全生育期干旱胁迫对‘青燕1 号’燕麦叶绿素荧光参数的影响
2022-06-11金祎婷刘文辉刘凯强梁国玲贾志锋
金祎婷,刘文辉,刘凯强,梁国玲,贾志锋
(青海省青藏高原优良牧草种质资源利用重点实验室,青海大学畜牧兽医科学院,青海 西宁 810016)
干旱是影响植物生长发育最主要的逆境胁迫因素之一[1],在我国干旱与半干旱地区,干旱又是限制作物生长及产量的重要因素[2]。近年来,由于人类活动的影响,全球气候变暖逐年增强,导致了全球或区域性气温上升以及水资源时空分布不均,干旱胁迫对农业的影响愈发严重。干旱胁迫能够对植物的外部形态、内源激素、光合作用等方面产生影响[3]。具体来说,干旱胁迫会引起植物细胞失水,破坏植物体内的水分代谢平衡[4],改变自身结构与功能,制约植物的生长发育[5],导致植物在内部生理生化特征和外部形态上发生显著变化[6]。短期或轻度干旱胁迫下植物叶片水势降低、气孔关闭,降低了CO2的摄取与光合作用;长期而严重的干旱胁迫可抑制植株生长,并引起外观形态和生物量的变化,甚至会致使植物死亡[7−9]。
叶绿素荧光参数是将植物光合作用和环境联系起来的内在指标,可以描述植物光合生理状况以及作用机理,是研究植物抗逆性指标的有效探针。干旱胁迫会使植物的叶绿素荧光参数发生变化,并降低光合电子传递链活性[10−11],破坏植物的光系统,造成光合代谢途径紊乱,使植物体内的叶绿体结构改变、叶绿素含量减少、光合作用下降[12]。关于该方面的研究也有许多,如杨文权等[13]研究了干旱对小冠花(Coronilla varia)叶绿素荧光参数的影响,发现胁迫程度和时间会影响其光合作用,严重时会使其光反应中心受损。Chen 等[14]研究发现,在短期干旱下,捕光复合体Ⅱ迅速解体,大部分的光系统蛋白在受到胁迫后,其速率会降低,尤其是PsbQ、Lhcb5 和Lhcb6 蛋白。王泳超等[15]研究了干旱胁迫下γ-氨基丁酸(GABA)保护玉米(Zea mays)幼苗光合系统的生理响应。由此可见,研究干旱胁迫对植物叶绿素荧光参数的影响可以更深入地探究植物在逆境胁迫下的反应机理,从而更好地指导生产实践。
燕麦(Avena sativa)是禾本科燕麦属一年生草本植物,其抗旱性和抗寒性较强,是适宜在青藏高原种植的高产优质、粮饲兼用作物之一[16]。燕麦种植对于维系青藏高原草牧业健康稳定发展、维持青藏高原冷季家畜饲草供给平衡以及保护高寒地区生态环境等方面具有重要意义[17]。目前,干旱已成为燕麦减产的重要因素。有关青藏高原燕麦的研究主要集中在抗倒伏性研究[17]、生产性能及品质评价[18]、干旱胁迫[19−20]、施肥、播量、播期和行距对燕麦生长特性[21]、碳氮储量[22]、种子产量[23]、光合特性以及激素含量[24]的影响等方面。国内学者在研究干旱胁迫对植物的影响方面,多侧重于探究其生理特性、产量和品质等,如刘凯强等[19−20]研究了水分胁迫对燕麦生长发育、水分利用效率及产量构成的影响,张曦[25]研究了紫花苜蓿叶性状对干旱胁迫的响应,而在干旱胁迫对植物光合性能、叶绿素荧光参数的影响方面的研究较少,且大多针对玉米(Zea mays)[26−28]、水稻(Oryza sativa)[29]、玉簪(Hosta plantaginea)[30]、山葡萄(Vitis amurensis)[31]和银杏(Ginkgo biloba)[32]等植物。另外,有关燕麦干旱胁迫与光合的研究同样很少,且这些研究多集中于某一个生育时期[33−37],但干旱胁迫对燕麦不同生育时期的影响应该是有所差异的。鉴于此,本试验结合青藏高原燕麦现状,以’青燕1 号’燕麦(A. sativacv. Qingyan No.1)为材料。‘青燕1 号’燕麦(A. sativacv. Qingyan No.1)是一个粮饲兼用优良新品种,有较强的适应能力、根系发达、产种和产草量高等特点,是由青海省畜牧兽医科学院通过杂交育种技术所育成的,现已成为青海省本地的主推品种。本试验拟研究‘青燕1 号’在不同生育时期干旱胁迫后叶绿素荧光参数的变化,明确其需水关键期,探究其作用机理,以期为青藏高原燕麦抗旱育种和节水高产栽培提供理论指导。
1 材料与方法
1.1 供试材料
试验选用青海省本地推广品种−青燕1 号,选择籽粒饱满、质地均匀且大小一致的燕麦种子。试验材料由青海省畜牧兽医科学院种质资源库提供。
1.2 试验设计和方法
试验于2019年在青海省畜牧兽医科学院的防雨棚内进行,采用盆栽方式,聚乙烯塑料盆高30 cm,直径28 cm。试验设置干旱和胁迫时期2 个处理。
干旱处理:试验设置4 个水分梯度:正常供水[CK,75% 田间持水量(75% of field water capacity,75%FWC)]、轻度胁迫(60%FWC)、中度胁迫(45%FWC)、重度胁迫(30%FWC)。
干旱胁迫时期设置4 个处理:1)苗期(seedling stage)−拔节期(jointing stage)干旱(SJ):在苗期进行干旱胁迫10 d;2)苗期−拔节期、孕穗期(booting stage)−抽穗期(heading stage)干旱(SH):分别在苗期和孕穗期进行10 d干旱胁迫;3)苗期−拔节期、开花期(flowering stage)−乳熟期(milking stage)干旱(SF):分别在苗期和开花期进行10 d 干旱胁迫;4)苗期−拔节期、孕穗期−抽穗期、开花期−乳熟期干旱(SM):分别在苗期、孕穗期和开花期进行10 d 干旱胁迫。在全生育期分别做轻度胁迫、中度胁迫和重度胁迫为对照处理(CK1),同时设置全生育期正常供水,作为计算各指标相对值的参考。
盆栽土壤取自大田土,25% 土壤含水量。将土壤晒干后混合均匀,等量装入聚乙烯塑料盆中,施加(NH4)2HPO4(含18% N,46% P2O5)38.98 g·m−2和尿素(含46% N)19.49 g·m−2作为底肥。在育苗盘中,选择健壮、大小与长势相同的幼苗,移栽至聚乙烯塑料盆中,在三叶期每盆定苗25 株,选择长势一致的植株进行干旱胁迫试验,具体处理见表1,共13 个处理,10 次重复,共130 盆,随机排列。每天17:00 采用称重差值法补充聚乙烯塑料盆缺失的水分(10 d),以控制水分处理,即通过称塑料盆的重量来计算预计灌水量之间的差值。

表1 干旱胁迫试验处理代码Table 1 Treatments code of drought stress
1.3 测定指标
采用Li-6800 便携式光合仪(LI-COR,USA),在生育期内所有干旱胁迫处理结束后,于9:00−11:30 和14:00−16:30 期间进行叶绿素荧光参数的测定,在黑暗条件下适应1 h 后,选择3 株长势一致旗叶,每个叶片重复3次,饱和脉冲光设为8000 μmol·m−2·s−1,待参数稳定后,读取暗适应下的初始荧光(F0)、暗适应下最大荧光(Fm)、光适应下的初始荧光(F0′)、光适应下最大荧光(Fm′)、稳态荧光(Fs)、表观电子传导速率(ETR)、可变荧光(Fv)。
根据Demmig-Adams 等[38]的方法计算光系统Ⅱ(PSⅡ)的最大光能转化效率(Fv/Fm)、PSⅡ电子传递量子效率(ФPSⅡ)、光化学猝灭系数(qP)、非光化学猝灭系数(NPQ)、PS Ⅰ激发能分配系数(α)、PSⅡ激发能分配系数(β)、非调节性能量耗散[Y(NO)]、调节性能量耗散[Y(NPQ)]。计算公式如下:
(1)光系统Ⅱ(PSⅡ)的最大光能转化效率(Fv/Fm) =(Fm−F0)/Fm
(2)PSⅡ电子传递量子效率(ФPSII)=(Fm′−Fs)/Fm′
(3)光化学猝灭系数(qP)=(Fm′−Fs)/(Fm′−F0′)
(4)非光化学猝灭系数(NPQ)=(Fm−Fm′)−1
(5)PSⅠ激发能分配系数(α)=(Fm−Fs)/(2Fm−F0−Fs)
(6)PSⅡ激发能分配系数(β)=(Fm−F0)/(2Fm−F0−Fs)
(7)非调节性能量耗散[Y(NO)]=1/[NPQ+1+qP×F0′/Fs×(Fm/F0−1)]
(8)调节性能量耗散[Y(NPQ)]=1 −ΦPSⅡ−1/[NPQ+1+qP×F0′/Fs×(Fm/F0−1)]
1.4 数据分析
采用Microsoft Excel 2016 软件进行数据整理。用IBM SPSS 25.0 统计软件对不同生育时期的数据进行单因素方差分析(one-way ANOVA)、相关性分析,Duncan 多重比较采用最小显著差异法(LSD),显著性水平设定为0.05。采用Origin 2021 软件绘图,胁迫时期和胁迫程度方差分析作图是用R 语言ggpubr 包实现的。数据结果采用平均值±标准差(mean±SD)的形式表示。
2 结果与分析
2.1 干旱胁迫对‘青燕1 号’F0、Fm、F0′和Fm′的影响
干旱胁迫的程度和时期对‘青燕1 号’燕麦F0、Fm、F0′和Fm′影响的方差分析见表2。从表中可以看出,胁迫程度(Sd)、胁迫时期(Sp)和二者互作(Sd×Sp)对Fm和Fm′的影响差异达到极显著水平(P<0.01);F0和F0′在胁迫程度(Sd)和胁迫程度与胁迫时期互作(Sd×Sp)下达到极显著差异水平(P<0.01),胁迫时期(Sp)对其的影响差异表现为显著水平(P<0.05)。各因素对F0、F0′和Fm′影响大小表现为:胁迫程度和胁迫时期互作(Sd×Sp)>胁迫程度(Sd)>胁迫时期(Sp);对Fm的影响大小表现为:胁迫程度和胁迫时期互作(Sd×Sp)>胁迫时期(Sp)>胁迫程度(Sd)。
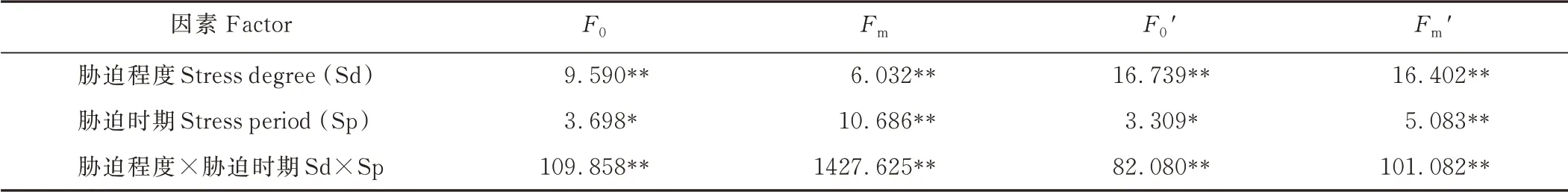
表2 干旱胁迫对‘青燕1 号’燕麦F0、Fm、F0′和Fm′影响的方差分析Table 2 Variance analysis of the F0,Fm,F0′and Fm′of‘Qingyan No.1’oat under drought stress
不同胁迫程度和胁迫时期对燕麦F0、Fm、F0′和Fm′的影响见表3 和图1。从图表中可以看出,在同一胁迫程度下,随着胁迫时期的变化(即SJ、SH、SF、SM)F0、Fm、F0′和Fm′变化存在差异。由表3 可知,在60%FWC 下,F0、Fm以SJ 时 期 最高,SF 时 期最 低;F0′以SJ 时 期 最高,SM 时 期 最 低;Fm′以SH 时 期 最高,SF 时 期最 低。在45%FWC 下,F0以SF 时期最高,SH 时期最低;Fm和F0′以SF 时期最高,SM 时期最低;Fm′以SF 时期最高,SM 时期最低。在30%FWC 下,F0以SJ 胁迫时期最高,SH 时期最低;Fm以SJ 胁迫时期最高,SM 胁迫时期最低;F0′和Fm′以SJ 时期最高,SM 时期最低。
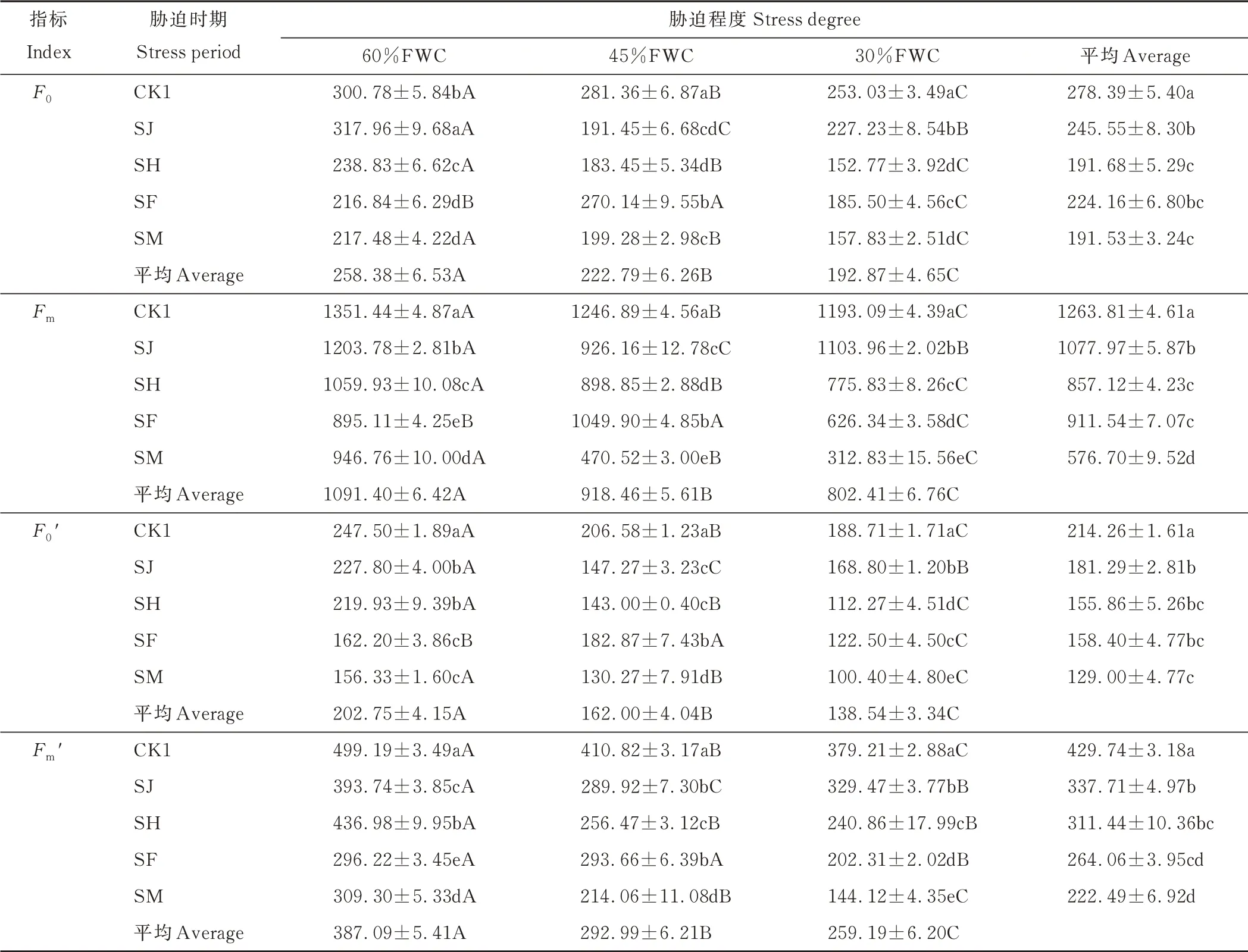
表3 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦F0、Fm、F0′和Fm′的影响Table 3 Effects of different stress degrees and stress periods on F0,Fm,F0′and Fm′of‘Qingyan No.1’oat
在同一胁迫时期下,随着胁迫程度的增加,各指标有增有减(图1)。在干旱胁迫处理的4 个时期,随着胁迫程度的增加,F0、Fm、F0′和Fm′总体均呈下降趋势。在SJ 时期,随着胁迫程度增加,F0、Fm、F0′和Fm′均先降低后逐渐增大,以45%FWC 最低,60%FWC 最高;在SH 和SM 时期,随着胁迫程度增加,F0、Fm、F0′和Fm′均逐渐降低,以30%FWC 最低,60%FWC 最 高;在SF 时 期,随着胁 迫程度增加,F0、Fm和F0′均先增大后降 低,以30%FWC 最低,45%FWC 最高,Fm′总体呈下降趋势,以30%FWC 最低。由表3 可知,在SH、SF 和SM 时期,四者均以30%FWC 最低,说明30%FWC 对其影响最大。根据平均值可以看出,与CK1 相比,F0、Fm、F0′和Fm′均在SM 时期,受到的影响最为显著。
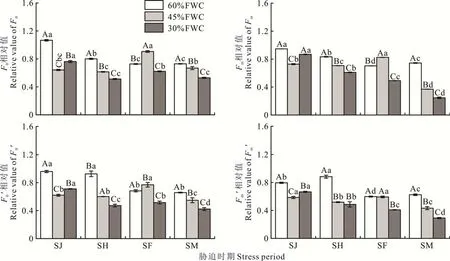
图1 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦F0、Fm、F0′和Fm′相对值的影响Fig.1 Effects of different stress degrees and stress periods on F0,Fm,F0′and Fm′of‘Qingyan No.1’oat不同小写字母表示同一胁迫程度下各胁迫时期相对值差异显著(P<0.05);不同大写字母表示同一胁迫时期下各胁迫程度相对值差异显著(P<0.05)。下同。Different lowercase letters indicate significant differences in relative values of different stress period under the same stress degree at the 0.05 level. Different capital letters indicate significant differences in relative values of different stress degree under the same stress period at the 0.05 level.The same below.
2.2 干旱胁迫对‘青燕1 号’Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 的影响
干旱胁迫的程度和时期对’青燕1 号’燕麦Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 相对值影响的方差分析见表4。从表中可以看出,胁迫程度(Sd)、胁迫时期(Sp)和二者互作(Sd×Sp)对ΦPSⅡ和ETR 的影响差异达到极显著水平(P<0.01);Fs在胁迫程度(Sd)和胁迫程度与胁迫时期互作(Sd×Sp)下达到极显著差异水平(P<0.01),Fv和Fv/Fm在胁迫时期(Sp)和胁迫程度与胁迫时期互作(Sd×Sp)下达到极显著差异水平(P<0.01)。各因素对Fs影响大小表现为:胁迫程度和胁迫时期互作(Sd×Sp)>胁迫程度(Sd)>胁迫时期(Sp);对Fv、Fv/Fm、ΦPSⅡ和ETR 的影响大小表现为:胁迫程度和胁迫时期互作(Sd×Sp)>胁迫时期(Sp)>胁迫程度(Sd)。

表4 干旱胁迫对‘青燕1 号’燕麦Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 影响的方差分析Table 4 Variance analysis of the Fs,Fv,Fv/Fm,ΦPSⅡand ETR of‘Qingyan No.1’oat under drought stress
不同胁迫程度和胁迫时期对燕麦Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 的影响见表5 和图2。从图2 中可以看出,各指标相对值均低于1.0,说明干旱胁迫处理的抑制效应明显,但其效应大小因胁迫时期和胁迫程度而异。由表5 可知,在同一胁迫程度下,随着胁迫时期的变化,Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 变化存在差异,但整体呈下降趋势。在60%FWC 下,Fs以SH 时期最高,SF 时期最低;Fv和Fv/Fm以SJ 时期最高,SF 时期最低;ΦPSⅡ无明显变化;ETR 以SH 时期最高,SF 时期最低。在45%FWC 下,Fs和Fv以SF 时期最高,SM 时期最低;Fv/Fm以SJ 和SH 时期均最高,SM 时期最低;ΦPSⅡ和ETR 以SJ 时期最高,ΦPSⅡ以SF 和SM 时期均最低,ETR 以SF 时期最低。
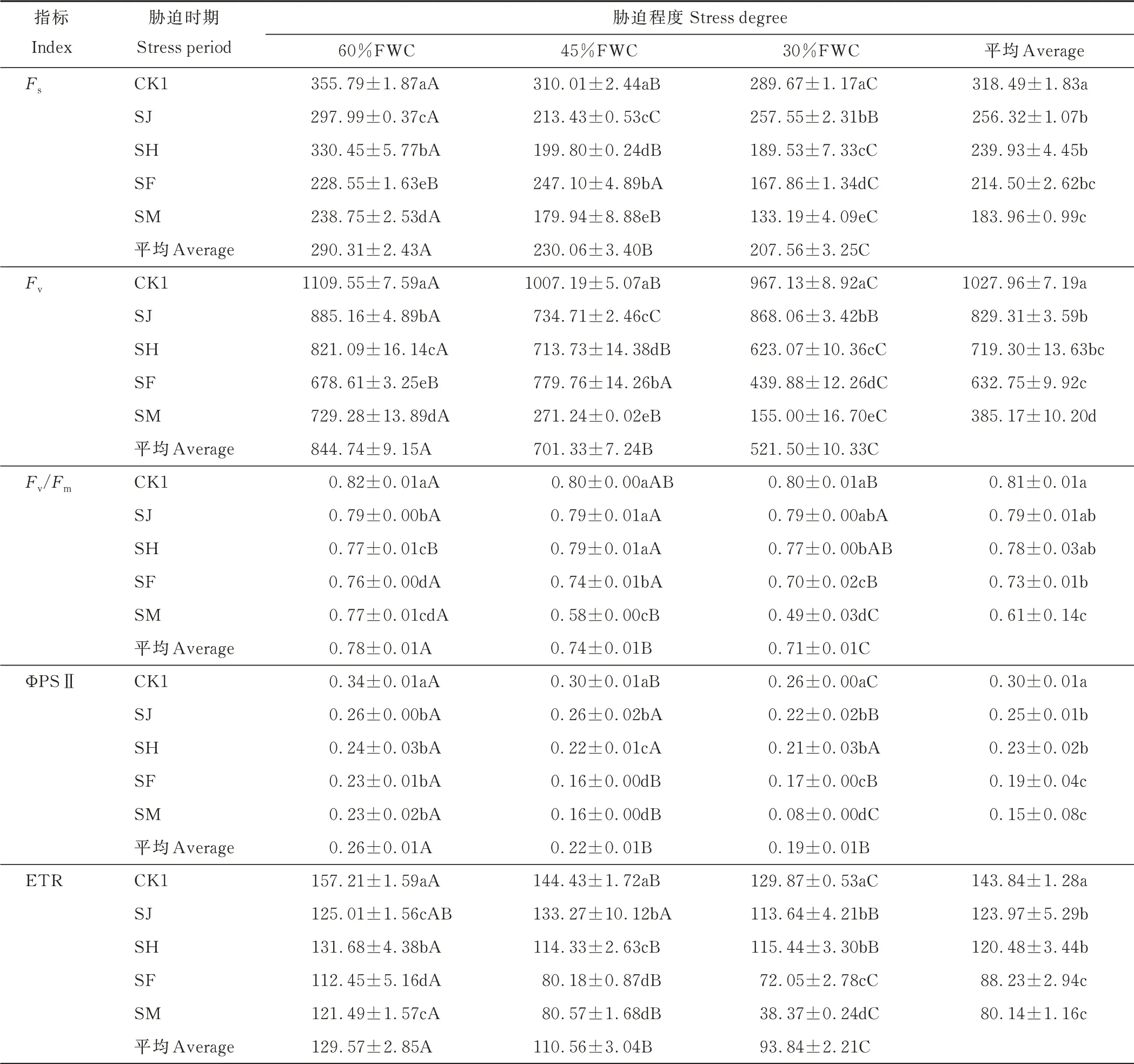
表5 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 的影响Table 5 Effects of different stress degrees and stress periods on Fs,Fv,Fv/Fm,ΦPSⅡand ETR of‘Qingyan No.1’oat
由表5 和图2 可知,在同一胁迫时期下,随着胁迫程度的增加,各指标总体呈下降趋势。在SJ 时期,随着胁迫程度的增加,Fs和Fv先降低后逐渐增大,以45%FWC 最低;ΦPSⅡ和ETR 先升高后逐渐降低,以30%FWC 最低,Fv/Fm无明显变化,处理间无显著差异(P>0.05)。在SH 时期,随着干旱胁迫程度的增加,各指标总体呈下降趋势,Fs、Fv、Fv/Fm和ΦPSⅡ以30%FWC 最低,ETR 以45%FWC 最低。在SF 时期,Fs和Fv/Fm先增大后逐渐降低,以30%FWC 最低,45%FWC 最高;ΦPSⅡ总体呈下降趋势,以45%FWC 最低,ETR 逐渐降低,以30%FWC 最低。在SM 时期,Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 均逐渐降低,以30%FWC 最低。根据平均值可以看出,与CK1 相比,Fs、Fv、Fv/Fm、ΦPSⅡ和ETR 均在SM 时期,受到的影响最为显著(表5)。

图2 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦Fs、Fv/Fm、ΦPSⅡ和ETR 的影响Fig.2 Effects of different stress degrees and stress periods on Fs,Fv/Fm,ΦPSⅡ,and ETR of‘Qingyan No.1’oat
2.3 干旱胁迫对‘青燕1 号’qP 和NPQ 的影响
干旱胁迫的程度和时期对’青燕1 号’燕麦qP影响的方差分析见表6。从表中可以看出,胁迫程度(Sd)、胁迫时期(Sp)和二者互作(Sd×Sp)对qP的影响差异达到极显著水平(P<0.01);而NPQ 仅在胁迫时期(Sp)和胁迫程度与胁迫时期互作(Sd×Sp)下达到极显著差异水平(P<0.01)。各因素对qP的影响大小表现为:胁迫程度(Sd)>胁迫时期(Sp)>胁迫程度和胁迫时期互作(Sd×Sp),对NPQ 的影响大小表现为:胁迫程度和胁迫时期互作(Sd×Sp)>胁迫时期(Sp)>胁迫程度(Sd)。

表6 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦qP、NPQ、Y(NO)、Y(NPQ)、α 和β 的影响Table 6 Effects of different stress degrees and stress periods on qP,NPQ,Y(NO),Y(NPQ),α and β of‘Qingyan No.1’oat
不同胁迫程度和胁迫时期对‘青燕1 号’燕麦qP和NPQ 相对值的影响见图3。从图中可以看出,同一胁迫程度下,随着胁迫时期的变化,qP和NPQ 的相对值变化存在差异。在60%FWC 下,qP相对值以SJ 时期最高,SM 时期最低;NPQ 相对值以SM 时期最高,SH 时期最低。在45%FWC 下,qP相对值以SJ 时期最高,SM 时期最低;NPQ 相对值以SF 时期最高,SM 时期最低。在30%FWC 下,qP和NPQ 相对值均以SJ 时期最高,SM 时期最低。

图3 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦qP 和NPQ 的影响Fig.3 Effects of different stress degrees and stress periods on qP and NPQ of‘Qingyan No.1’oat
在同一胁迫时期下,随着胁迫程度增加,qP和NPQ 相对值有增有减。在SJ 时期,随着胁迫程度的增加,qP相对值逐渐降低,以30%FWC 最低,NPQ 相对值则相反,以60%FWC 最低。在SH 时期,qP和NPQ 相对值先增大后逐渐降低,其中qP相对值以30%FWC 最低,NPQ 相对值以60%FWC 最低。在SF 时期,qP相对值先降低后逐渐增大,以45%FWC 最低;NPQ 相对值先增大后逐渐降低,以60%FWC 最低。
2.4 干旱胁迫对‘青燕1 号’Y(NO)和Y(NPQ)的影响
干旱胁迫的程度和时期对‘青燕1 号’燕麦Y(NO)和Y(NPQ)影响的方差分析见表6。从表中可以看出,胁迫时期(Sp)和胁迫程度与胁迫时期互作(Sd×Sp)对Y(NO)和Y(NPQ)的影响差异达到极显著水平(P<0.01);而Y(NO)和Y(NPQ)在胁迫程度(Sd)下均未达到显著差异水平(P>0.05)。各因素对Y(NO)和Y(NPQ)的影响大小表现为:胁迫程度和胁迫时期互作(Sd×Sp)>胁迫时期(Sp)>胁迫程度(Sd)。
不同胁迫程度和胁迫时期对‘青燕1 号’燕麦Y(NO)和Y(NPQ)的影响见图4。同一胁迫程度下,随着胁迫时期的变化,Y(NO)和Y(NPQ)的相对值变化存在差异。在60%FWC 下,Y(NO)相对值以SH 时期最高,SJ 时期最低;Y(NPQ)相对值以SM 时期最高,SH 时期最低。在45%FWC 下,Y(NO)相对值以SM 时期最高,SH 时期最低;Y(NPQ)相对值以SF 时期最高,SM 时期最低。在30%FWC 下,Y(NO)相对值以SM 时期最高,SJ 时期最低;Y(NPQ)相对值以SF 时期最高,SM 时期最低。
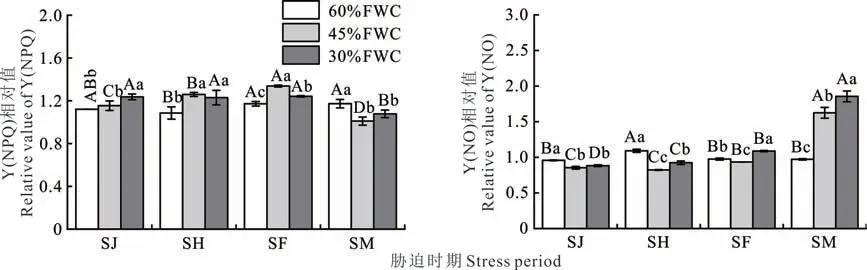
图4 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦Y(NO)和Y(NPQ)的影响Fig.4 Effects of different stress degrees and stress periods on Y(NO)and Y(NPQ)of‘Qingyan No.1’oat
在同一胁迫时期下,随着胁迫程度增加,Y(NO)和Y(NPQ)的相对值有增有减。在SJ 时期,随着胁迫程度增加,Y(NO)相对值先降低后逐渐增大,以45%FWC 最低;Y(NPQ)相对值逐渐增大,以60%FWC 最低,30%FWC 最高。在SH、SF 时期,随着胁迫程度增加,Y(NO)相对值先降低后逐渐增大,以45%FWC 最低,在SH 时期,Y(NO)相对值以60%FWC 最高,在SF 时期,其以30%FWC 最高;Y(NPQ)相对值先增大后逐渐降低,以60%FWC 最低,45%FWC 最高。在SM 时期,随着胁迫程度增加,Y(NO)相对值逐渐增大,以60%FWC 最低,30%FWC 最高;Y(NPQ)相对值先降低后逐渐增大,以45%FWC 最低,60%FWC 最高。
2.5 干旱胁迫对‘青燕1 号’燕麦α 和β 的影响
不同干旱胁迫程度对不同生育时期‘青燕1 号’燕麦α 和β 影响的方差分析见表6。从表中可以看出,胁迫程度(Sd)、胁迫时期(Sp)和两者互作(Sd×Sp)均对α 和β 的影响差异达到极显著水平(P<0.01)。各因素对α 和β的影响大小表现为:胁迫程度(Sd)>胁迫程度和胁迫时期互作(Sd×Sp)>胁迫时期(Sp)。
不同胁迫程度和胁迫时期对‘青燕1 号’燕麦α 和β 的影响见图5。同一胁迫程度下,随着胁迫时期的变化,α和β 的相对值变化存在差异。在60%FWC 下,α 相对值以SJ 时期最高,SM 时期最低;β 相对值则相反,以SM 时期最高,SJ 时期最低。在45%FWC 下,α 相对值以SJ 时期最高,SM 时期最低;β 相对值以SM 时期最高,SJ 时期最低。在30%FWC 下,α 相对值以SJ 时期最高,SM 时期最低;β 相对值相反,以SM 时期最高,SJ 时期最低。
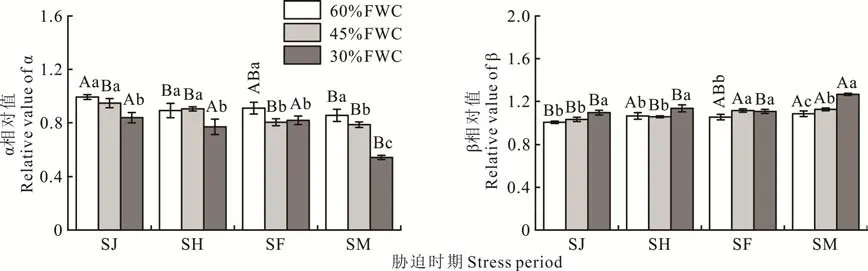
图5 不同胁迫程度和胁迫时期对‘青燕1 号’燕麦α 和β 的影响Fig.5 Effects of different stress degrees and stress periods on α and β of‘Qingyan No.1’oat
同一胁迫时期下,随着胁迫程度增加,α 和β 的相对值有增有减。在SJ 时期,随着胁迫程度增加,α 相对值逐渐降低,以30%FWC 最低;β 相对值逐渐增大,以60%FWC 最低,30%FWC 最高。在SH 时期,随着胁迫程度增加,α 相对值先增大后逐渐降低,以30%FWC 最低,45%FWC 最高;β 相对值先降低后逐渐增大,以45%FWC 最低,30%FWC 最高。在SF 时期,随着胁迫程度增加,α 相对值先降低后逐渐增大,以45%FWC 最低,60%FWC最高;β 相对值先增大后逐渐降低,以60%FWC 最低,45%FWC 最高。在SM 时期,α 相对值逐渐降低,以30%FWC 最低,60%FWC 最高;β 相对值逐渐增大,以60%FWC 最低,30%FWC 最高。
2.6 干旱胁迫对‘青燕1 号’燕麦荧光参数的影响
干旱胁迫对‘青燕1 号’燕麦荧光参数的影响见图6 和表7,图6 是利用R 语言ggpubr 包实现的,表7 是根据上述各图表数据统计分析而来,以便更直观地观察各荧光参数的变化。根据图表可看出,在SM 和SF 时期对燕麦进行干旱胁迫处理对其荧光参数影响最大,且干旱胁迫处理后,燕麦的Y(NO)、NPQ、Y(NPQ)和β 整体呈上升趋势,而Fv、Fm、ΦPSⅡ、ETR、Fm′、Fs、F0′、qP、F0、α 和Fv/Fm整体呈下降趋势。与CK 相比,β、Fv、Fm、ΦPSⅡ、ETR、Fm′、Fs、F0′、qP、α 和Fv/Fm均 在T13(SM,30%FWC)处 理 下 受 到 的 影 响 最 大,且β 与CK 相 比 增 加 了26.60%,Fv、Fm、ΦPS Ⅱ、ETR、Fm′、Fs、F0′、qP、α 和Fv/Fm分 别 降 低 了84.86%、75.41%、75.03%、75.00%、70.89%、61.38%、57.73%、57.06%、45.61%和38.50%;Y(NO)和Y(NPQ)在T9(SF,45%FWC)处理下,受到的影响最大,与CK 相比,分别增加了85.52%和33.76%;NPQ 在T12(SM,45%FWC)处理下,受到的影响最大,与CK 相比增加了65.21%;F0在T7(SH,30%FWC)处理下,受到的影响最大,与CK 相比降低了48.86%。

表7 干旱胁迫对燕麦荧光参数的影响Table 7 Effects of drought stress on fluorescence parameters of‘Qingyan No.1’oat
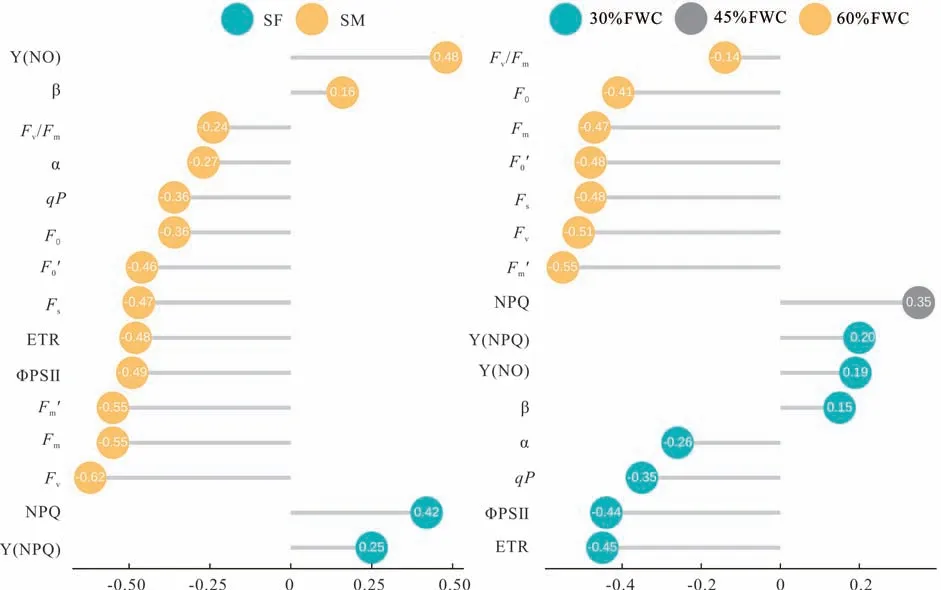
图6 胁迫时期(左)和胁迫程度(右)对‘青燕1 号’燕麦荧光参数的影响Fig.6 Effects of stress period(left)and stress degree(right)on fluorescence parameters of‘Qingyan No. 1’oat
3 讨论
苗期是牧草生长的关键时期,此时幼苗养分供给与耗散平衡正处于稳定性较差的阶段,在苗期,燕麦主要通过协调光合产物,优先给根系提供养分,以此响应干旱胁迫;在分蘖期干旱,燕麦通过降低根系的整体生长速度来响应干旱胁迫;在拔节期干旱,燕麦主要通过调节有机物质,即多分配给地上部分,来响应干旱胁迫[33]。干旱胁迫使植物叶片含水量降低,破坏了其光合机构,从而导致叶绿素荧光发生变化[32]。而叶绿素荧光参数可以间接反映植物的光合作用能力[39],因为正常情况下,叶绿体吸收的光能主要通过3 种途径来消耗,且三者之间联系密切,即叶绿素荧光、光合电子传递和热耗散[40]。有研究表明,植物在许多逆境胁迫中最先受到伤害的部位是叶片的PSⅡ[41−42],通过对其荧光动力学测定,可以分析植物光合机构的变化。
暗适应下的初始荧光(F0)代表不参与光系统Ⅱ(PSⅡ),处于完全开放时的荧光产量,可以反映PSⅡ部分反应中心是否失活,若值变大,则表明PSⅡ反应中心被破坏;暗适应下最大荧光(Fm)是PSⅡ反应中心在完全关闭时的荧光产量,可以反映通过PSⅡ的电子传递情况,若其值变小,则表明光合作用受到抑制;PSⅡ的最大光能转化效率(Fv/Fm)表示PSⅡ原初光化学效率,主要用来反映PSⅡ复合物的光抑制是否受到伤害,在正常生长情况下,Fv/Fm值较稳定,若值降低,则表明发生了光抑制,从而使光合活性降低;PSⅡ电子传递量子效率(ФPSⅡ)表示实际光化学效率,反映了光照条件下PSⅡ反应中心在部分关闭情况下的实际原初光能捕获效率,其值大小与下游电子传递、反应中心激发能捕获效率和开放程度有关[42];表观电子传导速率(ETR)反映实际光照强度下的光合能量传递速率[39]。许多研究都表明干旱胁迫会使Fs、Fv、Fm、Fv/Fm、ΦPSⅡ和ETR 显著降低,F0升高[32−44]。本研究发现,干旱胁迫显著降低了不同生育时期‘青燕1 号’燕麦Fv/Fm和ΦPSⅡ的相对值,并随着干旱胁迫程度的加剧二者降低幅度变大(表5),这是由于PSⅡ结构及其生理状态受到破坏,导致PSⅡ的最大光能转化效率Fv/Fm在后期大幅度降低。Fm和ETR 也显著降低(表3 和表5),反映了PSⅡ反应中心光合潜能的下降,这可能是因为干旱胁迫使叶绿体类囊体膜上的蛋白复合体部分失活所引起的,在Fv/Fm降低的同时,伴有F0的整体下降(图1和图2),表明干旱胁迫引起PSⅡ反应中心的光抑制损伤,尤其是在30%FWC,由于叶片失水,导致‘青燕1 号’燕麦叶片PSⅡ结构被破坏,使得光合电子由PSⅡ反应中心向下游电子传递受到抑制,大大降低了原初光能捕获效率。在本研究中,Fv/Fm相对值在SM 时期、45%FWC 下降最快,在SF 时期、30%FWC 次之;ΦPSⅡ相对值在SM时期、30FWC%下降最快,在SF 时期、45%FWC 次之,且随着胁迫程度的增加,两者整体上均以30%FWC 最低(图2)。因此,本研究得出,Fv/Fm和ΦPSⅡ以SM 时期影响最大,SF 时期次之,30%FWC 对其影响最大。
光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)可以反映过剩光能对光合系统产生抑制的程度和清除能力[20],qP可以代表光下PSⅡ的开放程度,反映了PSⅡ吸收的光能用于光化学电子传递的比例,值越大,表明PSⅡ反应中心的电子传递就越活跃,植物吸收的光能中用于光合电子传递的比例就越高,反之,值越小,表明原初电子受体的氧化还原程度越高[45]。本试验中,胁迫程度对‘青燕1 号’燕麦NPQ 无显著影响,qP受胁迫程度和时期以及二者互作的影响达到极显著差异水平(表6),干旱胁迫使qP和NPQ 的相对值发生显著变化,随着干旱胁迫程度的加剧,qP总体呈降低趋势,NPQ 总体呈升高趋势,qP相对值以SM 时期影响最大,SH 时期次之;NPQ 相对值以SH 时期影响最大,SM 时期次之(图3),这表明干旱胁迫降低了PSⅡ运转效率,同时产生的光抑制增强了过剩光能的天线热耗散。
非光化学猝灭系数(NPQ)主要分为非调节性能量耗散Y(NO)和调节性能量耗散Y(NPQ),二者是从qP和NPQ 的另一个角度来反映PSⅡ的光化学效率。Y(NO)是光损伤的负评价指标,反映了植物光合系统的非调节性量子产量;Y(NPQ)反映了植物不能用于光合电子传递的多余光能,而是采用热耗散的形式,是植物重要的自我保护方式之一[46]。在SJ 和SH 时期,随着胁迫程度的加剧,Y(NO)相对值整体呈下降趋势,在SF 和SM 时期,其相对值整体呈上升趋势,以SM 时期上升幅度最大(图4),表明‘青燕1 号’燕麦在SJ 和SH 胁迫时期通过自身调节机制增强了光能消耗能力,保护了光合系统,而在SF 和SM 时期,已经达到光能消耗极限,过剩的光能可能传递给O2,形成具有极大破坏性的单线态氧,从而使光合机构失活。相反,在SJ、SH 和SF 时期胁迫时,随着胁迫程度的加剧,Y(NPQ)相对值总体呈上升趋势,在SM 时期其总体呈下降趋势,该时期对燕麦Y(NPQ)的影响最大(图4),表明在胁迫初期,‘青燕1 号’燕麦通过自身调节机制,即快速消耗光能,来减轻光合系统的损伤,从而适应干旱环境。在60%FWC 和45%FWC 胁迫下,‘青燕1 号’燕麦Y(NPQ)相对值逐渐增大,30%FWC 胁迫下,其值在SM 时期降低幅度最大,表明‘青燕1 号’燕麦在60%FWC 和45%FWC 胁迫下,其仍具有一定的热耗散调节能力,但在30%FWC 胁迫下,已很难适应干旱胁迫环境,不能正常进行光能热耗散。因此,Y(NO)和Y(NPQ)均以SM 时期影响最大。
光合电子高效运转和协调传递的保证使植物PSⅠ和PSⅡ激发能均匀分配[47]。PSⅡ激发能分配系数(β)显著增加会诱导反应中心可逆失活,可能损坏类囊体膜结构和PSⅡ,从而阻碍光合电子的传递,降低光合作用的效率[48]。本研究得出,‘青燕1 号’燕麦在不同生育期干旱胁迫下分配给PSⅠ的激发能α 显著降低,而分配给PSⅡ的激发能β 显著增加,且在SM 时期两者的变化幅度最大(图5),说明干旱胁迫导致激发能在PSⅠ和PSⅡ间的分配失衡,破坏了‘青燕1 号’燕麦的光合电子传递。故α 和β 以30%FWC 和SM 时期影响最大,即重度胁迫和生育期持续干旱对两者的影响最为显著。
4 结论
1)‘青燕1 号’燕麦不同生育时期胁迫整体影响表现为:苗期−乳熟期干旱(SM)>苗期−开花期干旱(SF)>苗期−抽穗期干旱(SH)>苗期−拔节期干旱(SJ)。
2)‘青燕1 号’燕麦不同胁迫程度整体影响大小表现为:重度胁迫(30%FWC)>中度胁迫(45%FWC)>轻度胁迫(60%FWC)。
3)在重度胁迫(30%FW)和苗期−乳熟期干旱(SM)时,‘青燕1 号’燕麦的叶绿素荧光参数受到的影响最大;在轻度胁迫(60%FWC)和苗期−拔节期干旱(SJ)时,其受到的影响最小。
4)各叶绿素荧光参数以不同程度增加或降低,Y(NO)、NPQ、Y(NPQ)和β 整体呈上升趋势,Fv、Fm、ΦPSⅡ、ETR、Fm′、Fs、F0′、qP、F0、α 和Fv/Fm整体呈下降趋势。
