沙利度胺对阿尔茨海默病秀丽隐杆线虫模型的改善作用及机制
2022-06-23方亚影阎茹玉李玉贤吴宿慧李寒冰李根林
方亚影 阎茹玉 李玉贤 吴宿慧 李寒冰 李根林
关键词沙利度胺;秀丽隐杆线虫;阿尔茨海默病;磷脂酰肌醇3-激酶/蛋白激酶B信号通路
阿尔茨海默病(Alzheimer’s disease,AD)是一种常见的渐行性中枢神经系统退行性疾病,患者大脑中普遍呈现脑组织萎缩、老年斑、神经元纤维缠结等病理特征[1]。全球AD患者正逐年递增,目前临床上也缺乏能有效治疗AD的药物[2]。
沙利度胺是一种新型的免疫调节药和抗炎药,可抑制单核细胞和巨噬细胞分泌肿瘤坏死因子α(tumornecrosis factor-α,TNF-α),从而減少淀粉样蛋白的形成;其还能抑制白细胞介素1β(interleukin-1β,IL-1β)和IL-6等炎症因子的产生[3]。相关研究发现,上述炎症因子可参与Tau 蛋白的过度磷酸化,加快Tau 蛋白病理形成的过程[4-5]。
Tau 蛋白的病理变化与AD患者认知能力下降有较强的相关性。Tau 蛋白的正常生理功能是与微管蛋白结合,以维持细胞骨架的稳定性[6]。糖原合成激酶3β(glycogensynthase kinase 3β,GSK-3β)是磷脂酰肌醇3-激酶(phosphoinositide3-kinase,PI3K)和蛋白激酶B(proteinkinase B,Akt)的下游底物,其活化后可导致Tau 蛋白过度磷酸化,从而使Tau 蛋白大量沉积并形成缠结,进而破坏神经元的生理功能[7 - 8]。相关研究发现,激活PI3K/Akt途径可改善神经元细胞凋亡[9]。
秀丽隐杆线虫(以下简称“线虫”)因其生命周期短、神经系统结构简单、遗传信息清晰和易于获得大量个体模型等优点,常被用于神经退行性疾病的相关研究[10]。
BR5270 品系线虫体内可表达人源Tau 蛋白,具有加速Tau 蛋白聚集的特点,使其从成年第1 天起就出现明显的神经元功能障碍;BR5271 品系线虫体内表现出抗Tau蛋白聚集的特点,常作为BR5270 品系线虫的对照[11]。
另外,相关研究发现,线虫体内PI3K和Akt 的同源基因是Age-1 和Akt-1[12-13]。基于此,本研究以BR5270 品系线虫为AD模型,研究沙利度胺改善线虫体内Tau 蛋白毒性及提高线虫学习记忆能力的作用及机制,以期为沙利度胺治疗AD提供实验依据。
1 材料
1.1 主要仪器
本研究所用主要仪器有HVE-50 型高压蒸汽灭菌锅(日本Hirayama 公司),DL-CJ-2ND Ⅰ型洁净工作台(北京东联哈尔仪器制造有限公司),LY03-200 型生化培养箱(上海龙跃仪器设备有限公司),Nano Drop One C2000 型超微量紫外分光光度计、Fresco 21 型低温离心机(美国Thermo Fisher Scientific 公司),Quant Studio 7Flex 型逆转录聚合酶链式反应(PCR)仪(美国AppliedBiosystems 公司),Gel Doc XR+型自动凝胶成像系统(美国Bio Rad公司)。
1.2 主要药品与试剂
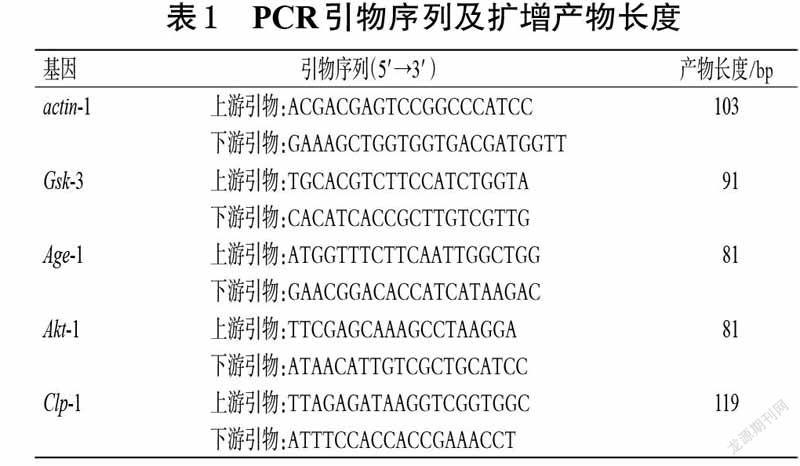
本研究所用主要药品与试剂有沙利度胺片(常州制药厂有限公司,批号19031332,规格50 mg/片),二甲基亚砜(DMSO,天津市风船化学试剂科技有限公司,批号20210415),蛋白胨、琼脂粉(北京奥博星生物技术有限公司,批号分别为20190126、20191030),酵母浸粉(美国Oxoid 公司,批号分别为1267856-03),Trizol 试剂(美国Ambion Life 公司,批号204202),TB GreenTM Premix ExTaqTM Ⅱ、PrimeScriptTM RT reagent Kit with gDNA Eraser(日本Takara 公司,批号分别为RR820A、RR047A)。Gsk-3、Age-1、Akt-1、Clp-1 基因(钙蛋白酶的同源基因)的引物由深圳华大基因科技有限公司设计并合成(PCR引物序列及扩增产物长度见表1)。其余试剂为实验室常用规格,水为纯净水。
1.3 动物
线虫(品系包括BR5270、BR5271)均购自美国线虫中心(https://cgc.umn.edu),大肠埃希菌Escherichia coliOP50 尿嘧啶合成缺陷菌株由北京生命科学研究所董梦秋教授课题组惠赠。
2 方法与结果
2.1 线虫的培养及同期化处理
2.1.1 NGM培养基的制备称取氯化钠0.60 g、蛋白胨0.50 g、琼脂粉3.40 g,加入5 mg/mL 胆固醇溶液200 μL和纯水200 mL,经高压蒸汽灭菌(121 ℃,15 min)后,依次加入浓度均为1 mol/L 的MgSO4溶液和CaCl2溶液各200 μL、1 mol/L 磷酸盐缓冲液(pH=6.0)5 mL,摇匀后倒板,加入100 μL 大肠埃希菌溶液(由于线虫以大肠埃希菌为食,故后续将其称为“食物”)涂布均匀,于37 ℃培养箱中培养8~12 h,备用。
2.1.2 线虫的同期化处理将2 种线虫(BR5270、BR5271 品系)置于含有食物的NGM 培养基上培养(16 ℃条件下)。挑取处于产卵期的线虫于含有食物的NGM培养基上,培养4 h 后,移除线虫,保留虫卵;继续孵育虫卵至产卵期,即得同期化的亲代线虫,再将亲代线虫继续培养至产卵期,保留虫卵,即得同时期的虫卵[14]。
2.2 沙利度胺对BR5270品系线虫的毒性预实验
取BR5270 品系线虫,按“2.1.2”项下方法进行同期化处理后,分为溶剂对照组(1%DMSO)和沙利度胺不同质量浓度组(10、25、50、100 mg/mL),每组20 条,并设个平行。将线虫置于含不同药物的NGM培养基(含食物)上,于16 ℃条件下恒温培养24 h 后,记录各组线虫的死亡数量。结果显示,溶剂对照组和沙利度胺10mg/mL组线虫无死亡;当沙利度胺给药质量浓度增加至25 mg/mL 时开始有线虫死亡,死亡数为(1.67±0.58)条;沙利度胺给药质量浓度为50、100 mg/mL时,线虫死亡数分别为(5.00±1.00)、(17.34±2.52)条。这提示沙利度胺对线虫的毒性随着质量浓度的增加而增强。因此,为了避免线虫毒理学特征的出现,本研究将沙利度胺最大给药质量浓度定为15 mg/mL。
2.3 BR5270 品系线虫的基础慢反应实验
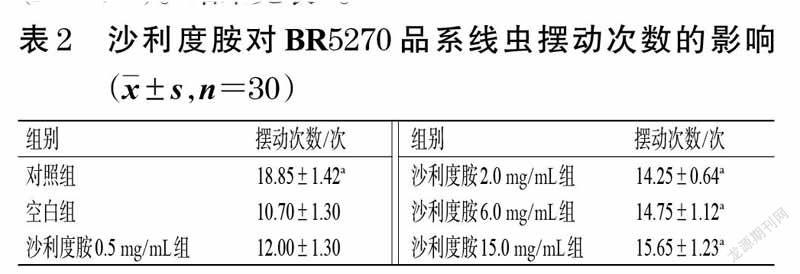
BR5270 品系线虫由于Tau 蛋白的聚集,表现为从成年第1 天开始就运动不协调、运动神经元轴突缺损[11],因此,通过测定线虫的摆动次数,可探究沙利度胺对BR5270 品系线虫运动能力的影响。按“2.1.2”项下方法将BR5271、BR5270 品系线虫进行同期化处理,然后于16 ℃条件下培养24 h 后,将BR5271 品系线虫作为对照组,BR5270 品系线虫分为空白组(不加药物)和沙利度胺不同质量浓度组(0.5、2.0、6.0、15.0 mg/mL),每組30条,并设3 个平行。将线虫置于含不同药物的NGM培养基(含食物)上,于16 ℃条件下培养至成年(培养5 d)后,测定各组线虫30 s 内的摆动次数[15]。采用GraphPadPrism 8.0.2 统计软件作图,以SPSS 24.0 统计软件分析实验数据,结果用x±s 表示;多组间比较采用单因素方差分析,组间两两比较采用LSD-t 检验(方差齐时)或Dunnett’s T3 多重比较(方差不齐),检验水平α=0.05(下同)。结果显示,与空白组比较,对照组与沙利度胺2.0、6.0、15.0 mg/mL组线虫在30 s 内的摆动次数均显著增加(P<0.01)。结果见表2。
2.4 BR5270 品系线虫的寿命实验
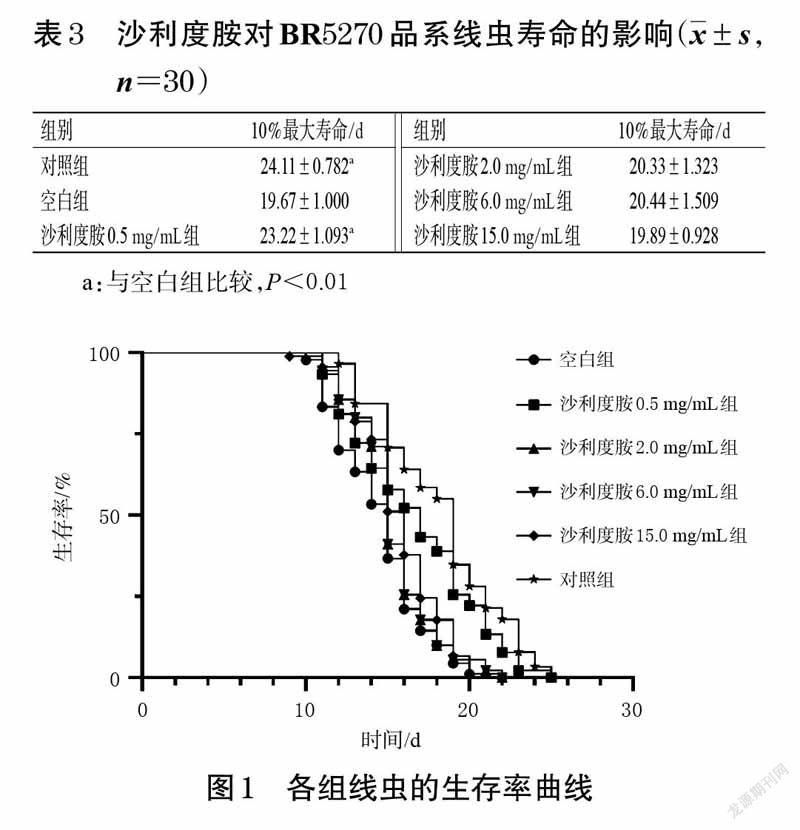
BR5270 品系线虫由于Tau 蛋白聚集,其生存时间比BR5271 品系线虫短[11],10%最大寿命可反映药物对线虫生存时间的影响[16],因此,通过分析线虫的10%最大寿命和生存率曲线,可探讨沙利度胺是否能延长BR5270品系线虫的寿命。按“2.3”项下方法将BR5271、BR5270品系线虫进行同期化处理及分组、给药,并设3 个平行,于16 ℃条件下进行培养。每隔24 h 观察线虫的生存状态,并记录线虫的死亡数量,直至各组线虫全部死亡,计算各组线虫10%最大寿命[15]。线虫死亡的判断标准:将线虫拾取器用酒精灯烧热后,轻触线虫并观察线虫是否有反应,如果没有反应则判断为死亡[16]。结果显示,与空白组比较,对照组和沙利度胺0.5 mg/mL 组线虫的10%最大寿命显著延长(P<0.01),且两者的生存率曲线也整体高于空白组;沙利度胺其余各组的10%最大寿命差异无统计学意义。后续实验的给药周期比寿命实验短,故将沙利度胺给药质量浓度设置为0.5、2.0、6.0mg/mL。结果见表3、图1。
2.5 BR5270 品系线虫的短期学习记忆能力实验
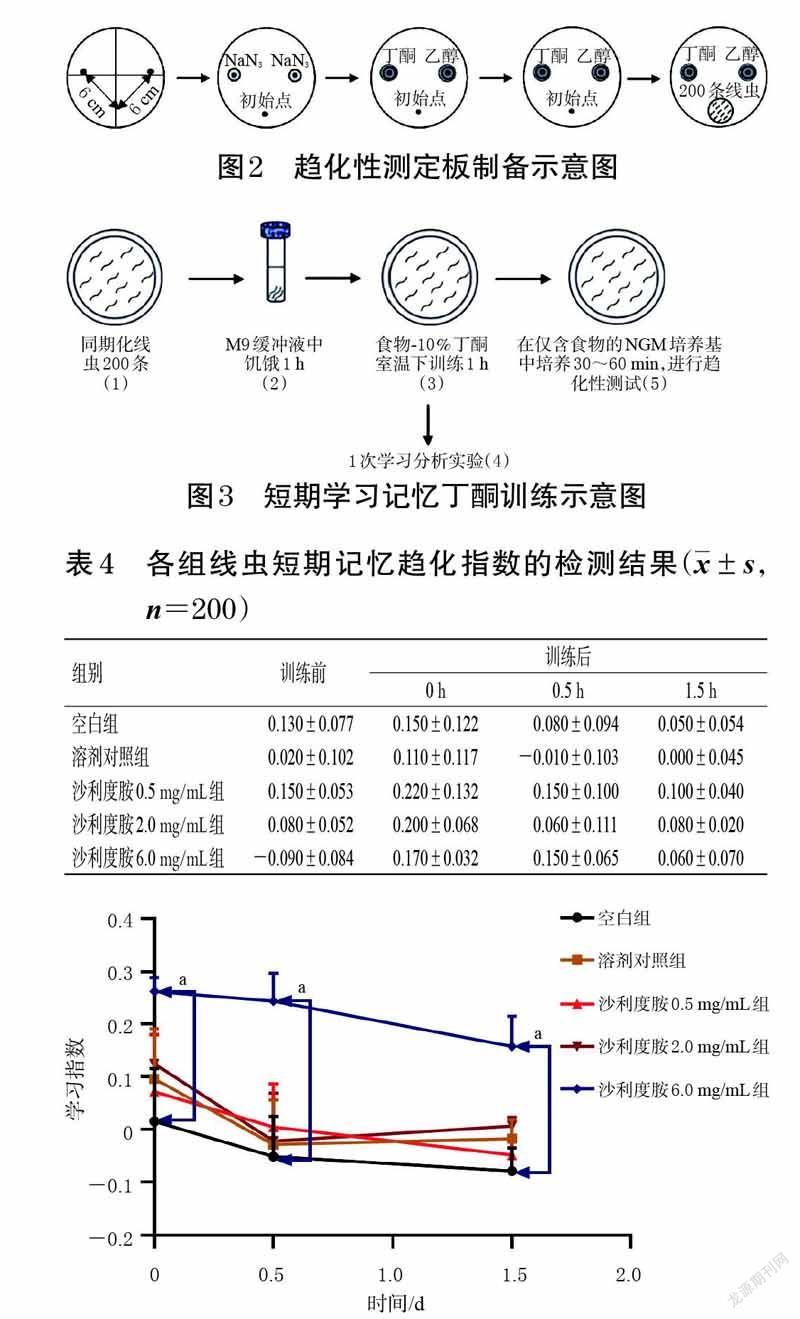
BR5270 品系线虫的学习指数与短期学习记忆能力呈正相关,因此,通过测定各组BR5270 品系线虫的学习指数,可探究沙利度胺是否能改善BR5270 品系线虫的短期学习记忆能力[17]。按“2.1.2”项下方法将BR5270 品系线虫进行同期化处理,然后将虫卵于16 ℃培养4 d后,分为空白组、溶剂对照组(1%DMSO)、沙利度胺不同质量浓度组(0.5、2.0、6.0 mg/mL),每组800 条,继续培养48 h 后,各组取200 条线虫置于不含食物的NGM培养基上进行趋化性测试(趋化性测定板制备如图2 所示),记录每组线虫训练前的趋化指数(CINaive)[17]。将各组剩余线虫进行短期学习记忆丁酮训练(如图3 所示),再转移至含有食物的NGM固体培养基上进行培养,分别于培养0、0.5、1.5 h 时,每组各取200 条线虫进行趋化性测试,并记录各时间点的趋化指数(CI0 h、CI0.5 h、CI1.5 h),再计算各时间点的学习指数[学习指数=CI0 h-CINaive(或CI0.5 h-CINaive,或CI1.5 h-CINaive)],实验重复3 次[18]。结果显示,与空白组比较,沙利度胺6.0 mg/mL 组线虫的短期学习指数在培养0、0.5、1.5 h 时均显著升高(P<0.01);其余给药组该指数差异无统计学意义,详见表4、图4。
2.6 BR5270 品系线虫长期学习记忆能力实验
将BR5270 品系线虫按“2.5”项下方法进行同期化处理和分组(每组1 000 条),培养48 h 后,各组取200 条线虫置于不含食物的NGM培养基上进行趋化性测试(每组平行3 个平板),记录每组线虫训练前的趋化指数(CINaive)。将各组剩余线虫进行长期学习记忆能力丁酮训练(如图5 所示)[17]。最后一次训练完成后,将线虫转移至含食物的NGM培养基上进行培养,分别于培养0、16、24、40 h 时,每组取200 条线虫进行趋化性测试,并记录各时间点的趋化指数(CI0 h、CI16 h、CI24 h、CI40 h),再按“2.5”项下公式计算各时间点的学习指数。结果显示,与空白组比较,沙利度胺6.0 mg/mL 组线虫的长期学习指数在培养0、16 h 时均显著升高(P<0.01);其余给药组该指数差异无统计学意义,详见表5、图6。
2.7 BR5270 品系线虫体内Gsk-3、Age-1、Akt-1、Clp-1基因mRNA的表达水平检测
将BR5270 品系线虫按“2.5”项下方法进行同期化处理和分组,继续培养48 h 后,采用低温Trizol 法提取各组线虫的总RNA,再按相应试剂盒说明书方法进行反转录得到cDNA。以cDNA 为模板,进行实时荧光定量PCR 反应。反应体系为:cDNA 模板1 μL,TB GreenTMPremix Ex TaqTM Ⅱ 10 μL,ROX Reference DyeⅡ 0.4 μL,上、下游引物各0.8 μL,RNase free water 7 μL。反应条件为:95 ℃变性30 s;95 ℃变性15 s,52 ℃退火30 s,72 ℃延伸30 s,共40 个循环。以actin-1 基因为内参,采用2-ΔΔCt法计算Gsk-3、Age-1、Akt-1、Clp-1 基因mRNA的表达水平。实验重复3 次。结果显示,与空白组比较,沙利度胺各质量浓度组线虫体内Age-1、Akt-1 基因(沙利度胺0.5、2.0 mg/mL 组除外)的mRNA表达水平均显著升高(P<0.05 或P<0.01),Gsk-3(沙利度胺0.5 mg/mL组除外)、Clp-1 基因的mRNA表达水平均显著降低(P<0.05 或P<0.01),结果见表6。
3 讨论
本研究使用的BR5270 品系线虫属于Tau 转基因线虫模型,人源Tau 蛋白聚集片段由于K280 基因的缺失增强了Tau 蛋白的聚集倾向,使该品系线虫的毒性急剧增加,表现为从成年第1 天开始就运动不协调、轴突缺损、运动能力严重受损等[11]。BR5271 品系线虫由于具有抗Tau 蛋白聚集的特点,常作为BR5270 品系线虫的对照组[11]。本研究的基础慢反应实验结果显示,沙利度胺可以明显增加BR5270 品系线虫的摆动次数,这提示其可改善线虫因Tau 蛋白聚集而导致的运动能力受损。寿命实验结果显示,低剂量(0.5 mg/mL)的沙利度胺可显著延长BR5270 品系线虫的10%最大寿命,这提示沙利度胺能较好地缓解该品系线虫因Tau 蛋白聚集而导致的毒性作用。
丁酮是一种带有气味的挥发性物质,在缺乏食物时,给予线虫食物并将其暴露于丁酮气味中,线虫会根据丁酮气味寻找食物,因此可将其用来研究线虫的学习记忆能力[18]。一般情况下,对线虫进行长期记忆训练的持续时间应大于16 h,短期记忆训练的持续时间则应小于2 h[19-20]。本研究结果显示,经沙利度胺(6.0 mg/mL)干预后,BR5270 品系线虫短期学习指数和长期学习指数(培养0、16 h 时)均显著升高,长期学习指数(24、40 h)与空白组相比有升高,但无显著性差异,这可能与长期学习记忆能力从培养16 h 后开始下降有关,与Amano等[20]的研究结论一致。这提示沙利度胺可改善BR5270品系线虫短期和长期的学习记忆能力。
过度磷酸化的Tau 蛋白与微管脱离后,可积累形成神经元纤维缠结,从而造成突触结构的可塑性损伤以及神经元的变性和凋亡,进而引发AD[21]。GSK-3β是诱导Tau 蛋白磷酸化的主要蛋白激酶,其在线虫体内的同源基因为Gsk-3[22]。在PI3K/Akt 信号通路中,PI3K(线虫体内的同源基因为Age-1)被激活后可活化Akt(线虫体内的同源基因为Akt-1)的功能,活化后的Akt 又可抑制GSK-3β的活性,进而减少Tau 蛋白的异常磷酸化[23]。本研究结果显示,沙利度胺可显著升高BR5270 品系线虫体内Age-1、Akt-1 基因(0.5、2.0 mg/mL 的沙利度胺除外)mRNA表达水平,显著降低Gsk-3 基因mRNA表达水平(0.5 mg/mL 的沙利度胺除外),这提示沙利度胺可能是通过活化PI3K/Akt 信号通路来减少Tau 蛋白的异常磷酸化。另外有研究发现,Tau 蛋白与突触后致密蛋白95(postsynaptic density-95,PSD-95)相互作用后,可使Ca2 +通过N-甲基-D-天冬氨酸(N-methyl-D-aspartic acidreceptor,NMDA)受体大量流入细胞,从而激活钙蛋白酶并引发线粒体功能障碍,最终诱导神经细胞凋亡[24]。在线虫体内钙蛋白酶的同源基因是Clp-1[25]。本研究结果显示,沙利度胺可显著降低BR5270 品系线虫体内Clp-1 基因mRNA表达水平,这提示沙利度胺可能是通过抑制钙蛋白酶来减轻Tau蛋白诱导的神经细胞凋亡。
综上所述,沙利度胺可改善AD線虫模型的运动障碍,并延长其寿命,增强其学习记忆能力;具体作用机制可能与激活PI3K/Akt 信号通路和抑制蛋白酶有关。本研究结果也初步证实沙利度胺抗AD的作用,可为沙利度胺治疗AD提供实验依据。
