研究的重要内容.
3.4 枝角类的古生态学意义
已有研究表明,枝角类是湖泊食物链上的重要组成部分,具有生命周期很短且响应快速,及分类清晰等优点[52],能够指示湖泊的环境指标如温度、盐度、营养、pH和食物网中生物的相互作用(如捕食、竞争)等[10,26],可用于生态响应模式的识别,从而实现湖泊历史变化从定性分析到定量反演的转变,还可用于确立湖泊治理的基准环境[53-54].例如,水生生态系统中历史鱼类的数量可通过鱼类的鳞片残体或DNA技术来予以重建[8],但由于鱼类残体浓度往往偏低,使得鱼类数量重建始终是一个难点.本研究展示了基于枝角类种群PPI指数对历史时期浮游性捕食鱼类数量的重建,能有效地反映太白湖历史时期的渔业养殖状况(图3).无疑,多古生态指标的不断扩充、方法体系的不断完善,可以为其他指标(如硅藻、摇蚊、介形类、色素、GDGT等)的古环境重建提供佐证,能提供认识过去湖泊环境变化历史更为全面的视角,有利于厘清湖泊演化的规律和原因,模拟和预测湖泊未来的变化趋势,进一步指导管理者更科学地、有针对性地管理湖泊[15-16].
和其他古生态指标一样,基于枝角类指标的过去环境重建也存在一些局限性.首先是沉积物中的枝角类与同期水体中现生的群落构成存在一定差异.尽管大多数枝角类的几丁质外壳能较好的保存[55],但是沉积过程会受多种因素如水流和湖底地形的影响,造成湖心沉积岩芯中的枝角类组合会过高于地表征浮游种枝角类而降低沿岸种枝角类的真实丰度[56]; 同时,由于枝角类容易被其他浮游动物捕食并消化,这样并非所有枝角类都能在沉积物中得到保存,这与枝角类几丁质外壳的厚薄程度、湖泊特征相关,如在上文太白湖沉积岩芯主要枝角类属种分布图中,象鼻溞属Bosmina丰度远高于其他属种,这与其较硬的外壳导致的保存性较好相关,这种不同属种在沉积物中的代表性将直接影响到定量重建的结果.其次,枝角类因处于食物网的中间位置,受到的影响要素众多,因此枝角类种群演替的驱动要素会随着外界环境发生变化,因此对单一环境要素的指示意义相较于硅藻、摇蚊等偏弱.太白湖长期的枝角类演化序列表明水位、营养、水生植被类型等均是可能的影响要素,如果就单一要素进行重建,可能存在较大误差,必须基于多影响要素的模式(如回归树模型)来进行重建[55].
基于上述枝角类指标应用的潜在缺陷,未来需加强下列研究内容: 首先,需要进行枝角类属种更多的现代生态学研究,完善不同区域枝角类群落个体生态学信息(如生态位、生理习性等),这些信息将有助于解释过去环境的变化; 其次,加强枝角类的埋藏学研究,建立不同属种的现生-沉积间的定量关系,对不同保存状况的属种进行校正[56],特别的,近期的枝角类被证明具有较短的进化周期,沉积物里的枝角类休眠卵的复活生态学研究已成为一个新兴的研究热点[57].最后,需加强更多不同古生态指标的挖掘工作,例如环境DNA方法的应用[58].本研究表明不同的生物群落对外界环境扰动呈现出不同的响应特征,多门类的指标体系有利于量化特定群落差异,提取丢失的环境演化信息.
4 结论
1)太白湖枝角类具有显著的亚热带浅水湖泊种群特征,以浮游种Bosmina为主,同时出现种类繁多的底栖、附生属种.沉积枝角类演替特征揭示了过去百余年太白湖4个环境演化阶段: 1910年前江湖自然联通,太白湖水位较高且波动明显,水动力条件强; 1910-1944年人类活动渐强但尚不剧烈,入湖营养缓慢增加,水位下降; 1944-1965年人类活动明显加强,营养累积,水生植被快速扩张; 1965年后人类干扰进一步加剧,修闸建坝后江湖连通性开始变差,水动力减弱,流域工农业的发展使得湖泊富营养化严重、水质下降,水生植被退化.
2)不同的生物门类对外界环境胁迫响应的敏感性和弹性存在较大差异.本研究揭示出枝角类相对于硅藻及摇蚊具有不同的响应特征(突变时间、幅度均存在差异),因此能提取额外的生态-环境演化信息.
3)枝角类亚化石是一种有效的重建历史环境的古生态指标,在重建鱼类产量方面有独特优势,但亦存在沉积代表性差异、影响要素众多、部分属种生态信息缺乏等问题,今后需加强枝角类个体生态学及埋藏学等研究,加强古今生态学间的融合.
致谢:感谢中国科学院南京地理与湖泊研究所张恩楼研究员提供摇蚊数据,伦敦大学学院Helen Bennion教授和南京信息工程大学Giri Kattel教授均提供了有益的建议.
