利用小麦90K SNP芯片研究泰山22的遗传组成
2022-06-14王瑞霞翟晓灵李玉刚牟秋焕孙盈盈孙宪印米勇吕广德盖红梅钱兆国
王瑞霞, 翟晓灵, 李玉刚, 牟秋焕, 孙盈盈, 孙宪印,米勇, 吕广德, 盖红梅*, 钱兆国*
(1.泰安市农业科学院,山东 泰安 271000;2.青岛市农业科学研究院,山东 青岛 266100)
在水资源短缺、耕地面积锐减、耕作环境恶化、新的病原菌和害虫不断出现、极端气候频繁危害的背景下,为满足老百姓“吃的好、吃的安全、吃的健康、吃的放心”的新需求,小麦产量、品质、抗性等方面的遗传育种工作面临前所未有的挑战[1]。在小麦遗传改良中,突破性小麦新品种的选育与优异种质资源密切相关[2],而骨干亲本的创制和应用又是取得突破性进展的关键[3]。小麦骨干亲本如碧蚂4号、南大2419和小偃6号等的利用使我国小麦单产得到大幅提高[4],促成了我国主要麦区小麦品种数次更新换代[5]。因此,阐明小麦骨干亲本或主推品种在后代育成品种中的传递规律和遗传贡献,对更好地利用骨干亲本培育高产、优质、多抗小麦新品种具有重要意义[6]。鲁麦14具有稳产、广适、高抗等优点,曾成为20世纪90年代初期黄淮冬麦区推广面积最大的品种(累计推广783.5万hm2)[7-8],也是重要的育种亲本,8个省份的34家科研单位和育种企业利用鲁麦14为亲本,育成济麦20、青丰1号、烟农5158、淮麦20、皖麦46等39个品种;间接应用鲁麦14为亲本,7省1市的68家科研单位和育种企业共育成济麦22、烟农 999、烟1212、石麦26、淮麦22、中麦29等92个品种[9-12]。鲁麦18在3年区域试验中平均产量均位居前列,其冬前大蘖多、成穗率高;不孕小穗少,结实率高;千粒重稳定性好,容重高、品质好,是黄淮麦区重要的主推品种之一。
近年来,许多学者对小麦主推品种或骨干亲本的遗传构成进行了研究,李小军等[13]通过SSR(simple sequence repeat)分子标记对小麦品种百农AK58的遗传构成进行了分析,发现周麦11对百农AK58的遗传贡献更大,且多数特异位点与许多控制重要农艺性状基因相关;李俊等[14]用SSR标记和DArT(diversity arrays technology)标记解析了川麦104,认为其更多地继承了川麦42的遗传物质,且发现川麦104来源于双亲的遗传物质在不同染色体上分布不同;邹少奎等[15]通过SSR标记分析了大面积推广品种周麦22的分子遗传基础,发现亲本对周麦22的遗传贡献率在不同染色体间差异较大,其中亲本温麦6号对周麦22的遗传贡献较大;孔子明等[16]利用90K SNP标记对小麦周麦16及其亲本进行了全基因组扫描,发现周8425B对周麦16的遗传贡献率远远高于周麦9号;杨子博等[5]通过625个SSR标记分析了淮麦33及其双亲的遗传构成,发现其更多地继承了母本烟农19的遗传物质;李玉刚等[17]利用350个SSR标记和26 026个SNP标记分析了鲁麦14对青麦2号的遗传贡献,发现鲁麦14对青农2号的贡献远远大于烟农15。
众多学者利用SSR标记对小麦基因组开展了很多研究[5,13-20],但是SSR标记密度相对较低,无法满足当前高通量基因组研究需要。基于分子遗传学和基因组学获得的单核苷酸多态性SNP标记具有数量多、分布广、高度稳定、适于快速规模化筛查、易于基因分型等特点,为小麦育种提供了广阔的前景[21]。目前,小麦检测SNP常用的方法有重测序、小麦55K芯片、90K芯片、660K芯片等。本研究采用小麦90K芯片检测SNP标记,并分析了鲁麦14衍生品种泰山22号的遗传组成,结合农艺性状解析了亲本鲁麦18和鲁麦14对泰山22号的遗传贡献,为小麦分子标记辅助育种提供了依据。
1 材料与方法
1.1 试验材料
供试材料泰山22号及其亲本鲁麦18(母本)和鲁麦14(父本)均由山东省泰安市农业科学研究院小麦研究所提供。
泰山22号系谱为鲁麦18/鲁麦14,于2004年通过山东省和国家审定,审定编号分别为鲁种审字〔2004〕027和国审麦2004013。系谱追踪发现其亲本鲁麦18和鲁麦14遗传基础特别丰富,鲁麦18含有丰产3号、阿勃、泰山4号等丰产源,蚂蚱麦、碧玛6号等抗病源,咸农39、辉县红、矮丰3号等矮源。鲁麦14含有我国蚰包麦、美国包打300炮、智利欧柔等丰产源,日本农林10号、小罂粟等抗旱源和矮源,罗马尼亚的洛夫林13、意大利Virgilio、智利Rulofen和L277/4等著名的抗病源。供试材料叶片取自青岛市农业科学研究院城阳基地。
1.2 供试材料的90K芯片分析
供试材料的iSelect Illumina 90K芯片分析由北京博奥晶典生物技术有限公司完成,采用Genome studio v2011.1软件对返回的*.bmp文件和*.idat文件进行数据校正和SNP分型[17]。
1.3 亲本遗传贡献及泰山22的遗传组成图谱绘制
统计亲本间存在差异的位点,当泰山22号的分型与鲁麦18一致时计为鲁麦18的贡献位点,当与鲁麦14一致时计为鲁麦14的贡献位点,计算母本和父本遗传贡献率。采用Python语言(https://www.python.org)绘制泰山22号遗传组成图谱[17]。

1.4 农艺性状调查及统计分析
供试材料自2013—2016年连续3个小麦生长季,在山东省3个试点(青岛城阳、青岛平度和泰安肥城)及河南新乡进行种植,共计10个年点。每品种2行,行长2 m,行距25 cm。其中,2014—2015和2015—2016年度在青岛城阳进行了雨养和灌溉对比研究,用于比较供试材料农艺性状在2种水分供应条件下的差异。调查株高、抽穗期、开花期、旗叶长、旗叶宽、穗长、穗下节间长、穗叶距、抽穗度、小穗数、最大小穗粒数、穗粒数、千粒重、粒长、粒宽和粒厚共16个性状。利用SAS软件进行最优无偏估计(best linear unbiased prediction,BLUP),获得不同环境条件下亲本和子代的农艺性状最优估计值[17]。
2 结果与分析
2.1 多态性SNP位点在染色体上的分布
采用90K SNP芯片共获得20 139个具有染色体定位的SNP位点,这些位点不均匀地分布于小麦的21条染色体上(图1)。每条染色体分布72~141个SNP标记,这些标记在染色体间差异较大,1B、5B和2B染色体上分布最多,4D上分布最少;SNP标记在A、B、D基因组上分布也不均匀,以B基因组最多(10 253个,占50.91%),A基因组次之(8 049个,占39.97%),D基因组最少(1 837个,占9.12%)。其中,有7 513个SNP在泰山22及其双亲鲁麦18和鲁麦14上有差异,差异性标记的频率为37.31%。

图1 总SNP位点及差异SNP位点在染色体上的分布Fig.1 Distribution of total SNPs and differential SNPs on each chromosome
2.2 鲁麦18和鲁麦14对泰山22号的遗传贡献
在染色体水平,母本鲁麦18对泰山22号的遗传贡献范围为4.51%(6A)~99.02%(5B),5B和2B几乎整条染色体的遗传信息从鲁麦18传递至子代;父本鲁麦14对子代的遗传贡献率为0.98%~95.49%,遗传贡献率超过80.00%的染色体有3条,分别为3B、6A、7A,其中6A染色体绝大部分传递至泰山22。从全基因组水平,两亲本对泰山22号的遗传贡献大致相当(母本51.87%,父本48.13%),如在1D、2B等12条染色体均表现为鲁麦14的遗传贡献小于鲁麦18,在1A、1B等9条染色体均表现为鲁麦14贡献超过鲁麦18(图2、表1)。从3个亚基因组看,在A基因组上,鲁麦18的遗传贡献率为36.41%,鲁麦14的遗传贡献率为63.59%;在B基因组上,鲁麦18和鲁麦14的遗传贡献率分别为58.16%和41.84%;在D基因组上,鲁麦18的遗传贡献率为61.04%,鲁麦14的遗传贡献率为38.96%。

表1 鲁麦18和鲁麦14在21条染色体对泰山22号的遗传贡献Table 1 Genetic contribution of Lumai 18 and Lumai 14 on Taishan 22 with SNP on 21 chromosomes
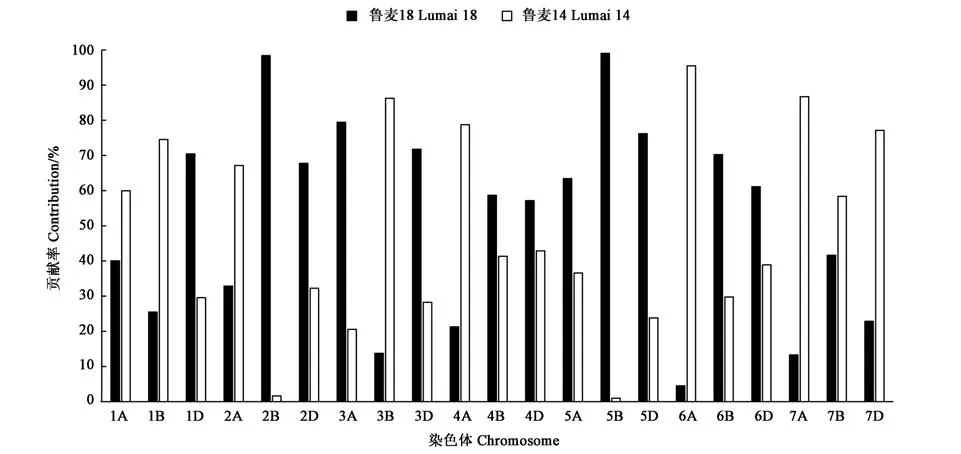
图2 SNP标记分析鲁麦18和鲁麦14对泰山22号的遗传贡献Fig.2 Genetic contribution of Lumai 18 and Lumai14 to Taishan 22 revealed by SNP markers
2.3 亲本遗传信息在泰山22号染色体上的分布
从图3可以看出,亲本遗传信息主要以染色体大片段形式传递到子代,如1B、2D、4A、5A、5B、6B、7A等染色体。有的染色体大片段是由位于同一遗传距离的数十甚至数百个SNP位点构成,如2D、5A、5B等的大片段;有的染色体大片段在整合遗传图谱中有较多重组,但泰山22号组合中表现为保守遗传,如2D、5B、6B等的大片段。

图3 泰山22号的遗传组成图谱Fig.3 Genetic composition map of Taishan 22
D组染色体由于标记数量较少,泰山22集成双亲鲁麦18和鲁麦14的区段较为集中,但是鲁麦18遗传给泰山22的区段较多。泰山22号的2B、2D、3A、5A、5B、5D、6B、6D染色体上遗传信息绝大部分来自鲁麦18,在1B、2A、3B、4A、5A、7A、7B染色体上则分布着较多的鲁麦14的遗传位点。
2.4 泰山22号及亲本的农艺性状表现
农艺性状BLUP分析(图4)表明,泰山22号的株高无论在自然降雨条件下还是灌溉和全部环境条件下,均高于高值亲本鲁麦18。泰山22号的抽穗期介于两亲本之间,并与两亲本差别不大。泰山22的开花期较两亲本提前。泰山22号的旗叶长和宽在自然降雨条件下介于两亲本之间,在灌溉和全部环境条件下,泰山22旗叶变长、变窄,更倾向于低值亲本鲁麦14。穗长在自然降雨条件下介于亲本之间,在灌溉和全部环境条件下变短,倾向于低值亲本。在3种不同的水分条件下,穗下节间长和穗叶距均高于高值亲本鲁麦18。自然降雨条件下,抽穗度相差不大,在灌溉和全部环境条件下高于高值亲本鲁麦18。泰山22号的有效小穗数、穗粒数和千粒重在自然降雨条件下介于二者之间,在灌溉和全部环境条件下显著高于高值亲本鲁麦18,呈明显的超亲遗传。泰山22号粒长性状在不同水分条件下均高于高值亲本鲁麦18,其粒宽和粒厚均介于两亲本之间。

图4 泰山22号及其亲本鲁麦18、鲁麦14在多个环境下的农艺性状表现Fig.4 Agronomic traits of Taishan22 and its parents in several cultivated environments
3 讨论
本研究利用小麦90K芯片获得SNP标记,绘制了小麦新品种泰山22的基因型图谱,明确了其遗传构成,发现基因组整体偏向性选择不明显,父母本对后代品种的贡献接近1:1。但A、B和D 3个亚基因组差别较大,A基因组鲁麦14的贡献(63.59%)远大于鲁麦18(36.41%),除3A、5A外,鲁麦14在其他A组染色体的遗传贡献远超过鲁麦18,尤其是6A染色体几乎整条染色体传递至子代泰山22。众所周知,A基因组在小麦进化过程中起着基础性核心作用,具有较多产量相关性状位点;而鲁麦14作为高产、稳产、适应性好的骨干亲本对子代泰山22的A基因组具有很大的贡献,可能是泰山22表现高产的重要基础。此外发现,泰山22在2B和5B染色体几乎全部从母本鲁麦18传递至子代,SNP位点传递率达98.38%和99.02%;6A染色体95.49%的遗传信息从鲁麦14传递至子代;这种染色体的极端偏分离现象在青农2号的遗传组成中也有发现[17]。这应与育种家对不同性状的选择息息相关,即育种家选择的性状跟这条染色体没有关系或者该染色体在育种家关心的性状方面已经达到最优。从另一个方面看,通过杂交组合产生的后代分离群体,可能还有许许多多的个体,会存在其他的整条染色体遗传现象,存在这种现象的原因有待于进一步深入分析。
小麦骨干亲本可以直接用来培育品种,由其能衍生出很多有广泛利用价值的育种材料[7,22],借助不同类型的分子标记,骨干亲本重要遗传区段研究已有较多报道[17,22-28],但多数还未将传递到衍生材料的保守基因组区段与重要农艺性状建立起直接联系,更未进入到基因的研究层面[29]。本研究虽然调查了10个环境的农艺性状数据,但也无法获得每个性状对应的染色体区段,它们之间的联系需要基于遗传群体的数量性状位点进行更深入的定位。
