增温及秸秆施用对大豆-冬小麦轮作农田土壤真菌群落组成及多样性的影响
2022-06-25张苗苗陈书涛丁司丞王瑾章堃
张苗苗 ,陈书涛 *,丁司丞,王瑾,章堃
1.南京信息工程大学江苏省农业气象重点实验室,江苏 南京 210044;2.南京信息工程大学应用气象学院,江苏 南京 210044
全球变暖是人们广泛关注的全球性环境问题,全球变暖对生态系统具有一系列影响(Ostberg et al.,2013)。农田生态系统不仅受人类农业管理措施影响,而且对全球变暖也会产生响应。在全球变暖背景下,作物生长条件的变化会影响土壤过程,而种植不同作物的农田土壤对变暖会产生不同的响应规律(Wall et al.,2011;Qin et al.,2013)。
人们以往通过增温试验研究变暖对农田生态系统的影响,而增温对农田生态系统的影响可能会对土壤微生物产生影响。温度是影响包括真菌在内的土壤微生物的重要环境因素(Castro et al.,2010),温度升高一方面直接影响微生物活性和类群(DeAngelis et al.,2015),另一方面通过影响植物生长进而影响微生物(Morrison et al.,2019)。培养实验研究表明,增温增加了赤壳科(Nectriaceae)真菌的相对丰度,但减少了被孢霉科(Mortierellaceae)和毛球壳科(Lasiosphaeriaceae)真菌的相对丰度(Habtewold et al.,2021)。以往研究表明苔原土壤真菌多样性对增温表现为正反馈响应模式(Feng et al.,2020)。不同生态系统土壤真菌群落结构对增温的响应模式存在差异,特定的植被和土壤条件影响土壤真菌群落结构对增温的响应程度(Wang et al.,2022),然而以往关于增温如何影响农田土壤真菌群落组成及多样性的研究还相对较少,以往的研究也主要集中在草地、苔原、森林等自然生态系统(Schindlbacher et al.,2011;Streit et al.,2014;Johnston et al.,2019)。
农田生态系统中,秸秆施用是常见的农业管理措施,秸秆含有的营养物质为微生物生存提供了物质条件。以往关于秸秆施用对真菌群落影响的研究结果存在差异,有研究表明,秸秆施用增加了稻田土壤真菌丰度,但对真菌α多样性无显著影响(Yuan et al.,2013);Wang et al.(2021)也报道秸秆施用对玉米田耕层土壤真菌α多样性无显著影响;Yang et al.(2020)发现秸秆施用降低了稻麦轮作农田土壤优势真菌目格孢腔菌目(Pleosporales)相对丰度,并使土壤真菌物种数和Shannon指数增大;Huang et al.(2019)研究表明,添加秸秆降低了玉米田土壤优势真菌门子囊菌门(Ascomycota)的相对丰度,并降低了真菌Chao1指数。在施用秸秆条件下,增温对农田土壤真菌的影响规律可能会比单因素的影响规律更为复杂,关于秸秆施用会增强还是减弱增温影响土壤真菌群落组成及多样性的效应这一问题还未能研究明确。
本研究设置了增温及秸秆施用田间试验,采用高通量测序对不同增温及秸秆施用处理下大豆(GlycinemaxL.)-冬小麦 (TriticumaestivumL.) 田土壤真菌群落组成及多样性进行分析,研究增温及秸秆施用如何影响大豆、冬小麦田土壤真菌门、纲、目水平上的组成及 α、β多样性,以期为探讨全球变暖和农业管理措施改变条件下农田土壤真菌群落的变异规律提供基础数据和理论支撑。
1 材料与方法
1.1 试验地点
田间试验地点位于南京信息工程大学农业气象试验站(32°12'N,118°42'E)。试验地为北亚热带季风气候,多年平均温度 15.4 ℃,多年平均降水量1090.6 mm,多年平均日照时数1902.5 h,无霜期237 d。土壤类型为黄棕壤,质地为壤质粘土。耕层(0—20 cm)土壤 pH 为 6.30,容重 1.54 g·cm−3,田间持水量25.6%,土壤有机质15.36 g·kg−1,全氮0.81 g·kg−1。
1.2 田间试验处理
增温及秸秆施用的长期试验开始于2014年11月(陈书涛等,2016),试验地采用大豆-冬小麦轮作。试验地设置3个区组(3个重复,分别以1、2、3标识),每个区组设置4个处理,分别为对照、增温、秸秆施用、增温及秸秆施用,处理代码分别为CK、WA、SA、WS,共12个小区,每个小区面积2.5 m×2.5 m。采用红外辐射加热管进行增温,这种增温方式为主动式增温,可同时改变显热和潜热(Bridgham et al.,1999;Luo et al.,2001)。在每个增温小区设置4根红外辐射加热管,每根加热管功率为250 W,加热管均匀分布于小区中,加热管悬挂于距离土壤50 cm(略低于作物最高冠层高度)的小区上方,每根加热管上方覆盖1个不锈钢挡雨板,挡雨板边缘向下倾斜。红外辐射加热管对每个增温小区以及增温及秸秆施用小区进行昼夜不间断加热。
向每个秸秆施用小区以及增温及秸秆施用小区分别添加2.86 kg秸秆,在大豆生长季施用冬小麦秸秆,在冬小麦生长季施用大豆秸秆。添加等量秸秆一方面是为了比较仅添加秸秆的 SA处理与CK处理土壤真菌群落组成的差异,并分析增温及秸秆施用对土壤真菌群落组成的复合影响规律;另一方面是为验证在不同生长季添加等量秸秆对土壤真菌群落组成是否会产生不同影响。将秸秆切成10 cm长小段,于翻耕前均匀施入每个小区。
大豆播种日期为2017年5月27日,于2017年10月9日收获;冬小麦播种日期为2017年10月28日,于2018年5月16日收获。在2017年6月—2018年5月期间(本研究中的大豆-冬小麦轮作年),CK、WA、SA、WS处理的年平均土壤温度分别为 (24.53± 0.04)、(25.58±0.04)、(24.66±0.04)、(25.78± 0.03) ℃。在每个大豆和冬小麦生长季之前以复合肥形式向各个小区施加 7.7 g·m−2N,7.7 g·m−2P2O5,7.7 g·m−2K2O,并在冬小麦生长季的 1月以尿素形式施用越冬肥,越冬肥为24.7 g·m−2N。
1.3 土壤样品采集及高通量测序
在 2017—2018大豆-冬小麦轮作年采集 0—20 cm土壤样品,在大豆生长中后期(2017年9月23日)采集大豆田土壤样品,在冬小麦生长中后期(2018年4月21日)采集冬小麦田土壤样品。在作物生长中后期采集土壤样品的原因为此时增温及秸秆施用的处理效应已经有一定时间的积累且作物此时仍具有较强生命活动。在各个取样小区以对角线法取样,将土钻插入20 cm深土层中,取出的样品进行混合,按四分法弃去多余样品,留取50 g样品装入塑料无菌自封袋。以高通量测序分析采集土壤样品的真菌群落组成。
利用 Illumina HiSeq测序平台进行土壤真菌ITS测序,构建小片段文库进行双末端测序。ITS(内转录间隔区)包括ITS1和ITS2,分别位于真核生物核糖体rDNA序列18S和5.8S之间及5.8S和28S之间。土壤样品进行冷冻研磨,以E.Z.N.A.®DNA 提取试剂盒(Omega Biotek,Norcross,GA,USA)提取土壤DNA,引物为ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和 ITS1R(5′-GCTGCGTTCTTCATCGATGC-3′),检测 DNA浓度及总量是否合格。提取合格的真菌基因以PCR(实时荧光定量聚合酶链式反应)进行扩增(95 ℃下反应5 min,95 ℃下反应30 s并进行27个循环,55 ℃下反应30 s,72 ℃下反应45 s,最后在72 ℃下扩增 10 min)(Rinke et al.,2013)。下机数据进行预处理,以 QIIME(微生物生态学定量研究)(Lozupone et al.,2011)进行质量控制,并进行OTUs(操作分类单元)聚类和注释,在过滤和筛查后以97%相似临界点进行 OUTs聚类(Edgar,2013),从而获得环境样品中真菌的物种组成及丰度信息,进一步进行多样性分析。
1.4 数据分析
对每个处理下 3个重复小区门水平上排名前10位土壤真菌类群相对丰度求平均值,并比较纲、目水平上相对丰度排前16位土壤真菌类群,应用Excel 2016中的柱状图表征4个处理的真菌相对丰度,以SPSS 19.0软件的单因素方差分析和多重比较分析不同处理下优势真菌门、纲、目的差异显著性。对于门水平上的微生物分类而言,一般选取相对丰度排前10位左右的类群(Wang et al.,2021;Cao et al.,2022),排在其后的微生物类群相对丰度一般很低。对于纲、目水平上的微生物分类而言,一般选取相对丰度较高的15—20种类群(Zhou et al.,2021;Ji et al.,2022),本研究中第 16种真菌纲、目相对丰度已低于1%,因而排在其后的真菌在图中不再表征。以箱线图分析大豆田和冬小麦田土壤不同处理下的真菌 α多样性指数(覆盖度、物种数、Shannon指数、Simpson指数、Chao1指数),采用一元回归分析冬小麦田土壤真菌α多样性指数与大豆田真菌 α多样性指数之间的关系。以热图和加权单比例(Weighted Unifrac)距离分析大豆田和冬小麦田土壤β多样性(即不同处理间物种的差异程度),若两处理距离越大,则物种差异性越大。
2 结果
2.1 真菌群落门、纲、目水平上的组成
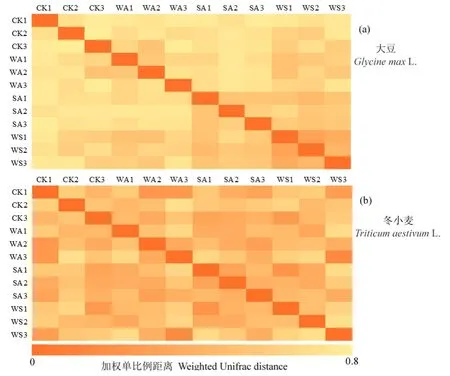
在门水平上,大豆田相对丰度最高的前10种土壤真菌门分别为子囊菌门、担子菌门(Basidiomycota)、接合菌门(Zygomycota)、纤毛亚门(Ciliophora)、壶菌门(Chytridiomycota)、球囊菌门(Glomeromycota)、类原生动物门(Rozellomycota)、绿藻门(Chlorophyta)、丝足虫门(Cercozoa)、芽枝霉门(Blastocladiomycota)(图 1a),这10种真菌门在CK、WA、SA、WS处理中的总相对丰度分别为 (81.0%±2.8%)、(49.8%±8.6%)、(81.0%±1.4%)、(74.3%±3.4%)。WA处理子囊菌门相对丰度显著低于CK(P=0.023)及SA(P=0.007)处理,且与WS处理存在边缘显著差异(P=0.063,表1)。CK与 WS处理接合菌门相对丰度存在显著(P=0.038)差异。不同处理间壶菌门、球囊菌门、绿藻门、丝足虫门也存在显著(P<0.05)或边缘显著(0.05 在门、纲、目水平上,冬小麦田与大豆田均存一些类似的土壤真菌类群。冬小麦田相对丰度最高的真菌门、纲、目分别为子囊菌门、粪壳菌纲、炭角菌目,与大豆田相同,但在其余类别组成上既存在相同类群又存在不同类群(图2a—c)。冬小麦田相对丰度最高的10种真菌门在CK、WA、SA、WS处理中的总相对丰度分别为 (80.8%±2.8%)、(77.7%±2.2%)、(77.0%±2.1%)、(69.5%±5.1%)。冬小麦田相对丰度最高的16种真菌纲在CK、WA、SA、WS处理中的总相对丰度分别为(67.0%±3.3%)、(55.7%±3.9%)、(62.6%±3.3%)、(53.3%±3.7%)。冬小麦田相对丰度最高的16种真菌目在CK、WA、SA、WS处理中的总相对丰度分别为 (44.2%±4.5%)、(38.1%±7.6%)、(43.6%±4.7%)、(37.4%±5.4%)。冬小麦田相对丰度最高的真菌门(子囊菌门)的相对丰度高于大豆田,在各个处理中的占比均超过50%。如表2所示,冬小麦田不同处理油壶菌门、球囊菌门、油壶菌纲、炭角菌目、粪壳菌目之间存在显著差异(P<0.05)或边缘显著差异(0.05 表2 冬小麦田存在显著差异的真菌门、纲、目相对丰度Table 2 Relative abundance of soil fungi at the phylum, class and order levels in the winter wheat crop field % 图2 增温及秸秆施用对冬小麦田土壤真菌门、纲、目相对丰度的影响Figure 2 Effects of warming and straw application on the relative abundance of soil fungi at the phylum, class and order levels in the winter wheat crop field 测序结果表明大豆田不同处理的不同α多样性指标之间的差异不同。各处理之间真菌群落覆盖度无显著差异(P>0.10,图3a)。4个处理中WA处理的物种数显著小于 WS处理(P=0.012),CK处理的物种数与WS处理存在边缘显著差异(P=0.052,图 3b)。SA(P=0.032)和 WS(P=0.014)处理 Shannon指数均显著高于WA处理(图3c),WA处理与SA(P=0.063)和WS(P=0.061)处理Simpson指数均存在边缘显著差异(图3d),各处理之间Chao1指数无显著差异(P>0.10,图 3e)。总体而言,大豆田WA处理物种数最少,且具有最低的多样性。 图3 不同增温及秸秆施用处理下大豆田土壤真菌α多样性箱线图Figure 3 Box and whisker plot of the α diversity of soil fungi under the different warming and straw application treatments in the soybean crop field 冬小麦田各处理真菌群落覆盖度无显著差异(P>0.10,图4a)。CK与SA处理真菌物种数存在边缘显著差异(P=0.087),WA与WS处理物种数也存在边缘显著差异(P=0.089,图4b)。WA和SA处理Shannon指数存在边缘显著差异(P=0.088,图4c)。各处理Simpson指数之间无显著差异(P>0.10,图4d)。与物种数相似,CK与SA处理Chao1指数存在边缘显著差异(P=0.088,图4e),WA与WS处理Chao1指数也存在边缘显著差异(P= 0.091,图4e)。与大豆田类似,冬小麦田WA处理物种数相对最少,且具有最低的多样性。冬小麦田SA处理比CK处理具有更多的物种数和更大的多样性。大豆田土壤真菌覆盖度低于冬小麦田(图 5a),物种数、Shannon指数、Simpson指数、Chao1指数均高于冬小麦田(图5b—e)。一元回归分析表明,冬小麦田与大豆田真菌物种数之间存在显著自然对数回归关系(P=0.047),冬小麦田与大豆田Shannon指数之间存在边缘显著线性回归关系(P=0.057),表明冬小麦田与大豆田各处理之间物种数及Shannon指数的相对增减规律类似,在大豆生长季具有较高物种数和 Shannon指数的处理在冬小麦生长季两个指标的数值也相对较高。从冬小麦田和大豆田物种数及 Shannon指数具有正相关性的规律来看,在大豆和冬小麦田增温及秸秆施用对土壤真菌群落物种数和Shannon指数均有类似的影响规律。 图4 不同增温及秸秆施用处理下冬小麦田土壤真菌α多样性箱线图Figure 4 Box and whisker plot of the α diversity of soil fungi under the different warming and straw application treatments in the winter wheat crop field 图5 大豆田与冬小麦田土壤真菌α多样性指数之间的关系Figure 5 Relationships between the α diversity indexes in the soybean cropland and those in the winter wheat crop field 土壤真菌β多样性以加权单比例距离衡量,距离越大,则两处理间的物种差异性越大。图6a表明,大豆田CK1、CK2、CK3(1、2、3代表3个重复的编号)与SA3、SA3、SA3之间的距离相对较大,例如,CK1与SA3之间的加权单比例距离为0.782;WA1、WA2、WA3与WS1、WS2、WS3之间的加权单比例距离相对较小,表明两处理间差异相对较小。图6b表明,冬小麦田CK1、CK2、CK3与WS1、WS2、WS3之间的距离相对较大,表明两处理间的物种具有较大差异。图6a、b表明,大豆田土壤真菌β多样性总体上大于冬小麦田土壤,大豆田各处理间土壤真菌物种差异性相对更大。 图6 大豆田与冬小麦田土壤真菌加权单比例距离热图Figure 6 Heatmap of weighted Unifrac distance of soil fungi in the soybean and winter wheat crop fields 以往关于增温如何影响土壤真菌群落多样性的研究存在差异,例如王军等(2018)研究表明,短期增温对紫花针茅草原土壤真菌丰度(真菌数量)无显著影响,分子鉴定技术结果表明增温对土壤丛枝菌根[球囊菌门 (Glomeromycota)]多样性无显著影响(Jiang et al.,2017;Wei et al.,2021);李欣等(2017)研究发现,增温增加了土壤高寒草甸真菌数量;Rinnan et al.(2007,2009)研究表明,增温降低了土壤真菌丰度;Feng et al.(2009)也发现增温减少了土壤真菌数量。本研究中WA处理具有最低的物种数和α多样性,但差异未达显著水平。本研究以及以往关于增温影响土壤真菌多样性的研究表明,增温对土壤真菌的影响可能因植物类型、气候条件而不同(Wang et al.,2022)。在农田中增温对真菌群落多样性的影响规律不同于其他生态系统,这可能与种植作物的作用更明显有关,大豆田物种数、Shannon指数、Chao1指数均高于冬小麦田,大豆作物更强的供氮能力更有利于真菌群落的多样性。 秸秆施用增加了微生物生存所需的含碳底物(Chahal et al.,2018;Thangarajan et al.,2013),改善了土壤的养分条件(Bakht et al.,2009;Sun et al.,2020),有利于真菌等微生物的生存。然而,在种植不同作物的农田中,由于作物类型、水分管理方式、秸秆施用方式等不同,秸秆施用对真菌α多样性会产生不同的影响规律。有研究表明,秸秆施用降低了玉米田土壤真菌的α多样性(Huang et al.,2019;Zhang et al.,2021);但对水稻田土壤真菌α多样性无显著影响(Wang et al.,2021)。Wang et al.(2021)发现秸秆施用对玉米田耕层土壤真菌 α多样性无显著影响,但显著增加了深层土壤真菌α多样性。本研究表明,在施用秸秆条件下冬小麦田土壤真菌物种数和Chao1指数增加,且大豆田WS处理物种数高于CK,与Huang et al.(2019)和Zhang et al.(2021)的研究结果一致,说明在本研究中的大豆和冬小麦田秸秆施用为土壤真菌的生存提供的养分条件有利于增加真菌多样性。 在秸秆施用条件下,增温对土壤真菌群落多样性的影响规律变得更为复杂。大豆和冬小麦田WA处理土壤真菌物种数均最低,且具有最小的Shannon指数,但秸秆施用抵消了这种效应。图3、4还表明增温及秸秆施用对土壤真菌的影响具有交互效应,这种交互效应在大豆和冬小麦田均存在。不仅增温及秸秆施用对土壤真菌α多样性的影响存在生长季间的差异,而且不同生长季土壤真菌α多样性也不同。种植豆科作物的农田可向土壤中供给更多氮源,与冬小麦田相比,提供了更多营养源,因此,与α多样性类似,大豆田土壤真菌β多样性总体上大于冬小麦田土壤,大豆田各处理间土壤真菌物种差异性相对更大。 子囊菌门是真菌界中种类最多的一个门,也是具有代表性的真菌类群,姚世庭等(2021)研究表明增温与对照条件下排名前3的优势真菌门均为子囊菌门、担子菌门、接合菌门,这与本研究中的 3种优势真菌门完全相同。本研究中,增温及秸秆施用改变了土壤真菌群落组成,在门水平上,大豆田WA处理子囊菌门相对丰度显著低于 CK,秸秆施用部分抵消了增温对子囊菌门相对丰度的减少效应,而秸秆施用本身对优势门的影响不显著,这与Zhang et al.(2021)的研究结果类似。与门水平上类似,秸秆施用抵消了增温对粪壳菌纲相对丰度的减少效应,秸秆施用还提高了散囊菌目的相对丰度。在冬小麦田,WS比CK也减少了子囊菌门相对丰度。在温度升高条件下,相对丰度最高的优势门和优势纲的真菌数量减少,表明增温导致真菌中优势门的减少。向田间施用的秸秆中包含了碳、氮等多种可供给真菌生存的营养物质,从而为优势门的生存提供了养分条件(Govaerts et al.,2007;Zhen et al.,2009),这与增温对优势门的胁迫效应相抵消。在大豆田,WA处理前10种相对丰度最高的真菌门和前 16种相对丰度最高的真菌纲、目的总相对丰度在4个处理中最少,而冬小麦田的这种规律不明显,这表明增温对真菌优势门、纲、目的影响因不同作物类型而异。冬小麦田种类最多的门(子囊菌门)的相对丰度高于大豆田,这可能与大豆田真菌物种数相对更多有关。此外,大豆田土壤真菌门、纲、目水平上存在差异的类群明显比冬小麦田多(表1、2),这说明大豆田真菌类群受增温及秸秆施用处理的影响相对更大。豆科作物根瘤具有固氮能力,对土壤具有自然供氮能力,大豆田土壤可为多种真菌类群的存在提供合适的场所以及营养条件(Strom et al.,2020;Guo et al.,2021),这其中的某些真菌类群易受增温及秸秆施用的影响。 (1)大豆田增温处理土壤真菌物种数最少,且具有最低的α多样性。冬小麦田秸秆施用处理比对照增加了土壤真菌物种数和α多样性。大豆田土壤真菌物种数和Chao1指数远高于冬小麦田,且具有更高的Shannon指数和Simpson指数。大豆田土壤真菌β多样性总体上大于冬小麦田,前者不同处理之间的物种差异性相对更大。 (2)增温及秸秆施用改变了大豆-冬小麦轮作农田的土壤真菌群落组成。在大豆田,增温降低了相对丰度最高的优势真菌门子囊菌门相对丰度;在纲水平上,增温降低了大豆田相对丰度最高的优势真菌纲粪壳菌纲相对丰度;在目水平上,秸秆施用提高了大豆田散囊菌目相对丰度。在冬小麦田,增温及秸秆施用降低了油壶菌门相对丰度,各处理优势真菌纲粪壳菌纲相对丰度之间无显著差异,增温及秸秆施用降低了炭角菌目相对丰度。 (3)大豆田自身的供氮作用使其具有更大的真菌物种数和α、β多样性,大豆田在不同增温及秸秆施用处理下存在差异的真菌门、纲、目类群也多于冬小麦田。种植大豆有利于改善土壤真菌群落结构。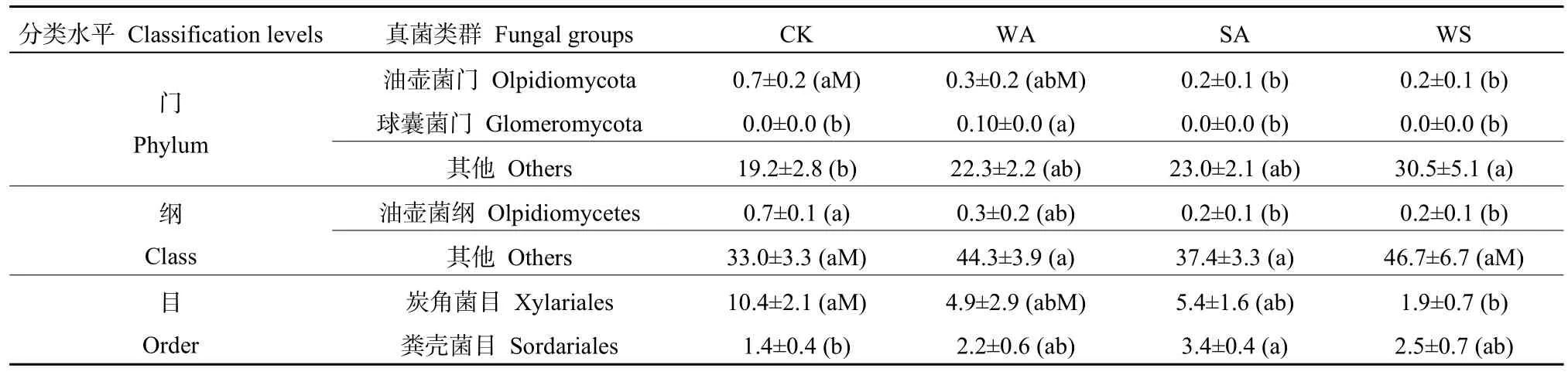
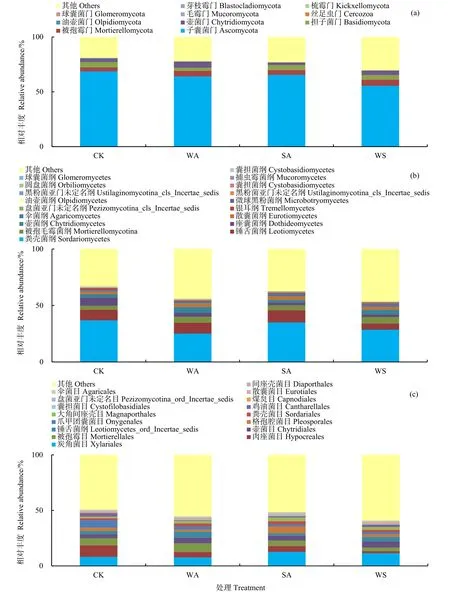
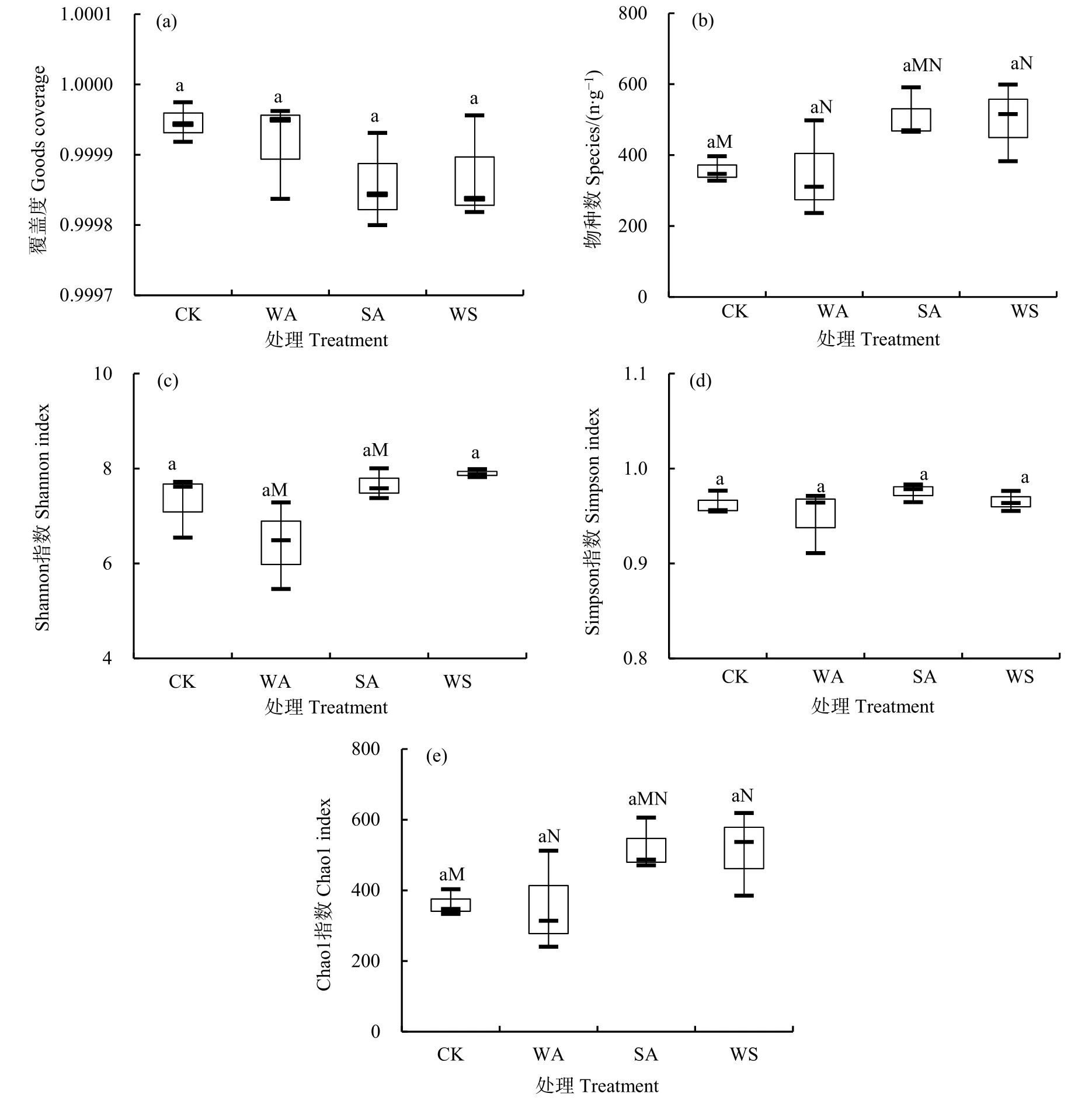
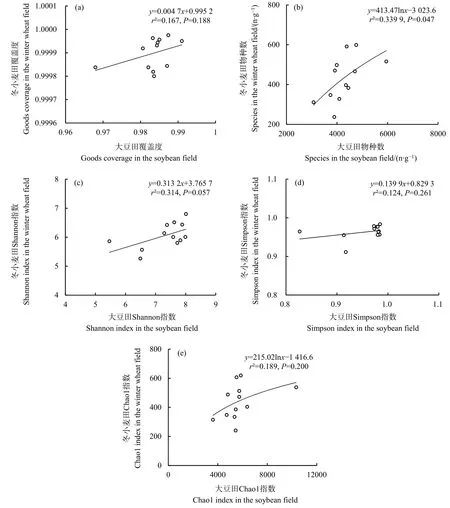
2.2 真菌群落α多样性

2.3 真菌群落β多样性
3 讨论
3.1 增温及秸秆施用对真菌群落多样性的影响
3.2 增温及秸秆施用对真菌群落组成的影响
4 结论
