PRL 和O2 对HC11 细胞氧化应激状态下乳蛋白合成的影响
2022-06-20AlexanderJonathanSpitzerZHAOFengqi
田 青 , Alexander Jonathan Spitzer, ZHAO Fengqi
(1.江苏食品药品职业技术学院,江苏淮安223003;2.佛蒙特大学,伯灵顿05405)
氧化应激是促氧化物和抗氧化物之间的不平衡所致。Saeidnia 和Abdollahi(2013)研究表明,奶牛围产期间随着代谢需求增加, 尤其是氧气需求的增加会导致活性氧(ROS)产量的增加,尤其是奶牛乳腺,作为乳汁合成的主要部位,其所承担的代谢压力更易发生氧化应激。 研究表明, 持续的氧化应激将诱导脂质过氧化, 继而引起线粒体损伤,造成细胞凋亡,降低机体免疫功能和炎症应答能力 (Ma 等,2018;Sordillolm 等,2005)。 Ma 等(2019)研究表明,氧化应激对免疫系统和炎性应答的影响可使奶牛对疾病的易感性增加,酮血症、脂肪肝、卵巢疾病、低血钙、低血镁等患病率随之升高。 研究表明, 奶牛分娩后乳腺代谢活跃而使高产奶牛乳腺产生大量的活性氧(ROS),而过量的ROS 会诱导隐性乳房炎的发生(Ma 等,2018;Mulligan 等,2008)。 同时,Ma 等(2018)研究还发现, 乳腺的密集生长和牛奶的合成也是乳腺产生氧化应激和炎症的原因。 促炎作用的发挥则主要通过有丝分裂原激活蛋白激酶(MAPK)、丝氨酸/苏氨酸激酶Akt (AKT) 和核因子kappa B (NFκB)介导,即过量的ROS 首先激活MAPK 和AKT信号通路,随后NF-κβ 通路被激活,继而引发炎症因子的高表达, 炎症反应程度和进程加重和延长,乳腺细胞开始受损伤,细胞开始凋亡,乳蛋白基因表达受阻, 乳品质变差, 甚至停乳(Abuelo等,2015;Liu 等,2012;Cargnello 等,2011)。 Ma 等(2018)指出,血红素氧化酶-1(HO-1)和磷酸酰胺腺嘌呤二核苷酸醌氧化还原酶-1(NQO-1)是两种重要的II 相解毒酶, 可促进细胞的抗氧化能力,硫氧还蛋白还原酶-1(TXNRD1)是一种重要的奶牛抗氧化硒蛋白,XCT 是一种控制细胞内谷脯甘化合成来源的重要转运载体, 他们是参与机体抗氧化的关键基因, 这四种基因的表达受到Nrf2-ARE 的调控。 奶牛泌乳、代谢增强、氧化应激、炎症损伤往往相伴而生,但又相互影响,甚至会影响奶牛产奶和乳品质, 给奶业生产带来巨大损失。 氧化应激反应的发生既与代谢中氧气需求增加有关,又与机体本身抗氧化能力有关,因此,
本文假设如果减少氧气的供应, 或者激活机体抗氧化基因表达,就会对氧化应激、炎症反应有一定的缓解作用。 本试验旨在以体外培养的大鼠乳腺上皮细胞(HC11) 为载体, 探讨催乳素和氧气对HC11 细胞中参与抗氧化的关键基因表达的影响。
1 材料与方法
1.1 细胞模型 HC11 细胞来自于妊娠中期BALB/c 大鼠的细胞系,由美国佛蒙特大学提供。
1.2 试验设计 采用随机分组试验设计,将细胞分为2 组,每组4 个处理,每个处理3 个重复,每个重复4 个孔, 研究有氧和缺氧状态下催乳素对H2O2诱导HC11 细胞乳蛋白相关基因、 葡萄糖转运因子和细胞因子基因表达的影响。 试验设计方案见表1。

表1 试验设计
1.3 试验方法 复苏后的HC11 细胞以2 ×105个/孔等密度接种于6 孔板, 先用生长培养基CGM(DMEM/F12,10% FBS,100 U/mL 双抗,5 μg/mL 胰岛素,10 ng/mL EGF,所有试剂均来自美国Gibco公司)培养至细胞80%贴壁后,从培养箱取出细胞,用PBS 清洗细胞3 次,换用前激素诱导培养基PIM (10% charcoal-treated 马血清) 培养24 h后,再用PBS 清洗细胞3 次,再换用激素诱导培养基HIM(0.1 μmol 地塞米松,5 μg/mL 催乳素)分常氧组 (5% CO2培养箱) (0 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+
0.01 μmol H2O2和5 μg/mL PRL+0.2 μmol H2O2)和缺氧组(缺氧培养箱,O2<6%)(0 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+0 μmol H2O2,5 μg/mL PRL+0.01 μmol H2O2和5 μg/mL PRL+0.2 μmol H2O2)分别培养3 h 后收获细胞,用于测定各基因表达。
总RNA 的提取采用RNeasy Plus Kit (Qiagen)试剂盒并按说明书进行,RNA 的转录采用Super-ScriptTMIII Reverse Transcriptase (Invitrogen)试剂盒并按说明书进行,以GAPDH、β-actin 和HPRT为内参,采用SYBR green 试剂盒并按说明书对各基因mRNA 表达量进行分析,基因表达量用2-ΔΔCT法进行计算。 各个基因引物信息详见表2。

表2 基因引物信息
1.4 数据统计分析 所有原始数据用SAS 9.1 中TUKEY 单因素方差分析进行数据处理,结果用“平均值±标准误”表示。P<0.05 表示差异显著。
2 试验结果
2.1 有氧和缺氧状态下催乳素对HC11 细胞蛋白质相关基因表达的影响 由表3 可见, 在有氧状态下, 与对照组相比, 催乳素均显著诱发了CSN2、CSN1S1 和α-LA 基因的表达(P< 0.05),且随着H2O2浓度的增加, 三种蛋白基因的表达量显著下降(P<0.05)。 在缺氧状态下,催乳素显著诱发了CSN2、CSN1S1 和α-LA 基因的表达 (P<0.05),随着H2O2浓度的增加,CSN2、CSN1S1 基因的表达量显著下降(P< 0.05)。但对α-LA 基因而言,随着H2O2浓度的增加,其表达量反而显著上升(P< 0.05)。 与有氧状态相比,在缺氧状态下,不管有无催乳素作用, 也不管H2O2浓度高低,CSN2和CSN1S1 基因表达量均显著降低(P< 0.05),而α-LA 基因的表达量则显著上升(P<0.05)。

表3 有氧和缺氧状态下PRL 对HC11 细胞乳蛋白相关基因表达的影响
2.2 有氧和缺氧状态下催乳素对HC11 细胞抗氧化基因表达的影响 由表4 可见, 与对照组相比,在不使用H2O2的条件下,不管有氧还是缺氧,催乳素对抗氧化基因的表达影响差异不显著(P>0.05)。 但在有氧且使用H2O2的条件下,与对照组相比,HO-1、XCT、TXNRD1 基因的表达量显著增加, 且随着H2O2剂量的增加而显著增加(P<0.05);对NQO-1 基因,其表达量却显著降低(P<0.05), 随着H2O2剂量的增加其表达量显著减少(P< 0.05)。 在缺氧并使用H2O2的条件下,XCT、TXNRD1、NQO-1 和HO-1 基因的表达量与有氧状态下的变化趋势一致(P< 0.05),即在有催乳素作用的基础上,与有氧状态相比,缺氧显著诱发了抗氧化基因XCT、TXNRD1 和NQO-1 的表达,并呈显著的剂量效应(P< 0.05),但对TXNRD1 基因,其表达量的变化却正好相反(P< 0.05)。
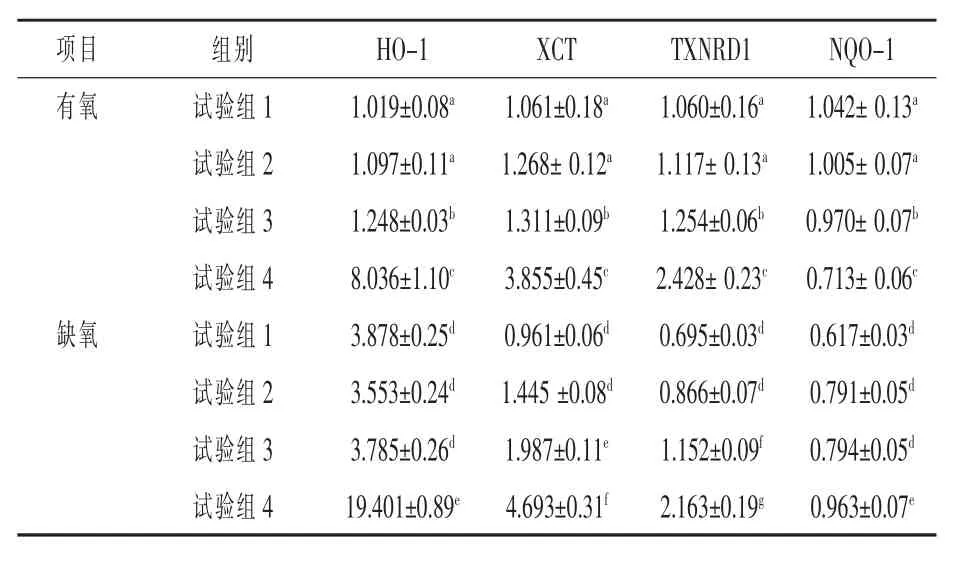
表4 有氧和缺氧状态下PRL 对HC11 细胞氧化应激信号通路相关基因表达的影响
2.3 有氧和缺氧状态下催乳素对HC11 细胞GLUTs 和炎症因子基因表达的影响 由表5 可见,在有氧状态下,催乳素对GLUTs 基因的表达影响差异不显著(P> 0.05),却显著降低了炎症因子IL-1β 的基因表达量(P< 0.05);当有氧并使用H2O2的条件下,GLUT8 基因的表达量显著降低, 且有明显的H2O2剂量效应 (P< 0.05),但GLUT1 基因表达变化不明显(P> 0.05)。 对于炎症因子,催乳素显著降低了IL-1基因表达量,且其表达量随着H2O2剂量的增加而增加 (P<0.05)。 在缺氧状态下,催乳素显著增加了葡萄糖转运因子GLUT1 和GLUT8 基因的表达 (P<0.05), 也显著增加了IL-1 基因的表达 (P<0.05)。 但在有氧并使用H2O2的条件下,低剂量的H2O2显著增加了GLUT1 基因的表达量, 高剂量却显著降低了其表达量 (P< 0.05);H2O2均显著增加了GLUT8 基因的表达量,且随着剂量的增加而增加(P< 0.05)。 对于炎症因子,H2O2显著增加了IL-1基因表达量,且均随着H2O2剂量的增加而增加(P< 0.05)。

表5 有氧状态下PRL 对HC11 细胞GLUTs和炎症因子基因表达的影响
3 分析与讨论
3.1 有氧和缺氧状态下催乳素对HC11 细胞蛋白质相关基因表达的影响 本试验在有氧和缺氧状态下采用不同浓度的H2O2对HC11 细胞诱导培养3 h,结果发现在有氧状态下,与未经H2O2诱导组相比, 催乳素显著诱发了CSN2、CSN1S1 和α-LA 基因的表达。当HC11 细胞被诱导处于氧化应激状态下时,其CSN2、CSN1S1 和α-LA 基因的表达依然显著高于未添加PRL 组,这说明催乳素在泌乳和乳蛋白合成方面的作用明显。 但常氧状态下随着H2O2浓度的增加,三种蛋白基因的表达量也显著下降, 说明氧化应激的确是奶牛养殖业和奶业生产中的一个重要问题, 虽然催乳素的存在对泌乳和乳蛋白的合成有一定的保护作用,但也不能完全抵挡氧化应激本身造成的损伤。 研究表明,PRL 是雌性动物乳腺生长发育、启动和维持泌乳必不可少的激素。在乳腺中,PRL 发挥作用主要通过JAK/Stat 和PI3K-mTORC2 信号通路调控乳蛋白的合成(丁洛阳等,2016)。金亚亚等(2021)研究发现,100 ~ 400 μg/L 催乳素可以促进BMECs 增殖。200 μg/L 催乳素对BMECs 酪蛋白合成相关基因表达具有较好的促进效果。 100 μg/L催乳素在翻译水平上对促进酪蛋白合成效果最好(Lee 等,2015)。 这和本试验结果基本一致。 由此可见, 催乳素对乳腺蛋白质合成的积极作用是基本得到定论的。
泌乳期奶牛需氧量增大, 导致活性氧产量的增加,进而导致奶牛产生氧化应激的产生,而减少氧气的摄入是否会缓解氧化应激反应所产生的氧化损伤鲜见报道, 因此本试验同时开展了缺氧状态下的研究。 研究表明,在缺氧状态下,催乳素显著诱发了CSN2、CSN1S1 和α-LA 基因的表达,但随着H2O2浓度的增加,CSN2、CSN1S1 基因的表达量也显著下降,这与有氧状态下的趋势一致。与有氧组相比,不管是在对照组还是催乳素诱导组,也不管是在低浓度还是高浓度的H2O2诱导组,其CSN2 和CSN1S1 基因表达量均显著降低。 这说明, 缺氧状态下催乳素虽然可以缓解氧化应激带来的损伤,但是缓解的能力非常有限,氧气不足更加剧了氧化应激造成的损伤, 对乳腺泌乳和乳蛋白合成的影响更大。 这一点可以由本试验中得到的在缺氧和氧化损伤状态下α-LA 基因的表达量的显著上升来解释,氧化损伤也严重,机体为了对抗损伤, 就需要动员诸如α-LA 等具有免疫功能的基因来参与损伤的修复, 进而尽可能的保持乳腺健康和正常泌乳功能。
3.2 有氧和缺氧状态下催乳素对HC11 细胞氧化应激信号通路相关基因表达的影响 郭咏梅(2020)研究发现,围产期和泌乳前期的高产奶牛代谢水平明显增强,机体抗氧化功能显著降低,很容易诱发氧化应激反应而导致疾病易感性增加,如乳腺炎、酸中毒、酮症等。因此,除了泌乳激素调控, 如何动员机体的抗氧化基因来对抗氧化应激而维持机体的代谢平衡也很关键。
抗氧化系统主要包括酶类抗氧化剂和非酶类抗氧化剂, 两者协同保护细胞免受自由基诱导的氧化损伤。 Lee 等(2015)研究表明,TrxR1 是细胞质中一种重要的硒蛋白,在氧化应激条件下,Nrf2快速地易位入核并与EpRE、ARE 结合,EpRE、ARE 存在于其目标抗氧化酶 (HO-1,NQO-1、GCLC 和TrxR1 等) 基因的启动子区域并诱发其转录。本试验研究表明,不管在有氧还是缺氧状态下,催乳素对抗氧化基因的表达影响不大。但在氧化应激状态下, 催乳素显著增加了HO-1、XCT、TXNRD1 基因的表达量,且随着H2O2剂量的增加而显著增加。 说明当机体受到氧化应激损伤时,HO-1、XCT、TXNRD1 基因表达就会被激活,从而提高机体抗氧化应激的能力, 这与李前辉等(2021)的研究结果一致。
Abd 等(2021)试验还发现,NQO-1 基因的表达量显著降低,且随着H2O2剂量的增加而显著减少。 研究表明,NQO-1 参与抗氧化活性和应激下关键调控蛋白的稳定, 对于本试验中NQO-1 基因的表达量显著降低,且随着H2O2剂量的增加而显著减少的结果与前人的研究结果不太一致,其原因可能是氧化应激状态造成了NQO-1 基因的损伤,也有可能与NQO-1 基因的多态性有关,同样的矛盾结果也有报道。 据报道,NQO1 基因缺失,易使小鼠受到氧化应激的影响,发展为肿瘤;但也有报道称NQO1 敲除可减少胶质母细胞瘤细胞的增殖, 而过度表达则增加细胞增殖(Dinkova等,2010)。除了酶活性、抗氧化活性和应激下关键调控蛋白的稳定作用之外,已有证据表明NQO-1在癌症生物学中具有双重效应, 既可以作为肿瘤抑制因子,也可以作为肿瘤促进因子,因此,产生此结果的具体原因还有待进一步研究。
本试验研究结果还发现, 在缺氧并处于氧化应激状态下,HC11 细胞中HO-1、XCT、TXNRD1和NQO-1 基因的表达量均显著上调, 且随着剂量的增加而显著增加。在有催乳素作用的基础上,与有氧状态相比, 缺氧显著诱发了抗氧化基因XCT、TXNRD1 和NQO-1 的表达, 并呈显著的剂量效应,说明缺氧更加诱发了氧化应激损伤,为了对抗这种损伤,机体会动员抗氧化基因的表达,进而增强机体的抗氧化能力。
3.3 有氧或缺氧状态下催乳素对HC11 细胞GLUTs 和炎症因子基因表达的影响 本试验研究结果表明, 在常氧状态下, 催乳素显著增加了GLUTs 基因的表达量,却显著降低了炎症因子的基 因 表 达 量 (P< 0.05)。 H2O2均 显 著 降 低 了GLUT8 基因的表达量(P< 0.05)。 葡萄糖转运因子(GLUTs)是一类调控细胞外葡萄糖进入细胞内的跨膜蛋白家族,参与糖代谢、炎性反应和免疫应答等过程。 张青玲 (2020) 研究表明,GLUT1 和GLUT8 转运蛋白在乳腺发育过程中表达增加,泌乳时GLUT1 mRNA 表达增加20 倍,GLUT8 蛋白表达增加2 倍。说明在未发生氧化应激的乳腺中,PRL 有利于增加葡萄糖的转运能力而为乳腺提供能量,此时炎症因子处于低表达状态;而在机体产生氧化应激时,炎症基因表达逐渐增强,说明氧化应激状态下炎症反应同步开始。本试验中,常氧状态下, 在存在PRL 的前提下,GLUT1 表达差异不显著,GLUT8 在氧化应激状态下表达量显著降低;催乳素显著降低了IL-1基因表达量,但随着H2O2剂量的增加而增加,这也正好说明在氧化应激状态下,葡萄糖的转运受阻,炎症反应加强,机体的主要机能由泌乳和乳成分合成向抗炎转变。这一推测在缺氧状态下表现的更为明显。 本试验结果中,在缺氧状态下,催乳素显著诱导了葡萄糖转运因子GLUT1 和GLUT8 基因的表达, 低剂量的H2O2显著增加了GLUTs 基因的表达量, 高剂量GLUTs 表达量与低剂量相比开始显著降低;H2O2均显著增加了GLUT8 基因的表达量, 且随着剂量的增加而增加。 H2O2显著增加了IL-1β 基因表达量,且均随着H2O2剂量的增加而增加(P<0.05)。 这说明缺氧加剧了机体对抗炎症时对能量的消耗,但随着氧化应激的持续加剧,机体功能开始紊乱,炎症反应持续增强,组织开始损伤。 本试验中GLUT1 和GLUT8 的比例关系的改变和对于炎症因子基因表达的持续增加就说明了这一点。
IL-1β 又称淋巴细胞活化因子, 也是由活化的单核巨噬细胞产生, 在组织炎症状态下发挥着重要功能。 Dinkova 等(2010)研究结果表明,不同浓度LPS 刺激奶牛乳腺上皮细胞24 h 后,细胞中IL-1β、IL-6 和IL-8 的mRNA 相对表达水平随着LPS 刺激浓度的增加而增加,说明IL-1β、IL-6 在炎症反应中会被激活以起到抗炎作用, 这也进一步验证了本试验结果及其相关推测。
4 结论
催乳素有利于改善氧化应激对乳腺乳蛋白基因表达的影响, 其作用是通过增加抗氧化基因的表达、 激活炎症因子的表达和维持葡萄糖转运因子的正常运转而实现的。
