草地贪夜蛾在玉米-小麦寄主转换中的转录组分析
2022-06-29王斯亮闫成进
王斯亮,邵 越,闫成进
(1.温州科技职业学院 农业与生物技术学院,浙江 温州 325000; 2.温州市植物保护与土壤肥料管理站,浙江 温州 325000)
草地贪夜蛾()原产于美洲热带和亚热带地区,具有迁飞习性,自2016年开始入侵非洲以来,现已扩散至世界各地,是一种破坏力强的重大入侵害虫。草地贪夜蛾寄主范围广,已知能为害76科353种植物,主要包括禾本科106种、菊科31种和豆科31种,重点为害玉米、小麦、水稻、高粱、甘蔗、棉花等作物。草地贪夜蛾由于不同的寄主偏好性,分化形成了“玉米型”(主要取食玉米、棉花和高粱)和“水稻型”(主要取食水稻和牧草)两个亚群,二者在形态学上没有显著差异,但是在交配行为、信息素成分等方面有明显的差别。张磊等鉴定我国的草地贪夜蛾入侵种群来源于美国的佛罗里达州,该种群大部分呈现核基因“玉米型”,线粒体基因“水稻型”的特征,故称之为“特殊玉米型”,推测其可能为“水稻型”母本与“玉米型”父本杂交产生。国内目前报道的草地贪夜蛾寄主植物主要包括玉米、小麦、水稻、青稞、甘蔗、花生、马铃薯、甘蓝等。
草地贪夜蛾适生区域广。北回归线以南地区,草地贪夜蛾可以周年繁殖,无滞育。在中国,长江中下游地区和东南丘陵地带是草地贪夜蛾的高度适生区。然而冬季草地贪夜蛾的越冬虫态和冬季寄主是需要调查研究的问题。入侵我国的草地贪夜蛾以“玉米型”和“特殊玉米型”为主,虽然草地贪夜蛾本身具有多食性,但在饲喂不同寄主植物的实验中,草地贪夜蛾明显表现出对玉米的偏好。在广东地区的1—3月,草地贪夜蛾仍然能以幼虫虫态为害并安全越冬,并且为害分布与冬种玉米区范围一致,极少数在甘蔗上发现,而在香蕉、花生、马铃薯等作物未见草地贪夜蛾。安徽和江苏地区均在10月发现草地贪夜蛾为害早播小麦,其中安徽寿县小麦平均被害株率高达77.51%,苏北地区小麦平均被害株率高达52.60%。12月上旬以后,主要在云南、四川冬麦区发现草地贪夜蛾为害。已有多种实验表明,草地贪夜蛾取食小麦能够完成生活史。由此可见,小麦是草地贪夜蛾的潜在越冬寄主之一,我国南方的冬小麦种植区,尤其是前茬种植玉米的小麦田,很可能成为草地贪夜蛾的冬季庇护场所。本研究通过转录组测序,分析草地贪夜蛾在取食玉米和小麦后基因表达水平变化,探究草地贪夜蛾在寄主转换过程中的内在适应性机制。
1 材料与方法
1.1 试虫来源
草地贪夜蛾幼虫于2019年10月16日在温州市藤桥镇温州种子种苗科技园(北纬28°5′42″,东经120°30′59″)玉米上采集,在人工气候箱(RXZ-500D-LED,宁波江南仪器厂)内培育数代,箱内温度25 ℃,湿度70%,光照周期为12 L/12 D。
1.2 取食转换实验
用玉米叶喂养初孵幼虫直至5龄初(蜕皮后24 h内),将幼虫放入35 mm培养皿中单独饲养,分成3组,每组分别饲喂玉米叶、小麦叶、人工饲料(参照华南农业大学提供的配方,主要成分为黄豆粉、麦麸等)。3 d后,提取单头幼虫总RNA,每组做3次生物重复。
1.3 RNA提取
将草地贪夜蛾幼虫放入试剂RNA-easy Isolation Reagent(南京诺唯赞生物科技有限公司)中研磨,参照该试剂说明书对应的方法操作提取RNA,并通过紫外分光光度计检测RNA的浓度和纯度,通过1%琼脂糖凝胶电泳检测RNA的完整性。
1.4 转录组测序与拼接
转录组委托南京集思慧远生物科技有限公司采用Nova seq6 000平台进行测序,测序读长为PE150。测序前使用Nanodrop和Agilent 2 100检测RNA的浓度和纯度以及完整性。测序得到的原始数据Raw Data,通过去除测序接头,过滤低质量序列,得到高质量的Clean Data,并使用软件FastQc(v0.11.8)对Clean Data进行质量控制,计算Q30等指标。使用软件Trinity(v2.4.0)对得Clean Data的Reads进行拼接组装,生成每个基因的转录本序列(transcript),取每条基因中最长的转录本作为Unigene。
1.5 Unigene功能注释
使用软件BLAST将Unigene序列与NR(Non-Redundant Protein Sequence Database)、Swiss-Prot、GO(Gene Ontology)、COG(Clusters of Orthologous Groups)、KOG(euKaryotic Orthologous Groups)、KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库比对,参数设置E-value<10。预测Unigene的氨基酸序列,使用软件HMMER将氨基酸序列与Pfam(Protein family)数据库比对。综合以上数据库比对结果,获得Unigene的功能注释信息。
1.6 差异表达分析
采用软件Bowtie2(V4.4.7)将测序得到的Reads与Unigene库进行比对,根据比对结果,结合软件RSEM进行表达量水平估计。利用FPKM(Fragments Per Kilobase of exon model per Million mapped fragments)值表示对应Unigene的表达丰度。差异表达分析中,使用R(v3.6.2)中的DESeq2程序包计算差异倍数(FC,Fold Change),筛选标准为|log2(FC)|≥1且FDR<0.05。筛选到差异基因后,根据超几何分布原理进行差异表达基因的GO富集分析和KEGG富集分析。
2 结果与分析
2.1 草地贪夜蛾幼虫转录组测序
本实验中,被测的草地贪夜蛾幼虫分为3组,其中处理组2组,分别是饲喂小麦叶的FAWw组和饲喂人工饲料的FAWsl组,对照组1组,为饲喂玉米叶的FAWym组。取5龄初幼虫(蜕皮后24 h内)开始饲喂,3 d后取样提取总RNA,每组3个生物重复,共9个RNA样本进行转录组测序。样本测序并过滤得到Clean Data(表1),结果显示,Q30的比例均达到92%以上(碱基质量值Q30对应碱基识别精度达到99.9%),未识别碱基(N)的比例均为0,表明测序数据可靠。
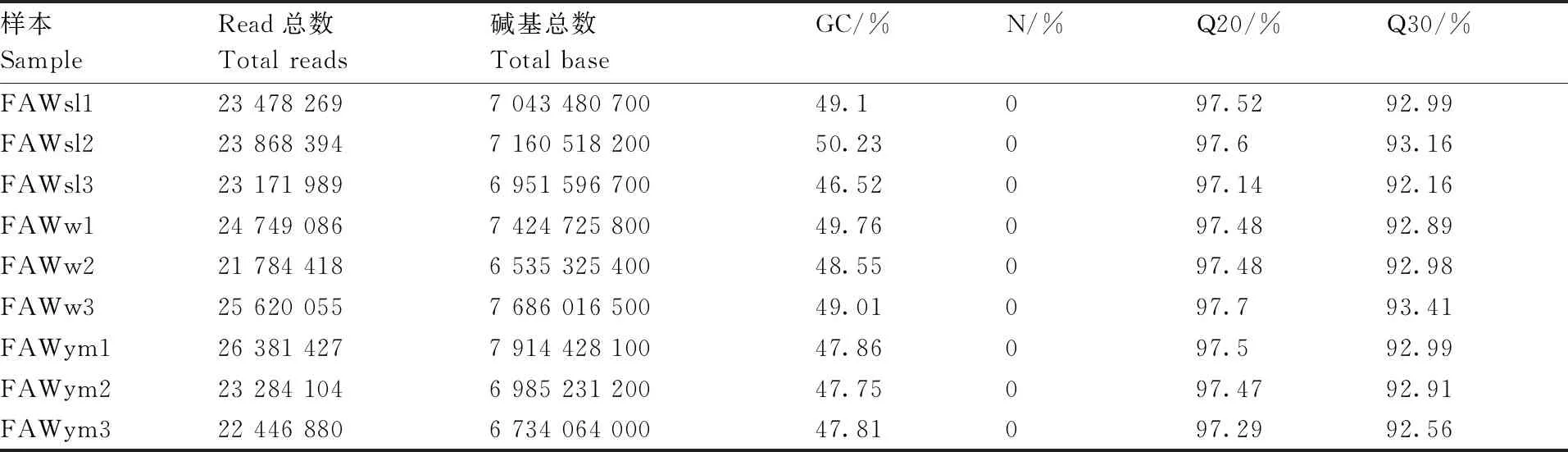
表1 草地贪夜蛾幼虫转录组测序结果(Clean Data)
将Clean Data中的Reads进行混合组装,最终得到66 407条Unigene,平均长度为1 063.99 bp,N50长度为1 768 bp。组装的Unigene序列长度全部大于300 bp,其中,长度大于1 000 bp的有17 957条,占27.04%的比例,长度大于2 000 bp的有8411条,占Unigene总数的12.67%(表2)。
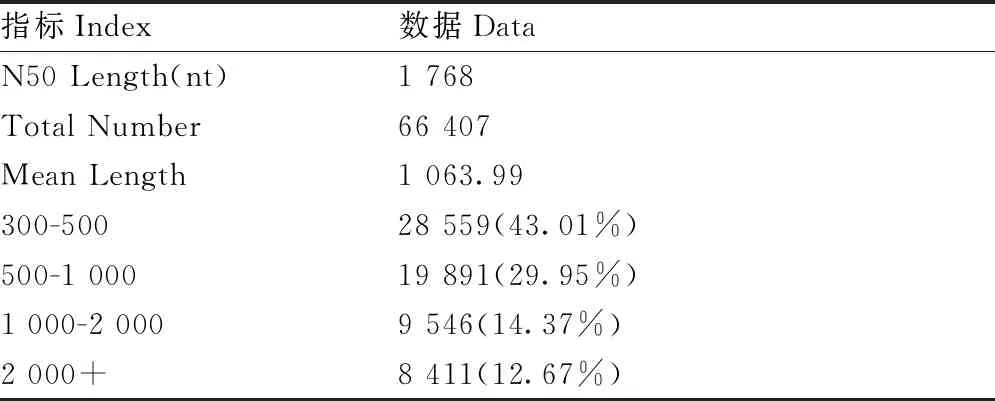
表2 草地贪夜蛾幼虫转录组组装结果(Unigene)
2.2 Unigene功能注释
将组装得到的草地贪夜蛾幼虫Unigene序列,与NR、Swiss-Prot、GO、COG、KOG、KEGG数据库进行比对,将Unigene的预测氨基酸序列与Pfam数据库进行比对,最终获得18 868条有注释信息的Unigene(表3)。其中在NR数据库中注释到的Unigene有18 645条,占比98.82%。通过NR库的注释信息,获得草地贪夜蛾幼虫Unigene序列与近缘种属的同源序列相似情况,其中匹配最多的是斜纹夜蛾的同源序列,共有13 248条,占NR注释Unigene总数的71.05%,其次是棉铃虫(5.77%)和烟芽夜蛾(5.49%)(图1)。草地贪夜蛾与斜纹夜蛾为同属近缘种,该结果表明Unigene注释信息可参考程度较高。
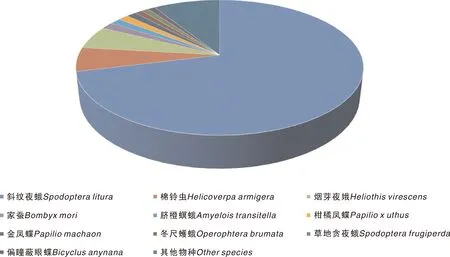
图1 草地贪夜蛾幼虫转录组Unigene在NR数据库的同源物种分布
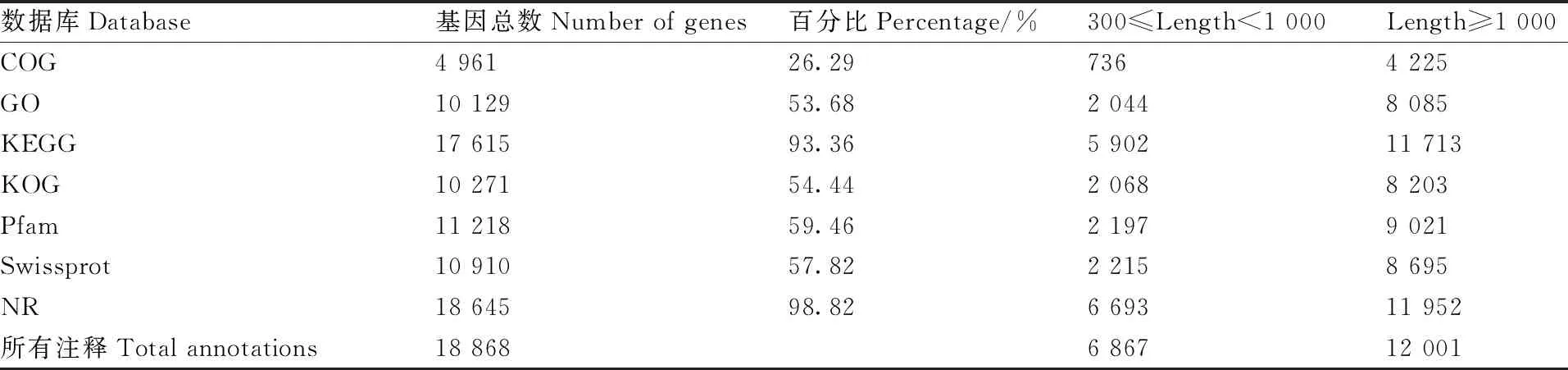
表3 草地贪夜蛾幼虫转录组Unigene功能注释
2.3 取食转换后的差异表达基因
以取食玉米叶的草地贪夜蛾幼虫为对照组(FAWym),与取食饲料3 d后的草地贪夜蛾幼虫(FAWsl)相比,共找到6 863个差异表达基因,与取食小麦叶3 d后的草地贪夜蛾幼虫(FAWw)相比,有542个差异表达基因(表4),其中共同的差异表达基因有384条。在差异表达基因列表中除去未获得注释的基因,得到取食饲料后(FAWsl组)的差异表达基因4 607条(上调表达基因2 364条,下调表达基因2 243条),取食小麦叶后(FAWw组)的差异表达基因432条(上调表达基因263条,下调表达基因169条),共同的差异表达基因304条,其中在FAWw组和FAWsl组中均表现上调表达的基因有82条,均表现下调表达的基因有98条(表5)。

表4 差异表达基因相关统计
在FAWw组的上调表达基因中,有82条基因是在FAWsl组中也表现上调表达的共同上调表达基因,有94条基因属于共同差异表达基因中在FAWsl组呈现下调表达的基因,87条基因为特异性上调表达基因(表5)。去掉82条共同上调表达基因后进行分析,发现FAWw组表达量显著上调的基因有大量涉及表皮相关功能,包括表皮蛋白(cuticle protein),类似兜甲蛋白(loricrin-like),类似节肢弹性蛋白原(pro-resilin-like),表皮生长因子受体激酶(epidermal growth factor receptor kinase),内表皮结构糖蛋白(endocuticle structural glycoprotein)等。共有32条表皮相关基因,其中27条在FAWw组上调表达,在FAWsl组下调表达(表6)。该结果表明不同食物对草地贪夜蛾幼虫的表皮发育有不同影响。
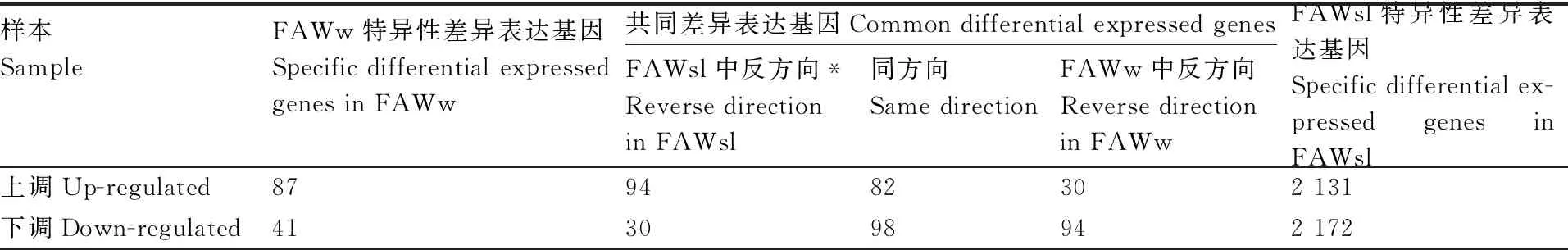
表5 特异性差异表达基因与共同差异表达基因
细胞色素P450(cytochrome P450)被认为与多食性昆虫的寄主适应性相关。FAWw组的差异表达基因中,细胞色素P450基因有16条,主要包含CYP4,CYP6,CYP9等家族的基因。其中下调基因有13条。在下调表达的细胞色素P450基因中,有5条属于FAWw组特异性下调基因,有8条属于共同下调基因(表6)。另外有2条解毒酶基因也在FAWw组和FAWsl组中共同下调,分别是谷胱甘肽S转移酶(glutathione S-transferase,GST)基因和过氧化物酶(peroxidase)基因。

表6 草地贪夜蛾取食小麦后(FAWw)的功能性差异表达基因
2.4 差异表达基因富集分析
GO包含3个大类,分别是生物学过程(Biological Process),细胞组分(Cellular Component),分子功能(Molecular Function)。通过GO对差异表达基因进行功能富集分析,得到在生物学过程大类中,FAWw组的上调表达基因的富集趋势与总差异表达基因的富集趋势相似,表现为在免疫系统过程(immune system process)和细胞杀伤(cell killing)功能上显著富集(图2)。富集显著程度排名靠前(前20项)的二级功能术语(Term)中,有4条术语涉及免疫功能,分别是防卫细菌反应(defense response to bacterium,GO:0042742),外来细胞杀伤(killing of cells of other organism,GO:0031640),防卫原生动物反应(defense response to protozoan,GO:0042832),抗真菌体液免疫反应(antifungal humoral response,GO:0019732)。而FAWsl组的差异表达基因在免疫功能上没有明显的富集(图2)。在细胞组分大类中,FAWw组和FAWsl组的差异表达基因在功能富集分析上差异不大,富集显著程度最高的二级功能术语均为胞外区(extracellular region,GO:0005576)和细胞器膜(organelle membrane,GO:0031090),每个术语内的上调和下调表达基因数量相近。在分子功能大类中,表皮结构成分(structural constituent of cuticle,GO:0042302)是FAWw组和FAWsl组的差异表达基因富集显著程度最高的二级功能术语,然而在FAWw组表现为上调基因在表皮结构成分中显著富集,在FAWsl组则是下调基因在表皮结构成分中显著富集,表明取食小麦后可能有利于草地贪夜蛾幼虫表皮的生成。
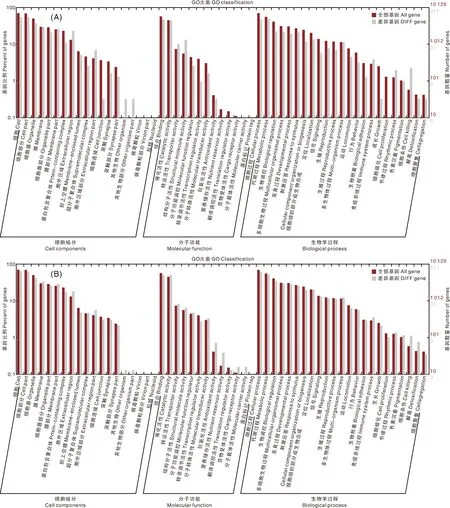
A,草地贪夜蛾幼虫取食小麦后(FAWw)的差异表达基因GO富集分析;B,草地贪夜蛾幼虫取食人工饲料后(FAWsl)的差异表达基因GO富集分析。
KEGG代谢通路富集分析结果显示,FAWsl组的上调基因富集的代谢通路有蛋白酶体(Proteasome),内质网蛋白质加工(Protein processing in endoplasmic reticulum),DNA复制(DNA replication),错配修复(Mismatch repair)等,这些通路都与生长发育密切相关。下调基因富集的代谢通路主要与解毒代谢有关,如过氧化物酶体(Peroxisome),细胞色素P450代谢外源物质(Metabolism of xenobiotics by cytochrome P450),药物代谢-细胞色素P450(Drug metabolism-cytochrome P450),药物代谢-其他酶(Drug metabolism-other enzymes),谷胱甘肽代谢(Glutathione metabolism)等(图3)。人工饲料相比与植物,有毒次生代谢物明显减少,因此,幼虫的解毒代谢活性显著下降。取食小麦的FAWw组,上调基因主要在甘氨酸、丝氨酸和苏氨酸代谢(Glycine, serine and threonine metabolism),半胱氨酸和甲硫氨酸代谢(Cysteine and methionine metabolism),DNA复制,内质网蛋白质加工等通路富集。下调基因富集较显著的通路有过氧化物酶体,苯丙氨酸代谢(Phenylalanine metabolism)等(图4)。表明取食小麦后,幼虫氨基酸代谢和转换反应更活跃。
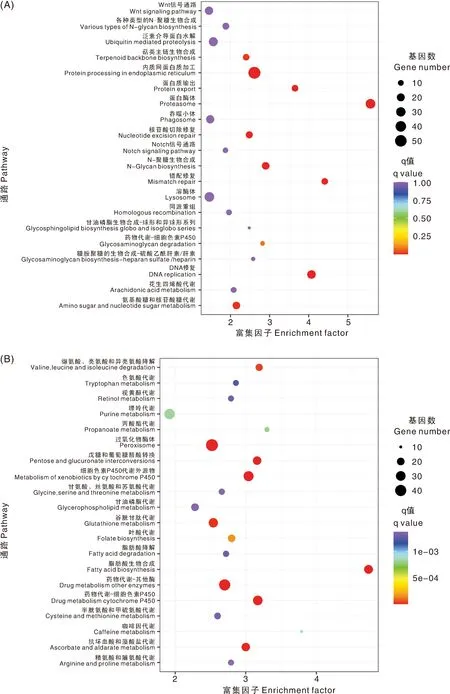
A,FAWsl上调表达基因的KEGG代谢通路富集分析;B,FAWsl下调表达基因的KEGG代谢通路富集分析。
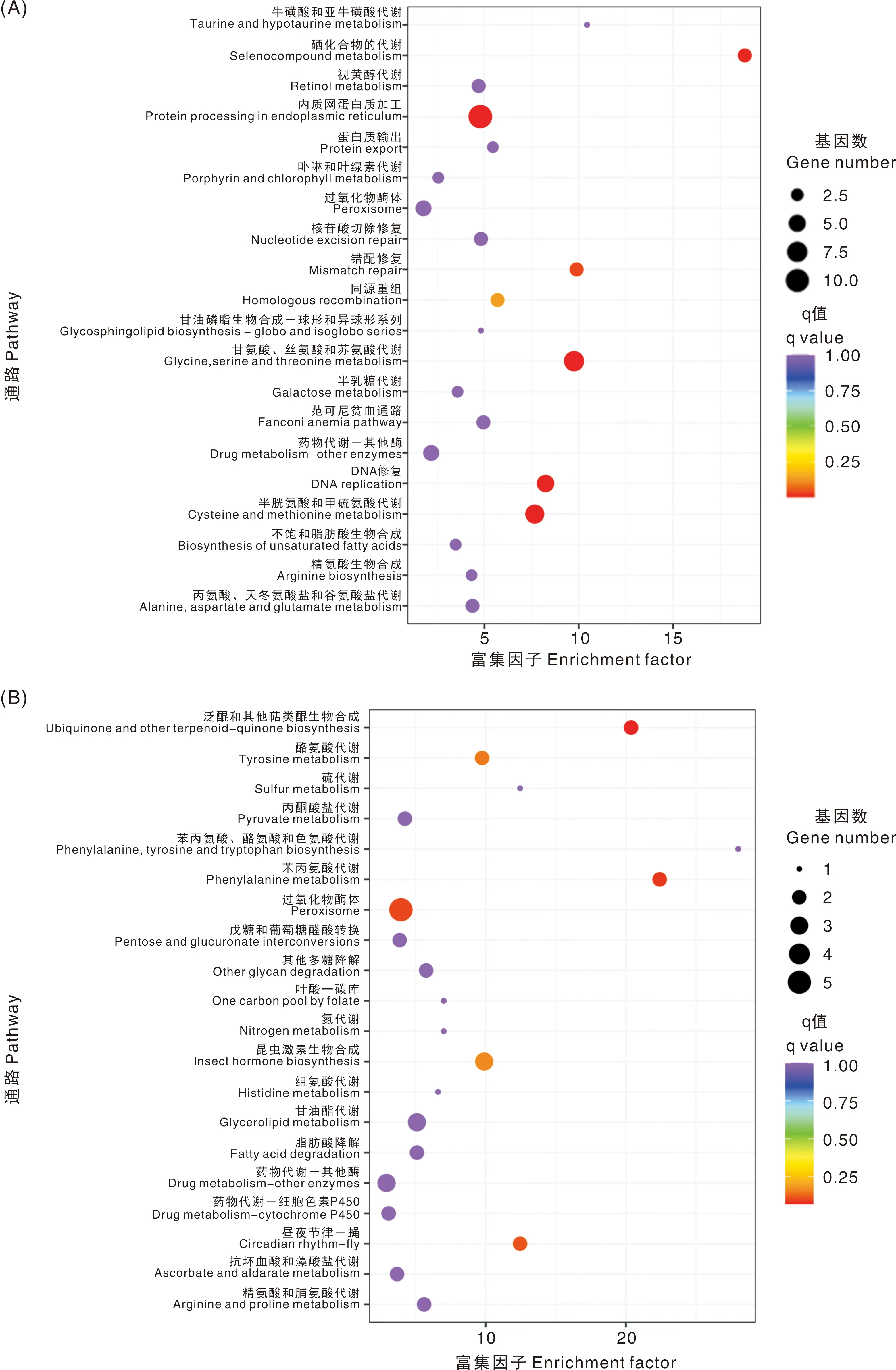
A, FAWw上调表达基因的KEGG代谢通路富集分析;B,FAWw下调表达基因的KEGG代谢通路富集分析。
3 讨论
本研究中,取食玉米的草地贪夜蛾幼虫转换饲喂小麦后的差异表达基因数量(542条)比转换饲喂人工饲料后的差异基因数量(6 863条)明显减少,表明小麦和玉米对草地贪夜蛾幼虫而言是非常相似的食物,可以互相替代。已有多项研究表明,草地贪夜蛾取食小麦后能顺利完成生活史。王芹芹等试验结果表明,草地贪夜蛾幼虫取食小麦和玉米的取食选择率、存活率、体重无显著差异。巴吐西等证明,草地贪夜蛾取食小麦后幼虫存活率和净增殖率低于取食玉米,但平均世代周期、内禀增长率、周限增长率无显著差异。吕亮等发现,草地贪夜蛾幼虫对小麦的食物利用效率低于玉米,但幼虫取食小麦生长发育速度比取食玉米更快。我国云南、安徽、江苏、河南、山东、四川均有草地贪夜蛾为害小麦田。因此,草地贪夜蛾取食小麦越冬的风险依然存在。
草地贪夜蛾是典型的多食性害虫,但是在取食上也有一定的偏好性。草地贪夜蛾有“玉米型”和“水稻型”两个亚群,“玉米型”草地贪夜蛾取食水稻后适合度显著降低。对“玉米型”和“水稻型”草地贪夜蛾幼虫进行基因组测序,发现“玉米型”和“水稻型”在细胞色素P450基因家族有显著分化,且“水稻型”的CYP4家族基因相比“玉米型”急剧扩张,表明细胞色素P450在草地贪夜蛾寄主适应性中起到重要作用。人工饲料里的有毒次生代谢物少,取食人工饲料后,草地贪夜蛾幼虫细胞色素P450家族基因显著下调,表明“适口性”更好的食物,可以减轻幼虫的解毒压力。而取食小麦后,细胞色素P450家族部分基因表达量有所下调,可能表明小麦对草地贪夜蛾幼虫而言“适口性”较好,也存在小麦的防御反应抑制了幼虫解毒能力的可能性。
本研究中还发现草地贪夜蛾在取食小麦之后,大量涉及免疫功能的基因,以及表皮相关的基因,表达量显著上调。而这一现象在取食人工饲料后没有发生。表明取食小麦可以提升草地贪夜蛾抵抗逆境的能力。尤其是大量表皮相关基因,在取食小麦后表达上调,而在取食人工饲料后表达下调,这个对比结果进一步说明了取食小麦对草地贪夜蛾的表皮生成有潜在的促进作用。这些差异表达的表皮相关基因主要涉及表皮蛋白基因。表皮蛋白对昆虫的表皮结构形成和稳固有关键作用,如在褐飞虱()中,一些表皮蛋白的缺失会造成表皮无法正常生成。昆虫在受到低温、干燥、农药等不利因素的影响时,表皮蛋白基因的表达量会上调。取食也会改变昆虫表皮蛋白基因的表达情况,如冈比亚按蚊()在血餐饲喂后,多种表皮蛋白基因的表达量发生变化。然而在草地贪夜蛾中,取食小麦对幼虫表皮的具体作用还需要进一步的实验验证,如取食小麦后表皮是否有增厚,是否能增强草地贪夜蛾的抗寒能力等。
综上所述,小麦是草地贪夜蛾幼虫较好的替代性寄主,秋冬季节玉米收割之后,草地贪夜蛾选择小麦作为越冬寄主的可能性很大。我国南方气候温暖,多数地区种植秋玉米和冬小麦。以温州为例,温州地区近年来的12月至次年2月平均气温均在10 ℃左右(温州气象局,http://www.wz121.com/menuDetail),2019年度种植玉米面积6 097 hm,小麦面积602 hm(温州市统计局,http://wztjj.wenzhou.gov.cn/art/2020/11/11/art_1468706_58725761.html),是适宜草地贪夜蛾越冬的地带。一些零星调查显示,温州地区在1月仍能够在玉米田边的禾本科杂草上以及残留的玉米株上发现4~6龄的草地贪夜蛾幼虫。需加强冬季小麦田的草地贪夜蛾幼虫及蛹的普查,尤其是前茬种植玉米的小麦田。
