细菌对抗宿主细胞自噬策略
2022-06-17赵维薇
赵维薇 ,王 辰 ,肖 鹏,高 洪
(1.大理大学 公共卫生学院,云南 大理 671000;2.云南农业大学 食品科学技术学院,云南 昆明 650201;3.大理大学 工程学院,云南 大理 671000;4.云南农业大学 动物医学学院,云南 昆明 650201)
自噬(autophagy),是一种进化保守的细胞利用溶酶体降解自身胞质内物质及其受损细胞器的过程。当面临营养缺乏、氧化应激、缺氧和病原体入侵等细胞压力时,自噬可以迅速诱导细胞适应环境变化。自噬在包括细胞发育在内的众多细胞生理过程中发挥着重要作用,比如面临饥饿时维持氨基酸库的平衡;同时与多种人类疾病有关,比如神经退行性变、癌症、病原体感染、炎症性疾病等[1-3]。当机体面临细菌、病毒等病原体入侵时,自噬会被激活,在胞质中的细菌、囊泡或吞噬体中的细菌被清除过程中发挥重要作用[4]。经过与宿主细胞自噬系统的长期互作,细菌进化出各种各样的机制来逃避、对抗自噬。现介绍细胞自噬系统靶向病原菌的机制,并重点探讨细菌应对细胞自噬的策略。
1 自噬概述
按照自噬识别的特异性,细胞自噬分为选择性自噬和非选择性自噬。一般认为,营养缺失导致的自噬为非选择性自噬,其作用主要是为细胞提供能量和必需品。选择性自噬参与特异性物质的降解,包括线粒体自噬(mitophagy)、过氧化物酶体自噬(pexophagy)、异体自噬(xenophagy)、聚集体自噬(aggrephagy)、脂噬(lipophagy)等[5]。选择性自噬需要受体(cargo receptor)与自噬体之间的识别,而非选择性自噬没有[6]。根据吞噬物进入溶酶体的途径,可将细胞自噬分为巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导的自噬(chaperone-mediated autophagy)三类[7]。巨自噬是自噬的一种经典形式,也是目前研究得最深入的一种形式,通常将巨自噬简称为自噬。经典的自噬过程包括几个步骤:启动(initiation)、成核(nucleation)、形成分隔膜及分隔膜延伸(formation and expansion of phagophore)、自噬体与溶酶体融合(fusion of autophagosome with lysosome)、自噬溶酶体内物质降解(degradation in autolysosome)[8-10]。它受到一系列信号通路的严格调控,涉及一系列事件,包括自噬相关(autophagy-related ,ATG)蛋白含量的改变,亚细胞定位的改变和翻译后修饰。在酵母细胞中,研究人员鉴定了36个ATG蛋白,可以在不同的环节参与自噬反应。15个ATG核心蛋白(ATG1-10,12-14,16,18)参与自噬体生物合成,它们在哺乳动物中高度保守[11]。自噬的启动受到细胞中两个重要蛋白的调控,能量传感器AMP活化激酶(AMP-activated kinase,AMPK)和哺乳动物雷帕霉素靶蛋白复合物1(mammalian target of rapamycin complex 1,mTORC1)。两个泛素化结合系统(ubiquitin-like conjugation systems,UBL system)控制ATG12-ATG5-ATG16L1复合物和微管相关蛋白轻链3(microtubule-associated protein light chain 3,LC3)蛋白-磷脂酰乙醇胺(phosphatidylethanolamine,PE)的产生,这对于自噬体的延伸和闭合非常重要[12]。不断延展的分隔膜最终自我融合形成了一个封闭的双层膜结构——自噬体,待降解的物质被包裹在自噬体中。最后在小GTP酶Rab7、C类空泡蛋白分拣蛋白(class C vacuolar protein sorting,VPS)、可溶性N-乙基马来酰亚胺敏感因子黏附蛋白受体(soluble N-ethylmaleimide-sensitive factor attachment protein receptor,SNARE)样蛋白以及溶酶体膜蛋白等因子的参与下,自噬体与内源溶酶体的囊泡融合,以实现对囊泡内物质的降解和循环利用[8]。
2 异体自噬
异体自噬作为细胞利用自噬清除入侵病原体的方式,需要经典自噬的核心组件,同时作为一种选择性自噬,还不能缺少自噬受体(autophagy receptor)的参与,也被称为自噬接头蛋白(adaptor)[13-14]。比如,结核分枝杆菌(Mycobacteriumtuberculosis)感染巨噬细胞时,被吞噬的结核分枝杆菌可以通过破坏吞噬体膜逃逸到细胞质中[15]。逃逸的细菌DNA被胞质传感器环磷酸鸟苷-腺苷合酶(cyclic guanosine monophosphate-adenosine monophosphate synthase,cGAS)识别,cGAS通过泛素连接酶Parkin和Smurf1激活泛素化,并招募自噬受体,如p62/SQSTM1,BRCA1基因1邻接蛋白(neighbor of BRCA1 gene 1,NBR1),核结构域10蛋白52(nuclear domain 10 protein 52,NDP52),视神经蛋白(optineurin,OPTN)诱导异体自噬[16-19]。
不仅胞浆中游离的细菌,含有细菌并被破坏的吞噬体也能被宿主吞噬体腔内的糖链所靶向[20]。最终,细菌被自噬体包裹并被送到溶酶体进行降解。在这一过程中,自噬可以产生抗菌肽并将其输送到自噬溶酶体腔室,从而促进细菌的杀灭[21-22]。
自噬受体的共同点是都包含2个重要的结构域,泛素结合域(ubiquitin-binding domain,UBD)和LC3结合域(LC3-interacting region,LIR)[23]。因此自噬受体可以通过UBD结合泛素化的病原体,再通过LIR基序将它们募集到自噬体膜上,这就是被普遍认识的异体自噬靶向病原体的泛素依赖途径。例如上皮细胞受感染时,鼠伤寒沙门菌(Salmonellatyphimurium)存在于含沙门菌的囊泡中(Salmonellacontaining vacuole,SCV)。然而,Ⅲ型分泌系统(typeⅢ secretion system,T3SS)的膜穿孔活性造成SCV膜的损坏,导致细菌渗出到胞质中。其中一些细菌被泛素包被,p62、NDP52和OPTN识别泛素化的鼠伤寒沙门菌,并将它们靶向至自噬体[24]。p62和NDP52也参与了福氏志贺菌(Shigellaflexneri)的异体自噬[25]。病原菌的泛素化是在E3泛素连接酶的作用下完成的,近年来研究的较为深入鉴定出了一些E3连接酶。比如LRSAM1(leucine-rich repeat and sterile α motif-containing-1)是沙门菌感染时进行泛素化的一种重要的酶[26]。在结核分枝杆菌诱导的异体自噬中,E3连接酶Parkin和Smurf1分别是K63和K48泛素化所必须的[27-28],其中Parkin是线粒体自噬中最主要的E3泛素连接酶。Smurf1还介导K48相关的单增李斯特菌(Listeriamonocytogenes)泛素化,限制李斯特菌在巨噬细胞中的增殖[27]。最近,有研究表明E3连接酶LUBAC(linear Ub chain assembly complex)在胞质鼠伤寒沙门菌表面结合M1型线性多聚泛素链(M1-linked linear poly-Ub patches),进而传递抗菌和促炎信号[29]。
除了泛素依赖途径,其他非泛素依赖途径也在异体自噬的细菌靶向中发挥着重要作用,包括核苷酸结合寡聚化结构域蛋白(nucleotide-binding oligomerization domain-containing,NOD)、半凝乳素(galectin)、甘油二酯(diacylglycerol,DAG)和补体C3蛋白(the complement protein C3),它们在福氏志贺菌和单增李斯特菌、沙门菌等的异体自噬中发挥作用[4]。另外SUDHAKAR等[30]通过对病原菌效应蛋白进行结构生物学和生物信息学分析,认为一些细菌效应蛋白可以直接与自噬受体(如p62和NDP52)甚至自噬核心组分相互作用。
3 细菌对抗细胞自噬的方式
有些细菌会被异体自噬靶向并清除,但也有一些细菌进化出了对抗或逃避这种宿主防御系统的机制。异体自噬发生时,宿主体内的自噬体可以包裹多种不同的胞质内细菌或载菌囊泡,但是不同种类的细菌进化出它们独特的机制以对抗宿主细胞自噬。CEMMA等[12]、WU等[4]以及KWON等[13]从不同的角度对细菌与宿主细胞的互作方式进行了综述和分类,本文进一步将细菌对抗宿主细胞自噬的策略分为三大类:逃避自噬、抑制自噬和操纵自噬。
3.1 逃避自噬(evasion of autophagy)福氏志贺菌进入宿主细胞后被囊泡包裹,之后可以逃避自噬体的捕捉进入胞质中。其细菌表面毒力蛋白IcsA(也被称为VirG)被ATG5识别后,激活异体自噬,这一过程需要宿主蛋白Tecpr1(tectonin domain-containing protein)将Atg5-靶向的细菌和WIPI2阳性吞噬体膜连接起来;与此同时,为了对抗宿主自噬防御系统,志贺菌分泌另一种细菌蛋白IcsB,与ATG5竞争结合IcsA,从而逃避自噬途径的识别[31-32]。近期研究表明IcsB是一种C18脂肪酰基转移酶,介导赖氨酸Nε-脂肪乙酰化。一种核内体ESCRT-Ⅲ复合物的组分CHMP5,是抗志贺菌自噬所必需的,IcsB介导的自噬抑制是CHMP5被修饰失活的一种间接结果[33]。
单增李斯特菌可以利用一种产生孔洞的毒素李斯特菌溶血素O(listeriolysin O,LLO)和两种磷脂酶C逃离吞噬囊泡。一旦进入了细胞质中,细菌表面的毒力因子肌动蛋白A(actin A,ActA)募集肌动蛋白相关蛋白Arp2/3复合体,催化肌动蛋白在细菌表面聚合,掩盖细菌以逃避自噬的识别[34]。李斯特菌还可以利用其表面的内化素 K(internalins K,InlK)将穹隆蛋白(major vault protein,MVP)招募到细菌表面,随后 ActA 取代 Inlk、肌动蛋白取代MVP来修饰细菌,阻止细菌泛素化以及自噬组件的识别[35]。
一些细菌不形成保护罩,而是通过靶向自噬受体而来逃避自噬。其中的一个例子是A 群链球菌(group AStreptococcus,GAS)。GAS的M1T1所克隆表达的半胱氨酸蛋白酶SpeB可以在宿主细胞中降解p62、NDP52和NBR1 ,以防止被宿主自噬所识别[36]。
3.2 抑制自噬(inhibition of autophagy)结合自噬发生的过程来看,细菌对自噬的抑制主要包括抑制自噬启动、抑制自噬体形成、抑制自噬体的成熟3种情况。
3.2.1抑制自噬启动 一些细菌通过抑制自噬的启动以逃脱宿主细胞自噬。一种小GTP酶Rab1A,是自噬启动中的必要因子。鼠伤寒沙门菌分泌的效应蛋白SseF和SseG通过直接与宿主的Rab1A互作抑制自噬的启动。这种互作影响了Rab1A 与鸟苷酸交换因子(guanine nucleotide exchange factor,GEF)、TRAPPIII复合体的相互作用,从而阻碍Rab1A的激活,导致自噬启动组件ULK1的募集和激活受到抑制,还降低了磷脂酰肌醇3-磷酸(phosphatidylinositol 3-phosphate,PI3P)的生物发生,最终阻碍自噬小体的形成[37]。与此类似,志贺菌T3SS效应蛋白VirA和胞外肠道致病性大肠杆菌(enteropathogenicEscherichiacoli,EPEC)效应蛋白EspG含有GTP酶激活蛋白区域,能特异性地使Rab1失活,导致自噬诱导的抑制[38]。
鼠伤寒沙门菌通过调控AMPK依赖的mTOR激活途径抑制自噬启动。沙门菌感染通过沙门毒力岛2(Salmonellapathogenicity island 2,SPI2)T3SS依赖的方式诱导溶酶体降解Sirt1、LKB1和AMPK。AMPK激活复合体被降解,导致mTOR的激活以及自噬的抑制[39]。囊泡型H+ATP酶(Vacuolar-type H+ATPase,V-ATPase)是发挥异体自噬作用所必需的,沙门菌T3SS效应蛋白SopF催化宿主载菌囊泡上V-ATPase的ADP核糖基化,解除载菌囊泡上mTORC1的功能,模仿氨基酸充盈的状态,从而抑制异体自噬的启动[40]。结核分枝杆菌H37Rv菌株通过其效应蛋白Eis干扰JNK-ROS信号通路抑制自噬的启动,增强其在细胞内的存活。Eis是一种N-乙酰转移酶,介导JNK特异性磷酸酶丝裂原活化蛋白激酶磷酸酶-7(mitogen-activatedprotein kinase phosphatase-7,MKP-7)的乙酰化和激活,导致JNK失活[41]。
一些胞外细菌毒素通过升高胞内环状AMP(cAMP)的水平抑制自噬的启动,cAMP是很多细胞内活动的一种二级信使,负向调控自噬。两种升高cAMP的毒素,炭疽杆菌的水肿毒素(edema toxin ,Edtx)和霍乱弧菌的霍乱毒素(cholera toxin ,Ctx),都可以通过蛋白酶K(PKA)途径抑制自噬包括抗菌自噬[42]。霍乱弧菌MARTX毒素具有PI3P特异性磷脂酶A1(PI3P specific phospholipase A1,PLA1)活性,可降低细胞内PI3P水平,抑制自噬和核内体运输[43]。
3.2.2抑制自噬体形成 核心自噬组件对于清除细胞内的细菌是必需的,一些细菌通过直接干扰核心自噬组件以抑制自噬体的形成。
嗜肺军团菌(Legionellapneumophila)利用T4SS效应蛋白RavZ抑制自噬。RavZ是一种半胱氨酸蛋白酶,可以不可逆的将LC3蛋白从PE上解离,抑制自噬体的形成[44]。RavZ通过自噬体的C端PI3P结合域和α3螺旋靶向自噬体膜,并通过其N端LC3结合域(LC3-interacting region,LIR)基序识别膜上的LC3分子[45-46]。近期的研究表明RavZ的蛋白水解活性干扰泛素向含沙门菌的囊泡募集[47]。
军团菌的另一个效应因子Lpg1137是一种可以降解突触融合蛋白17(syntaxin 17,Stx17)的丝氨酸蛋白酶,Stx17通过与ATG14L结合将PI3KC3复合物招募到线粒体相关的内质网膜,并参与自噬体-溶酶体融合[48]。 Lpg1137可导致Stx17的损耗以及PI3P形成的减少,说明军团菌可以干扰自噬体生物发生的早期[49]。李斯特菌产生两种磷脂酶PlcA和PlcB,下调PI3P水平,导致自噬前体结构产生的抑制,进而抑制异体自噬靶向胞质细菌[50]。
3.2.3抑制自噬体的成熟 自噬体包裹细菌后,还需要一个成熟的过程,包括自噬体与溶酶体融合,形成自噬溶酶体,从而利用溶酶体的酸性环境和丰富的酶类将细菌降解。
结核分枝杆菌的两种效应蛋白SapM磷酸酶和PknG激酶通过SecA2-依赖的蛋白转运系统进行转运,在抑制吞噬体和自噬体成熟、促进结核分枝杆菌在宿主细胞中生存和繁殖发挥作用。Rab5-Rab7交换对自噬体成熟很重要,但是SapM利用其磷酸酶活性抑制Rab5-Rab7交换[51]。另外,结核分枝杆菌H37Rv毒力因子PhoP和Esat-6通过抑制Rab7向含菌自噬体募集,阻断巨噬细胞和树突状细胞中自噬体的成熟[52-53]。
李斯特菌表达的毒力蛋白LLO可以阻止非经典自噬途径产生的LC3标记吞噬体的成熟[12]。金黄色葡萄球菌(Staphylococcusaureus)可以通过免疫显性表面抗原B(immunodominant surface antigen B ,IsaB)蛋白的表达抑制自噬体-溶酶体融合[54]。金黄色葡萄球菌侵入HeLa细胞后,激活丝裂原活化蛋白激酶14(mitogen-activated protein kinase 14,MAPK14)、磷酸化Atg5以及抑制自噬体与溶酶体的融合,从而逃避自噬的降解[55]。假结核耶尔森菌(Yersiniapseudotuberculosis)在巨噬细胞中也可以阻止含菌自噬体的成熟,避免菌体被自噬降解[56]。这些能阻断自噬溶酶体成熟的细菌逃避了自噬系统的降解作用后,还能采用多种策略在宿主细胞中生存并增殖。
3.3 操纵自噬(manipulation of autophagy)一些细菌在长期的进化过程中,进化出操纵自噬组件进行生存和繁殖的能力,它们将自噬组件如自噬体、自噬溶酶体变成其复制龛。
假结核耶尔森菌在巨噬细胞中改变自噬途径以阻止自噬体与溶酶体的融合,细菌被包裹在LC3标记的双层膜或者多层膜包裹的非酸性囊泡中进行复制[56]。吞噬体囊泡中的鼠疫耶尔森菌(Yersiniapestis)募集宿主蛋白Rab4a、Rab1b、Rab11b至吞噬体膜上,被LC3标记的自噬体包裹后,宿主蛋白阻止自噬体与溶酶体的融合,从而实现菌体在非酸性自噬体中增殖[56]。与此类似,金黄色葡萄球菌也可以在LC3标记的非酸性双层膜囊泡中存活和增殖;在李斯特菌感染的后期,细菌可以在LC3标记的单层膜含菌大吞噬泡中缓慢增殖[12]。在一些比较宽容的宿主细胞中(比如人和A/J鼠巨噬细胞),嗜肺军团菌可以延迟自噬的成熟至感染后4~6 h以分化成为耐酸形式,自噬体最终与溶酶体融合,而嗜肺军团菌最终也可以在酸性环境中复制,此过程涉及多种细菌效应蛋白[57]。
除了异体自噬,近期的研究发现线粒体自噬等选择性自噬也可能被细菌利用以促进自身的繁殖。比如李斯特菌通过LLO毒素的作用操纵宿主细胞发生线粒体自噬,从而降低线粒体产生的活性氧含量以促进自身繁殖[58]。还有一些细菌可以主动诱导宿主细胞的自噬,进而加以利用。幽门螺杆菌(Helicobacterpylori)感染时,利用空泡毒素(VacA)抑制mTORC1以诱导宿主自噬,并能在自噬体中增殖[59]。
细菌对宿主细胞自噬系统的操纵作用不一定都导致宿主细胞的损伤和死亡,也可以帮助细菌逃离宿主细胞。MIAO等[60]的研究发现尿路致病性大肠杆菌(uropathogenicE.coli,UPEC)感染膀胱上皮细胞后,会被自噬体靶向,随后穿梭到多泡小体中以形成自噬内涵体,最终停留在溶酶体中。但是它们没有在溶酶体中被降解,因为细菌可以上调溶酶体的pH值至中性,抑制溶酶体酸化。黏膜TRP通道3(mucolipin TRP channel 3,TRPML3)是溶酶体上一种瞬时受体电位阳离子通道,能触发Ca2+流出,启动溶酶体的外排作用,导致被外泌体包围的细菌排出。在此过程中自噬减轻了受感染细胞的细菌负载。
4 展望
细菌病原体和宿主自噬系统之间是相互作用的。一方面,细菌可以被宿主自噬系统限制,最终被自噬清除;另一方面,病原细菌进化了很多机制对抗宿主细胞自噬,逃避、抑制甚至操纵宿主自噬组件。要解答胞内细菌如何被异体自噬靶向,细菌如何操纵宿主细胞自噬仍然需要更加深入的研究。细胞自噬系统对细菌感染的响应,以及细菌对自噬系统的调控,都涉及到细菌表达的效应蛋白,在细菌对抗自噬的诸多研究报道中可以看到研究人员已经发现了多种细菌调控自噬的效应蛋白,这是清晰阐述细菌与宿主细胞自噬系统相互作用过程的一个关键点。构建基因缺失突变菌株是研究细菌蛋白在自噬中功能的重要手段,需要综合运用多种新的技术,比如邵峰课题组在沙门菌抑制宿主细胞自噬的研究中就运用了转座子遗传筛选技术和CRISPR 筛选技术[40,61]。另外,SUDHAKAR等[30]还利用结构生物学和生物信息学技术,通过识别预测含有选择性自噬受体SQSTM1/p62,CALCOCO2/NDP52和MAP1LC3/LC3识别基序的细菌蛋白,预测了人自噬蛋白和56种病原菌效应蛋白的潜在互作关系,并通过试验证实了沙门菌效应蛋白YhjJ和自噬组件的相互作用。继续研究异体自噬靶向细菌的机制,发现细菌调控自噬的效应蛋白,探明其与宿主自噬系统的互作机理具有重要的意义,其理论可以指导靶向抗菌药物的开发,为防控动物和人的细菌感染性疾病治疗提供新的策略。
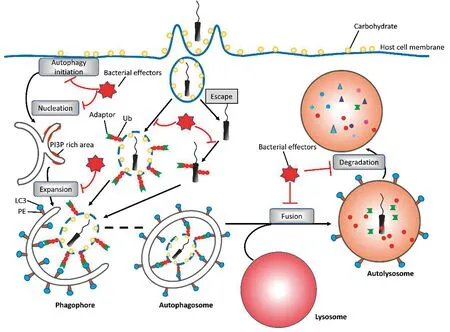
细菌对抗宿主细胞自噬示意图
