胡枝子幼苗对干旱胁迫的生理响应
2022-06-16曾继娟岑晓斐李双喜
曾继娟,朱 强,岑晓斐,李双喜
(1.宁夏林业研究院股份有限公司,宁夏银川 750004;2.种苗生物工程国家重点实验室,宁夏银川 750004;3.宁夏大学,宁夏银川 750004;4.宁夏西胡生态科技有限公司,宁夏泾源 756400)
干旱是全世界对植被影响最深、范围最大的自然因素之一[1-2]。随着全球气温的逐渐升高,极端高温天气反复出现,导致中纬度干旱半干旱地区的面积不断扩大,加剧了土地荒漠化[3-4]。土地荒漠化已成为国内外十分重视的环境问题之一,植物是构建防风固沙综合体系的主要因素[5],在我国北方干旱半干旱地区,发展沙生灌木是防治荒漠化的重要手段。干旱和半干旱地区的降水量较少、蒸发量大[6],水成为限制沙生植物生长发育的主要因素[7],缺水使植物面临干旱环境。研究表明,干旱胁迫可引起植物体内活性氧含量增加、抗氧化酶系统破坏、光合作用受阻、内源激素和渗透调节物质含量变化等一系列生理生化反应[8-10],轻度干旱可影响作物产量,严重干旱会对植物造成不可逆的伤害,甚至死亡[11-12]。
胡枝子(Lespedeza bicolor)是豆科胡枝子属多年生草本状半灌木,在我国东北、华北、西北等地广泛分布,具有耐旱、耐贫瘠等特性,是荒山造林的先锋树种;其营养价值高、生长快、萌蘖性好、利用年限长,也是优良的木本饲料[13-14]。目前,关于胡枝子的研究大多集中在生长特性[15-16]、栽培管理[17]、器官解剖结构[18-19]、营养价值[20]、光合作用[21]等方面,从生理角度探究其抗旱性的研究较少。因此,本试验研究了胡枝子幼苗对干旱胁迫的响应机制,旨在探明其生长过程中的需水关键期,为阐明胡枝子的抗旱能力及适宜的栽培技术应用提供依据。
1 材料和方法
1.1 试验地概况
试验地为宁夏银川市植物园(38°25′N,106°10′E),日照充足,干旱少雨,海拔1 115 m,年平均气温9.5~11.4 ℃,年平均降水量180 mm 左右,年平均蒸发量1 500~2 200 mm。
1.2 试验材料
试验材料为银川市植物园沙生植物国家林木种质资源库栽培的2年生健壮胡枝子苗。营养土、沙土以1∶1 的比例作为栽培土,经称重加入口径30 cm、高45 cm 的塑料桶,10 kg/桶。选取大小、生长表现基本一致的胡枝子苗,每桶栽植1 株,浇足水置于大棚内缓苗,避免自然降水进入桶内。缓苗期间进行正常的管理,以确保成活。
1.3 试验设计
于2020年4月11日开始试验,缓苗20 d。于4月30日测定桶内土壤田间持水量为28.16%,开始水分控制处理。采用随机区组试验,设置4 个水分梯度模拟干旱胁迫:对照(CK,田间持水量75%~80%),轻度干旱(T1,田间持水量60%~65%),中度干旱(T2,田间持水量45%~50%),重度干旱(T3,田间持水量30%~35%),5 次重复。采用便携式土壤水分速测仪(TDR-3,USA)及称量桶重两种方法保证每一梯度含水量保持在设定范围内,对因水分蒸发而降低水分含量的植株利用精度为0.05 L的量杯于17:00 及时加水补充。同时,需在桶面裸土上覆盖塑料薄膜防止水分流失。经干旱胁迫处理30 d,调查形态指标并测定光合特性,采集各处理植株叶片,用冰盒带回实验室,在-80 ℃条件保存备用,进行生理指标的测定。
1.4 指标测定与方法
新梢长、新梢粗分别用钢卷尺、游标卡尺进行测定。叶绿素相对含量采用SPAD-502 叶绿素仪测定。参照高俊凤[22]《植物生理实验指导》中的方法测定超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性;参照李合生[23]的实验方法测定丙二醛、游离脯氨酸(Pro)以及可溶性蛋白(SP)含量。
利用GFS—3000 便携式光合仪在晴朗无风的天气下测定光合作用。于8:30—11:00,从不同方位分别选取同一植株的叶片进行测定并标记(3 次重复),将叶片带回室内测量叶面积。测定时环境条件控制在空气CO2浓度为(375±5)滋mol/mol,光合作用的有效辐射率为(2 000±1)滋mol/(m2·s),空气温度(28.0±0.5)℃,使用自然光源,控制测定时间120 s。仪器自动记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)以及胞间CO2浓度(Ci)等参数。
1.5 数据处理与分析
利用Microsoft Excel 2010 软件处理数据并作图;利用SPSS 20.0 统计软件进行数据单因素方差分析,运用Duncan 法进行各处理间指标的差异显著性分析。
2 结果与分析
2.1 干旱胁迫对胡枝子幼苗生长特性的影响
胁迫30 d 后,胡枝子幼苗的新梢长、新梢粗及叶片数均随着干旱程度的加剧呈先升高后下降的趋势(表1),T2 处理最大,分别显著高于CK 76.80%、50.79%和13.68%(P<0.05);T3 处理最小,生长表现较弱,叶片出现萎蔫现象,说明重度干旱条件严重影响胡枝子幼苗的表型性状。
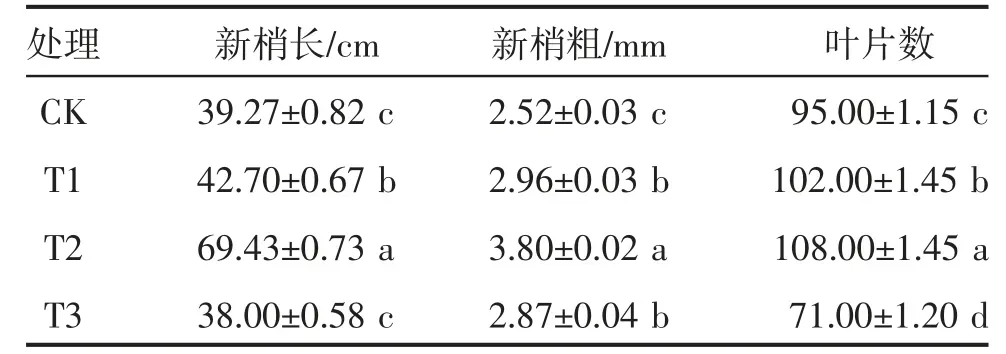
表1 干旱胁迫下胡枝子幼苗的生长特性
2.2 干旱胁迫对胡枝子幼苗光合特性的影响
2.2.1 干旱胁迫对胡枝子幼苗叶绿素含量的影响
胁迫30 d 后,叶绿素含量随着干旱胁迫的加剧呈先升高后降低的趋势(图1),各处理差异显著(P<0.05)。T1 处理最大,分别较CK、T2、T3 处理显著高5.0%、21.5%、63.3%(P<0.05),T3 处理叶绿素含量最低。
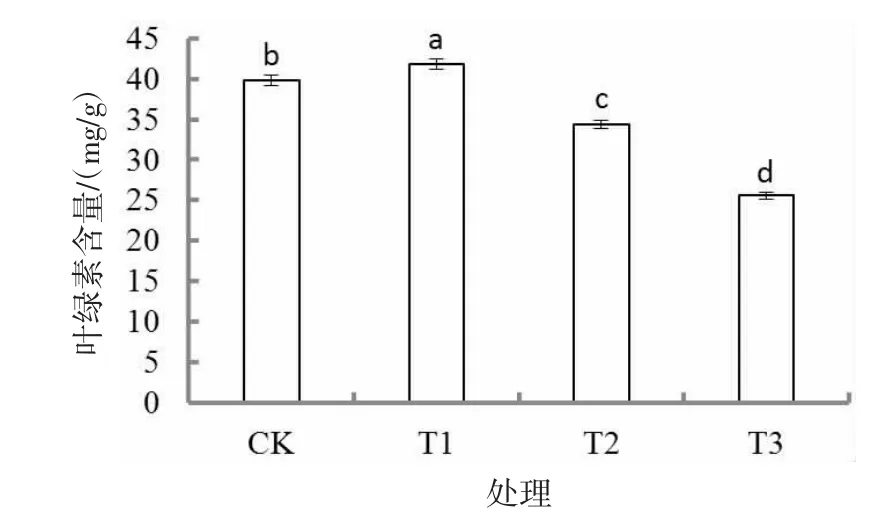
图1 干旱胁迫对胡枝子幼苗叶绿素含量的影响
2.2.2 干旱胁迫对胡枝子幼苗光合指标的影响
胁迫30 d 后,胡枝子幼苗Pn、Tr和Gs的变化规律一致(表2),均随着干旱胁迫的加剧直线下降,CK为最大值,T3 处理为最小值,各处理均存在显著差异(P<0.05)。其中,Pn、Gs在T2 处理下降幅度较大,分别较CK 显著下降了62.22%和59.71%(P<0.05);Tr在T1 处理时下降幅度最大,较CK 显著下降了37.09%(P<0.05)。Ci随干旱程度的加深变化趋势不明显,只是小幅度的升高再下降,T2 处理最大,较CK 高2.79%,但各处理差异不显著(P>0.05)。

表2 干旱胁迫下胡枝子幼苗的光合特性
2.3 干旱胁迫对胡枝子幼苗生理特性的影响
2.3.1 干旱胁迫对胡枝子幼苗抗氧化酶活性的影响
胁迫30 d 后,CAT、SOD 活性随着干旱胁迫的加剧先升高后降低(图2Ⅰ和图2Ⅱ)。CAT 活性T1 处理最高,分别较CK、T2 处理和T3 处理显著高190.5%、37.7%和265.8%(P<0.05),T3 处理为最低,且显著低于CK(P<0.05),各处理差异显著(P<0.05,图2Ⅰ)。T1 处理的SOD 活性最高,分别较CK、T2 处理和T3 处理显著高13.9%、3.9%和4.9%(P<0.05),T3 处理最低,但显著高于CK(P<0.05),T2 与T3 处理差异不显著(P>0.05,图2Ⅱ)。POD 活性随着干旱胁迫的加剧呈下降趋势,T2 处理急剧降低,CK 最大,分别较T1、T2 和T3 处理显著高22.9%、510.9%和777.4%(P<0.05),T2 和T3 处理差异不显著(P>0.05,图2Ⅲ)。
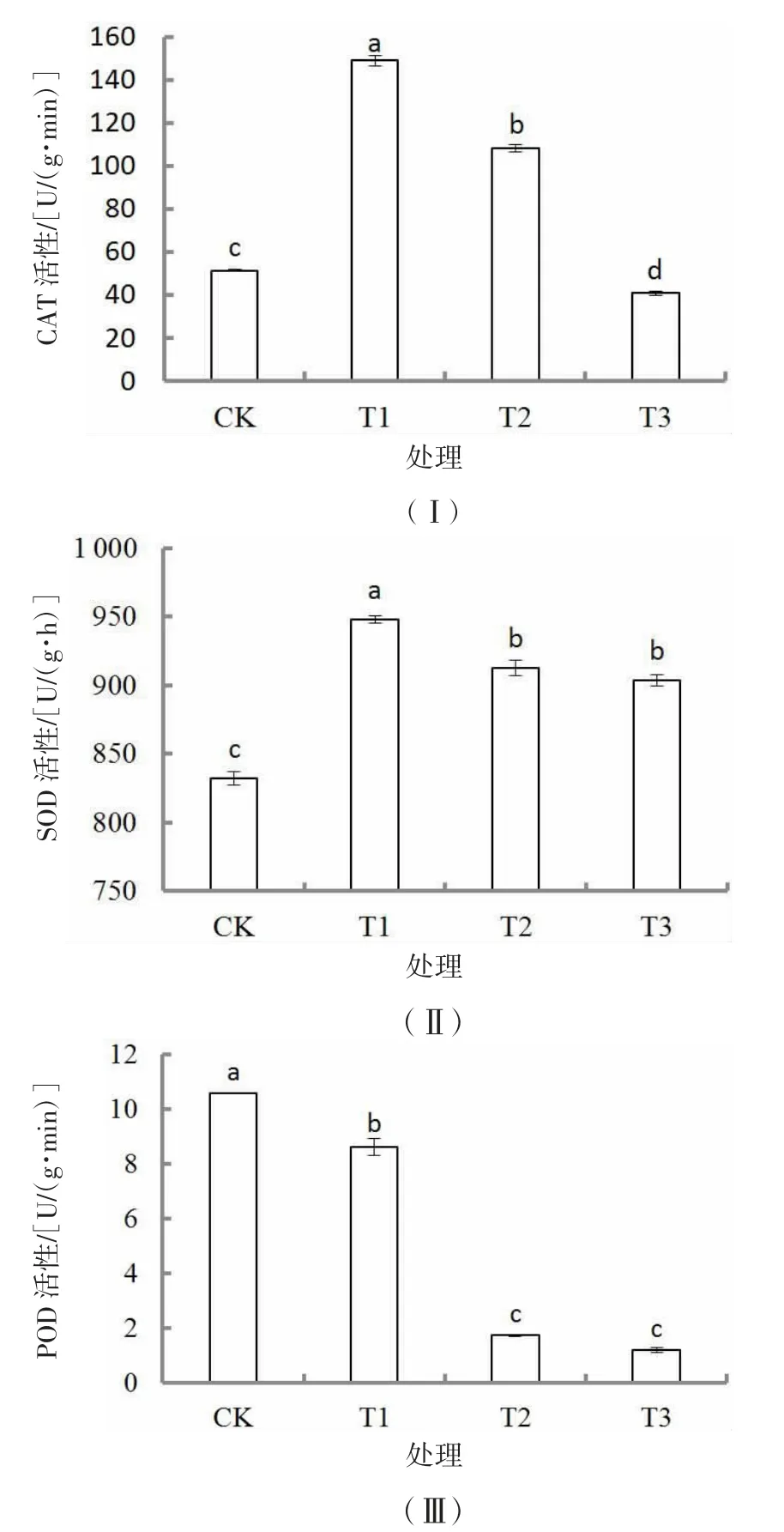
图2 干旱胁迫对胡枝子幼苗抗氧化酶活性的影响
2.3.2 干旱胁迫对胡枝子幼苗丙二醛含量的影响
胁迫30 d 后,丙二醛(MDA)含量随着干旱胁迫的加剧呈先升高后降低的趋势(图3),各处理均差异显著(P<0.05)。T1 处理MDA 含量最大,分别较CK、T2 处理和T3 处理显著高18.8%、72.1%和193.6%(P<0.05);T3 处理的MDA 含量最低,且T2与T3 处理较CK 显著下降(P<0.05)。
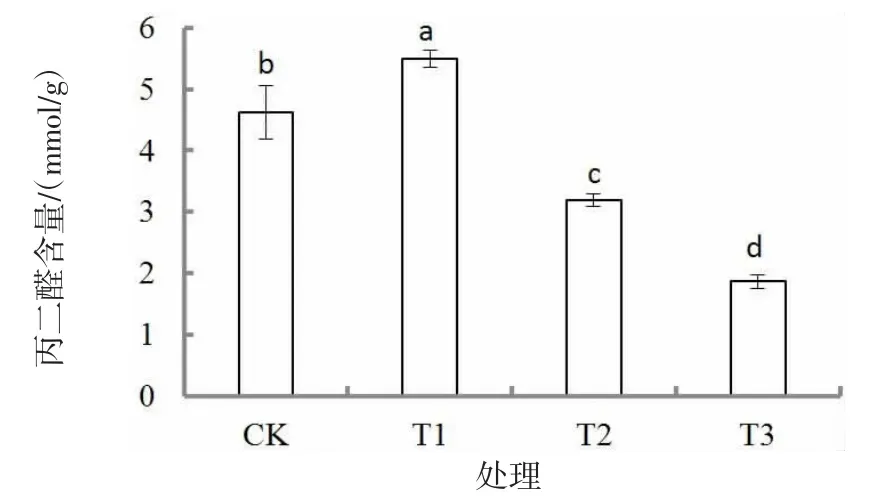
图3 干旱胁迫对胡枝子幼苗丙二醛含量的影响
2.3.3 干旱胁迫对胡枝子幼苗渗透调节物质含量的影响
胁迫30 d 后,脯氨酸和可溶性蛋白含量均随着干旱程度的加剧先升高后下降(图4)。T2 处理脯氨酸含量最大,分别较CK、T1 处理、T3 处理显著高65.3%、49.3%和46.5%(P<0.05),随着胁迫的进一步加剧,其含量快速下降;T3 处理脯氨酸含量为最低值,但显著高于CK(P<0.05)。T1 与T3 处理差异不显著(P>0.05,图4Ⅰ)。可溶性蛋白含量在T1 处理下急剧增加,达到峰值,分别较CK、T2 处理、T3 处理显著高155.5%、258.4%、383.1%(P<0.05);T3 处理最低,且显著低于CK(P<0.05);CK 与T2 处理、T2 与T3 处理差异不显著(P>0.05,图4Ⅱ)。
2.4 干旱胁迫下胡枝子幼苗各生理指标的相关性分析
由表3 可知,干旱胁迫下,胡枝子幼苗各指标的相关性分析结果表明,MDA 与SP、SOD、POD、CAT以及光合指标呈正相关(表3),与叶绿素含量呈显著正相关(P<0.05),与Pro 和生长指标新梢长、新梢粗呈负相关;SP 与除Pro 和生长指标新梢长、新梢粗外的其他指标呈正相关;SOD 与POD、叶绿素以及光合指标呈负相关;POD 与Pn、Gs,Pro 与生长指标新梢长、新梢粗呈显著正相关(P<0.05);CAT 与所有指标呈正相关;Pn与Tr、Gs呈极显著正相关(P<0.01)。
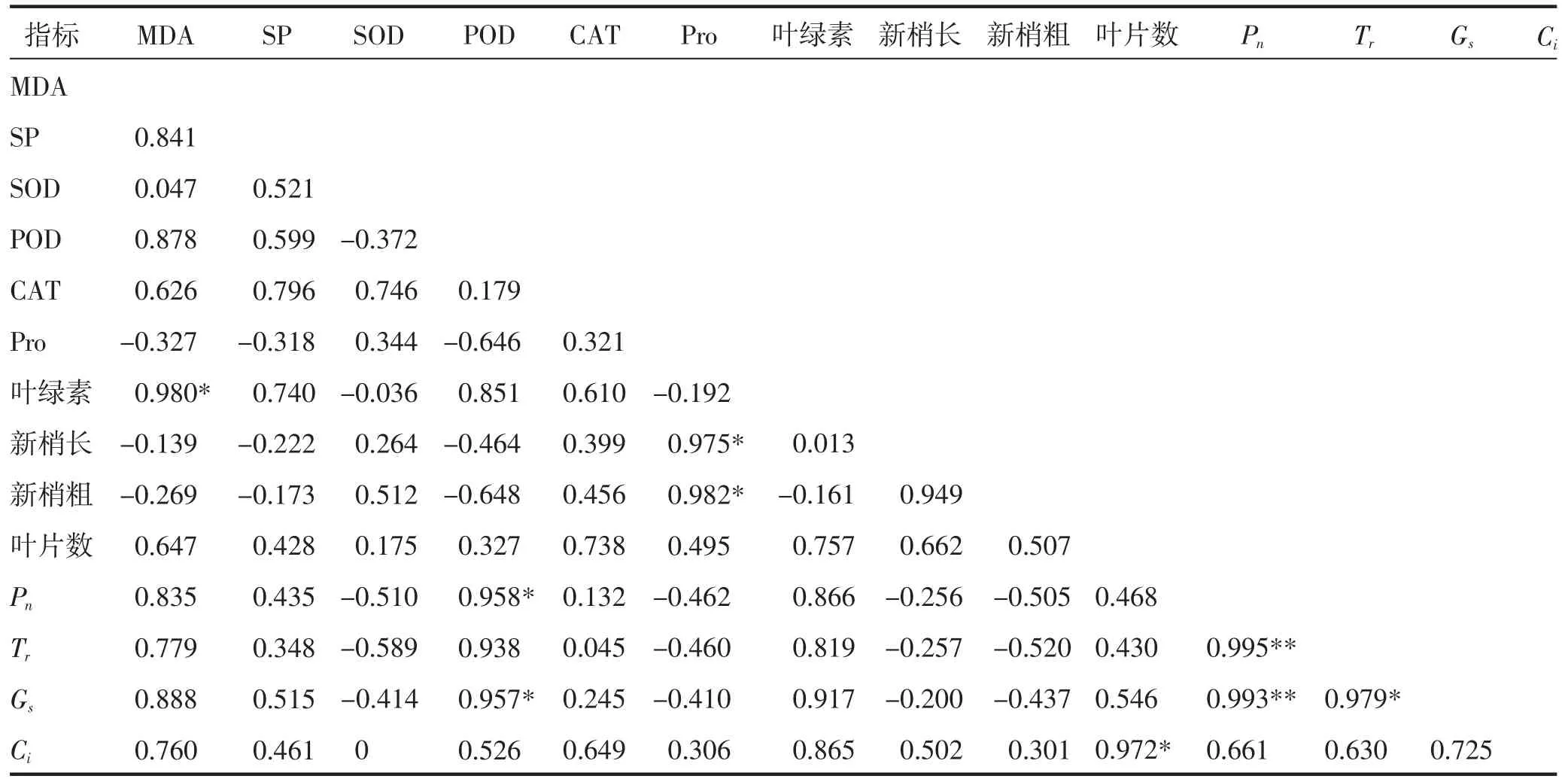
表3 干旱胁迫下胡枝子幼苗各指标间的相关性分析
3 讨论与结论
3.1 干旱胁迫对胡枝子幼苗生长变化的影响
水分是影响植物生长的重要因子,株高、新梢长和生物量等都是衡量植株生长质量的关键要素。本试验结果表明,胡枝子幼苗的新梢长、新梢粗及叶片数均随干旱胁迫的加剧表现出先升高后下降的趋势,在中度干旱下有峰值,重度干旱下有最小值,胡枝子幼苗逐渐出现萎蔫,单从生长变化来看,胡枝子幼苗可能不适合在重度干旱环境,即土壤含水量为30%~35% 田间持水量下生长。
3.2 干旱胁迫对胡枝子幼苗光合特性的影响
叶绿素是植物器官的基本组成部分,是植物光合作用、营养状况和生长趋势的重要指标[24]。本试验中,随着干旱胁迫的加剧,胡枝子幼苗的叶绿素含量先升高后降低,在T1 处理中达到峰值,在T3 处理中降幅最大。胡枝子幼苗的叶绿素含量在轻度干旱胁迫条件下显著高于水分充足和重度干旱胁迫条件下(P<0.05),这与郭相平等[25]对水稻研究中轻度干旱胁迫条件下叶绿素含量高于对照,而重度干旱胁迫叶绿素含量低于对照的试验结果不完全一致。其原因可能是干旱胁迫下胡枝子幼苗的叶片含水量降低,抑制其伸展,叶片增厚所致,同时,它也表明胡枝子幼苗的叶绿素在轻度干旱条件下没有受损或受损程度较轻[26]。试验随着干旱胁迫加剧,胡枝子幼苗叶绿素含量显著降低(P<0.05),这可能是在中度和重度干旱胁迫下植物叶绿体膜被破坏所致。
水分状况是影响植物光合作用的重要因素,干旱胁迫导致木本植物叶片的Pn和Tr降低[27]。自然条件下,光合有效辐射、气孔导度、蒸腾速率和胞间CO2浓度等因素在一定程度上决定了植物的光合特征[28]。净光合速率的变化规律在一定范围内与气孔导度和蒸腾速率变化一致[29]。本研究中,胡枝子幼苗的净光合速率、蒸腾速率和气孔导度的变化规律一致,均随着干旱胁迫的加剧直线下降。在中度干旱之前,Ci变化趋势与Pn相反,重度干旱时,Ci变化趋势与Pn相同。通常光合速率的降低有气孔限制和非气孔限制这两个原因。已有研究表明,如果Pn和Ci变化方向相同,Gs降低时Pn的降低是气孔因素造成的;如果Pn和Ci变化方向相反,Gs增大时Pn的下降主要是由非气孔限制,即叶肉细胞同化能力的降低引起[30]。因此,胡枝子幼苗光合速率的降低主要受气孔因素影响,同时,Ci随干旱程度加深的变化趋势不显著(P>0.05),只是小幅度先升高后降低,T2 处理最大,这是胡枝子幼苗适应干旱环境的主要调节机制,通过降低气孔导度减缓植物体内的水分散失。由此说明,胡枝子幼苗能够忍耐的干旱条件是中度干旱,即土壤含水量为45%~50%田间持水量,当干旱胁迫进一步加重时不利于该植物正常生长。
3.3 干旱胁迫对胡枝子幼苗生理特性的影响
3.3.1 干旱胁迫对胡枝子幼苗抗氧化酶活性的影响
在逆境中,植物会通过调节自身的保护酶防疫系统清除体内的活性氧,从而保护细胞免受损伤。研究表明,SOD、POD 和CAT 的酶活性与植物的抗旱性呈正相关[31-32],一般来说,在短期或轻度胁迫下,保护酶活性呈上升趋势,而在长期或中度胁迫下,保护酶活性先上升后下降[33]。本试验结果表明,随着干旱胁迫的加剧,CAT 和SOD 活性先升高后下降,峰值在轻度干旱,POD 活性呈下降趋势,尤其是中度干旱时急剧下降,说明一定强度的干旱胁迫提高了胡枝子幼苗叶片中抗氧化酶的活性,而重度干旱胁迫抑制了抗氧化酶的活性,这与罗婷等[34]对乳油木的研究一致。从试验结果可以看出,SOD、POD 活性CK、T1、T2 处理差异显著(P<0.05),而T2 与T3 处理差异不显著(P>0.05)。同时,CAT 酶活性随着土壤含水量的降低出现了不同幅度的波动,尤其在T1 处理时急剧升高,说明随着干旱胁迫,SOD 含量的增加使活性氧清除能力上升,CAT 含量增加。当干旱胁迫进一步加剧,即土壤含水量低于60%~65%田间持水量时,随着SOD、POD 活性的下降,部分来不及清除的自由基堆积后与CAT 反应形成复合物,可能导致其含量降低。可见,SOD、POD 和CAT 对干旱胁迫的响应有一定差异,但又相互协同作用,适当的干旱胁迫可使植物细胞产生更多的超氧阴离子自由基,并显著提高植株防御活性氧伤害的能力。
3.3.2 干旱胁迫对胡枝子幼苗膜脂过氧化作用的影响
植物所受的损害大多和活性氧引发的膜脂过氧化有较大联系,MDA 是膜脂过氧化终产物,它的产生会加剧膜的损伤,可间接反映膜系统所受伤害程度及其植物自身的抗逆性[35]。本试验中,T1 处理的MDA 含量显著高于CK(P<0.05),说明轻度干旱胁迫后植株的膜脂过氧化作用增强,加剧了膜损伤。T2 和T3 处理的MDA 含量显著低于CK(P<0.05),分别降低了30.95%、59.52%,说明中度和重度干旱胁迫均可减缓膜脂过氧化反应,但中度胁迫对其植株体内的MDA 含量影响程度低于重度胁迫。综合考虑,中度胁迫既可以满足植物生长需求,又是影响膜脂过氧化反应的转折点,能够减轻细胞损伤程度,保证植株细胞膜系统的稳定性和完整性,进而提高胡枝子幼苗的抗逆性,这与不同种衣剂配方对达乌里胡枝子抗旱中的MDA 含量变化结果相近[36]。
3.3.3 干旱胁迫对胡枝子幼苗渗透调节物质的影响
在逆境中,植物主要通过增加渗透调节物质(脯氨酸、可溶性蛋白等)的含量使细胞液浓度升高,降低渗透势抵御环境胁迫,维持细胞结构的稳定性,进而减轻逆境对自身的损害[36]。植物中的大多数可溶性蛋白是参与各种代谢的酶,蛋白质和脯氨酸会相互作用以增加蛋白质的亲水性[37]。本试验中,脯氨酸含量在T1 处理时上升幅度不大,与CK 相比差异不显著(P>0.05),但在T2 处理下急剧增加,达到最大,显著高于CK(P<0.05);可溶性蛋白含量随着干旱胁迫的加剧增长到一定程度则表现出下降的趋势,T1 处理最大,之后快速下降,当重度干旱胁迫时,下降速率逐渐放缓。由此推测,土壤含水量高于45%~50%田间持水量时,即轻度干旱时,胡枝子幼苗叶细胞膜系统未受到胁迫的伤害,细胞膜的稳定性强,有助于其生长;中度干旱时,脯氨酸含量急剧增加,可溶性蛋白含量迅速下降,可能是胡枝子幼苗适应干旱环境主动进行渗透调节的结果,脯氨酸、可溶性蛋白在调节细胞的渗透势、保护蛋白质分子和酶活性方面起着重要作用。干旱胁迫从中度(T2 处理)到重度(T3 处理)时,叶片的脯氨酸和可溶性蛋白含量均降低,可能是达到胡枝子幼苗对干旱条件的承受极限和需水关键时期,这与该试验中光合变化的趋势相符。
综上所述,适当的干旱胁迫有助于胡枝子幼苗生长,但是超过一定的干旱条件将起抑制作用。本试验通过分析胡枝子幼苗在干旱胁迫条件下的生理响应发现,土壤含水量为60%~65%田间持水量(T1 处理)时较利于胡枝子幼苗生长,土壤含水量为45%~50%田间持水量(T2 处理)是其生长的需水关键期。当然,植物对逆境的响应是受多因素、多途径调控的复杂过程,各因素相互配合、相互制约、协同作用,基于该试验仅是1年的研究结果,对于解释胡枝子在干旱胁迫下的生理生化机理尚有欠缺,需开展试验深入研究。
