青藏高原高寒草地土壤微生物量碳氮含量特征及其控制要素
2022-06-11李洋王毅韩国栋孙建汪亚峰
李洋,王毅,韩国栋,孙建,汪亚峰
(1. 内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010011;2. 中国科学院青藏高原研究所青藏高原地球系统科学国家重点实验室,北京 100101;3. 中山大学生命科学学院,广东 广州 510275)
土壤微生物对生态系统功能具有重要影响,几乎可以驱动所有的土壤生化过程[1−3]。土壤微生物量是指土壤中体积小于5×103μm3的生物总量,但不包括活的植物有机体,其中土壤微生物生物量碳(microbial biomass carbon,MBC)和土壤微生物生物量氮(microbial biomass nitrogen,MBN)是有机碳氮和无机碳氮之间转换的枢纽,在生态系统碳氮循环中起着重要的作用[4−5]。此外,MBC 和MBN 是评价土壤质量和健康状况不可缺少的重要生物学指标[6],也是土壤有效养分的重要来源,与植物根部营养密切相关[7]。因此,开展对土壤MBC 和MBN含量特征及其影响因素的研究,对于认识和研究土壤中的微生物以及生态系统的维持机制具有重要意义。
草地作为陆地生态系统的重要组成部分[8−10],研究气候等环境因子对土壤微生物的影响,已经成为该领域研究的热点[11−12]。研究表明,气候因子(如降水和温度)、生物因子(如群落多样性和生物量)和土壤理化性质(如土壤温度和营养水平)对土壤微生物量的含量都有一定的影响[13−15],但是目前对土壤微生物量与其影响因子之间存在何种关联还没有统一的结论[16]。例如,气候因子方面,在贺兰山地区MBC 和MBN 均与年均降水量呈显著正相关[17];而在黄土高原西北部地区,年均降水量对MBC 的影响并不显著[18]。生物因子方面,有研究表明不同的植物功能群也会对土壤微生物量造成不同的影响[19]。土壤理化性质方面,由于地域性差异明显,东北羊草(Leymus chinensis)草原在一定范围内土壤微生物量与pH 呈指数负相关[20];而在贺兰山地区土壤微生物量与pH 呈显著线性负相关[21]。由此可见,生物因子和非生物因子对土壤微生物量含量的影响有可能在不同的地理区域存在着不同的变化规律和主导因子。特别是在高寒草地,由于其独特的地理环境和气候条件,导致土壤MBC 和MBN 对环境因素存在着不同的响应机制。
青藏高原是地球上海拔最高,面积最广的高寒地区,生态系统异常敏感,是全球气候和生态环境变化的指示器和调节器[22]。高寒草甸和高寒草原是青藏高原两种主要的草地类型,占青藏高原总面积的60%以上,在维持青藏高原生态系统功能方面起着至关重要的作用[23−24]。青藏高原由于其独特的地理环境和气候敏感性,是研究土壤微生物量碳氮含量特征及其影响因素的理想区域。本研究通过对青藏高原高寒草甸和高寒草原样带调查,研究了两种草地类型MBC 和MBN 含量与气候、植物群落多样性以及土壤理化性质等之间的关系,并且探究造成两种草地类型微生物量差异的主要因素,以期为青藏高原高寒草地生态系统可持续管理提供理论参考。
1 材料与方法
1.1 研究区概况
青藏高原(80°−105° E,27°−37° N)主要包括四川省西北部、青海和西藏,总面积约257.24万km2,约占我国国土面积的1/4,平均海拔约4000 m[25]。全年气温较低,区域差异较大,降水时期高度集中,但在时间和空间上的分布极不均匀,雨季主要集中在5−9月,占到全年降水量的60%左右[26]。高寒草甸和高寒草原是青藏高原两种主要的草地类型,其中,针茅属(Stipa)植物是高寒草原主要的建群种,土壤类型以高山草原土为主,而高寒草甸则以嵩草属(Kobresia)和薹草属(Carex)植物为主要建群种[27],土壤类型以高山草甸土为主,高寒草甸年均降水量、地上生物量和物种丰富度均高于高寒草原[28]。
1.2 样品采集和植被调查
本研究于2015年7−8月开展采样工作,共采取了115 个样点,其中包括高寒草原62 个,高寒草甸53 个[25,29]。在每个样地选取10 m×10 m 的1 个大样方,在其对角线上分别设置3 组进行取样,样方大小为50 cm×50 cm,调查植被群落特征。接着,用剪刀将样方框内的植物贴近地面剪取后带回实验室用烘箱65 ℃烘干至恒重,进而测定其地上生物量[30]。用直径为5 cm 的土钻分别在每个组的小样方框内取3 钻(0~10 cm 土层),带回实验室用于测定土壤理化性质。用环刀在0~10 cm 的土壤深度进行取样,用于土壤容重的测定。所有样品采集后严格保存并带回实验室进行测定[31]。
1.3 群落和土壤理化指标
群落特征采用优势度指数(Simpson 指数,D)、多样性指数(Shannon−Wiener 指数,H′)和均匀度指数(Pielou指数,E)来计算,计算公式[32]:

式中:S为样方中的总物种数,Pi表示第i种个体数占群落中总个体数的比例,H为实际观察的物种多样性指数。
土壤含水量(soil moisture,SM)通过便携式时域反射仪(Spectrum Technologies TDR-100)现场测定,使用环刀法测定土壤容重(soil bulk density,SBD)。将野外采集的植物地上和地下部分装在纸袋中放入75 ℃烘箱中烘干至恒重,分别计算其地上生物量(aboveground biomass,AGB)和地下生物量(belowground biomass,BGB);采用Vario MICRO Cube 元素分析仪(Elementar,Hanau,德国)测定土壤全氮(soil total nitrogen,STN)和土壤有机碳(soil organic carbon,SOC)含量,分别采用重铬酸盐氧化法、碱解还原扩散法和奥尔森法测定土壤可溶性有机碳(dissolved organic carbon,DOC)、土壤速效氮(soil available nitrogen,SAN)和土壤速效磷(soil available phosphorus,SAP)[33]含量;用钼锑抗比色法测定土壤全磷(soil total phosphorus,STP)含量;采用熏蒸提取法测定微生物量碳和微生物量氮含量[34]。
1.4 气候数据来源
通过中国气象数据网(http://data. cma. cn/)获取青藏高原5−10月生长季降水量(growing season precipitation,GSP)和生长季均温(growing season temperature,GST),使用Anusplin 4.2 进行空间插值生成栅格。通过ArcGIS 10.2 软件(ESRI,Redlands,美国)进行研究区115 个样点气候数据的提取。
1.5 数据分析方法
将数据整理在Microsoft Excel 2010 中,用SPSS 25.0(IBM SPSS Statistics 25)对数据进行分析,不同草地微生物量碳(MBC)和微生物量氮(MBN)采用单因素方差分析,用Duncan 方法检验在不同草地中MBC 和MBN 差异的显著性(P<0.01);用ArcGIS 10.2 绘制研究区概况图;用Origin 2018 和R 3.6.1 绘制箱线图、主成分分析图、热度图和散点图,包括R 语言的“FactoMineR”“factoextra”“corrplot”“ggplot2”和“gridExtra”语言包;最后使用Amos Graphics(IBM SPSS Amos 24.0)分别对两种草地的MBC 和MBN 的影响因素建立结构方程模型,探究不同因子对不同草地类型土壤微生物量碳氮的影响机制。
2 结果与分析
2.1 不同草地类型土壤微生物生物量碳氮特征
高寒草甸和高寒草原MBN 含量的平均值分别为185.2 和70.4 mg·kg−1,且高寒草甸总体水平高于高寒草原;两种草地类型MBC 含量平均值分别为637.8 和194.9 mg·kg−1,且高寒草甸总体水平也高于高寒草原,高寒草原的MBN 和MBC 含量分布相对稳定,而高寒草甸的MBC 和MBN 含量分布相对离散。单因子方差分析结果显示,不同草地类型MBC 和MBN 含量差异显著(P<0.01,图1)。

图1 不同草地类型土壤微生物生物量碳氮含量特征Fig.1 Characteristics of soil microbial biomass carbon and nitrogen of different grassland types**表示在0.01 水平上差异显著。** indicates significant differences at the 0.01 level.
2.2 生物和非生物因子对土壤微生物量碳氮的影响
高寒草甸MBC 和MBN 含量均与GSP、SM、SAN、DOC、SAP、SOC、STP 和STN 含量呈显著正相关,且MBN 含量与Shannon−Wiener 指数呈显著正相关(P<0.01,图2a);高寒草原MBC 和MBN 含量与GSP、SM、AGB、Simpson 指数、Pielou 指数、Shannon−Wiener 指数、SAN、DOC、SOC、STP 和STN 含量呈显著正相关(P<0.01,图2b);SBD 与两种草地类型MBC 和MBN 含量均呈显著负相关(P<0.01)。

图2 高寒草甸(a)和高寒草原(b)微生物生物量碳氮、气候、群落多样性及土壤理化性质之间的相关性Fig. 2 Correlation among microbial biomass carbon and nitrogen,climate,community diversity and soil physicochemical properties in the alpine meadow(a)and the alpine steppe(b)MBC,土壤微生物生物量碳含量;MBN,土壤微生物生物量氮含量;SM,土壤含水量;SBD,土壤容重;BGB,地下生物量;AGB,地上生物量;SAN,土壤速效氮含量;STN,土壤全氮含量;SAP,土壤速效磷含量;STP,土壤全磷含量;SOC,土壤有机碳含量;DOC,土壤可溶性有机碳含量;D,Simpson 指数;E,Pielou 指数;H′,Shannon−wiener 指数。 MBC,microbial biomass carbon;MBN,microbial biomass nitrogen;SM,soil moisture;SBD,soil bulk density;BGB,belowground biomass;AGB,aboveground biomass;SAN,soil available nitrogen;STN,soil total nitrogen;SAP,soil available phosphorus;STP,soil total phosphorus;SOC,soil organic carbon;DOC,dissolved organic carbon;D,Simpson diversity index;E,Pielou’s evenness index;H′,Shannon diversity index.
2.3 气候因子对土壤微生物生物量碳氮的影响
建立两种草地GSP 和GST 与土壤微生物生物量碳氮回归模型发现,高寒草甸和高寒草原的MBC 含量与GSP 呈显著正相关(R2=0.49,P<0.01;R2=0.44,P<0.01),且高寒草原对降水的敏感性大于高寒草甸(slope 1=1.55<slope 2=1.87);两种草地类型的MBN 含量与GSP 呈显著正相关(R2=0.50,P<0.01;R2=0.35,P<0.01)。GST 对高寒草原MBN 含量的影响显著(R2=0.14,P<0.01),而对高寒草甸MBC(R2=0.03,P>0.05)、MBN 含量(R2=0.01,P>0.05)和高寒草原MBC 含量(R2=0.06,P>0.05)均无显著影响(图3 和表1)。
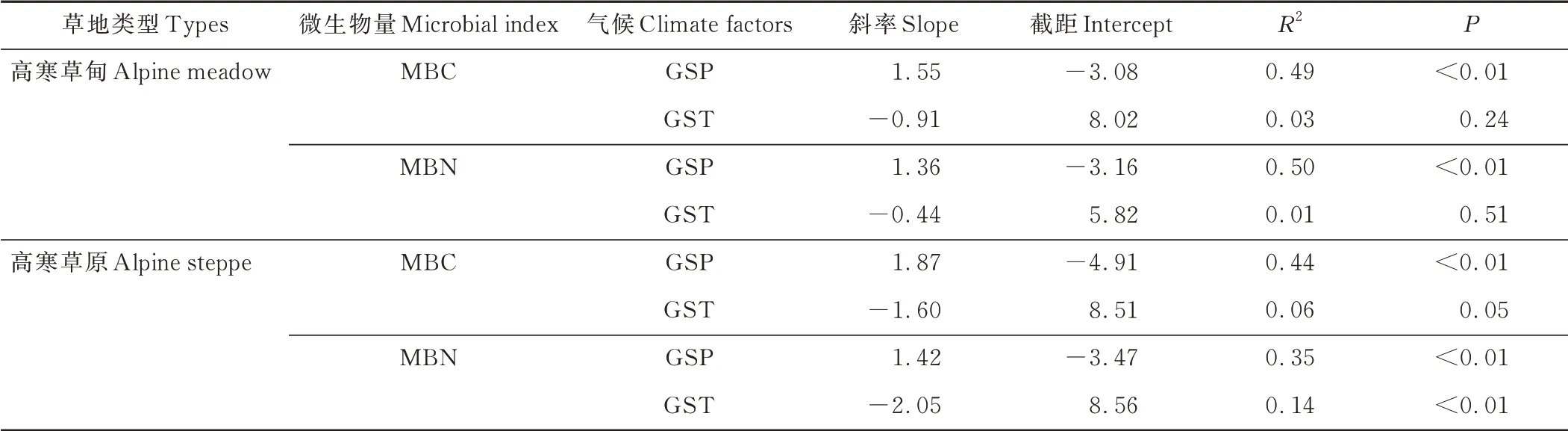
表1 不同草地类型微生物量碳氮与气候因子回归及相关性分析Table 1 Regression and correlation analysis between microbial biomass carbon and nitrogen and climate factors in different grassland types
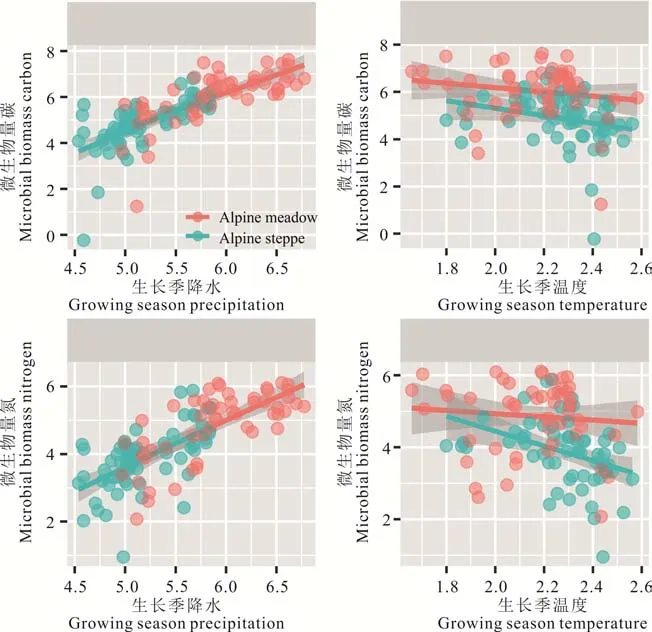
图3 ln 对数变换数据后土壤微生物量碳、氮与气候因子的关系Fig.3 The relationship between soil microbial biomass carbon/nitrogen and climate factors after ln transformation
2.4 群落和土壤因子对微生物量碳氮的影响
群落多样性指标主成分分析结果表明,在高寒草甸中第1 主成分(Dim1)和第2 主成分(Dim2)方差贡献率分别为61.7%和23.0%,且对第1 主成分(Dim1)起主要作用的因子是Simpson 指数,对第2 主成分(Dim2)起主要作用的因子是BGB(图4a);在高寒草原中第1 主成分(Dim1)和第2 主成分(Dim2)方差贡献率分别为64.5%和21.4%,且对第1 主成分(Dim1)起主要作用的因子是Simpson 指数,对第2 主成分(Dim2)起主要作用的因子是BGB(图4c)。土壤理化性质主成分分析结果表明,在高寒草甸中第1 主成分(Dim1)的方差贡献率为67.6%,且对第1 主成分(Dim1)起主要作用的因子是STN 和SOC(图4b);在高寒草原中第1 主成分(Dim1)的方差贡献率为64.9%,且对第1 主成分(Dim1)起主要作用的因子是STN 和SOC(图4d)。

图4 土壤理化性质和植物群落特征指标主成分分析Fig.4 Principal component analysis of soil physicochemical properties and plant community characteristicsBGB,地下生物量;AGB,地上生物量;SM,土壤含水量;SBD,土壤容重;SAN,土壤速效氮含量;STN,土壤全氮含量;SAP,土壤速效磷含量;STP,土壤全磷含量;SOC,土壤有机碳含量;DOC,土壤可溶性有机碳含量。a,c 分别为高寒草甸和高寒草原群落多样性指标主成分分析;b,d 分别为高寒草甸和高寒草原土壤理化性质主成分分析。BGB,belowground biomass;AGB,aboveground biomass;SM,soil moisture;SBD,soil bulk density;SAN,soil available nitrogen;STN,soil total nitrogen;SAP,soil available phosphorus;STP,soil total phosphorus;SOC,soil organic carbon;DOC,dissolved organic carbon. a,c indicate principal component analysis of community diversity indicators in alpine meadow and alpine steppe;b,d indicate principal component analysis of soil physical and chemical properties in alpine meadow and alpine steppe.
通过对2 种草地类型土壤微生物量碳氮含量构建结构方程模型,发现STN 对高寒草甸MBC 和MBN 含量均有显著正效应(P<0.05),效应系数分别为0.40 和0.42(图5a),而SOC、Simpson 指数对高寒草原MBN 含量均有显著正效应,且SOC 对MBC 含量也具有显著正效应(P<0.05),效应系数分别为0.32、0.31 和0.25(图5b)。GSP 对高寒草甸MBC 和MBN 含量均有显著正效应(P<0.05),效应系数分别为0.51 和0.47,对高寒草原MBC和MBN 含量也具有显著正效应,效应系数分别为0.43 和0.24(P<0.05)。

图5 气候、植物群落和土壤关键因子对不同草地类型土壤微生物量碳(MBC)和氮(MBN)的效应Fig. 5 Effects of climate,plant community and key factors on soil microbial biomass carbon(MBC)and nitrogen(MBN)in different grassland types图上表示的均为显著影响的路径(P<0.05),绿色实线表示正效应。GSP,生长季降水量;a,高寒草甸;b,高寒草原。All the paths shown in the figure had significant effects(P<0.05).The solid green line represents a positive effect.GSP,growing season precipitation;a,alpine meadow;b,alpine steppe.
3 讨论
3.1 不同草地类型土壤微生物量碳氮含量的差异
土壤微生物量碳氮含量的多少是表征土壤碳氮循环的重要指标,直接影响到土壤养分含量以及供给植物养分的能力[35−36]。在不同的地区,由于气候因子、植被特征、土壤理化性质以及土壤养分含量的不同,这使得土壤微生物生物量碳氮含量有所差异[37−39]。高寒草甸和高寒草原由于物理和生物学性质的差异,导致两种不同的草地类型土壤微生物量碳氮含量存在差异。在本研究中,青藏高原高寒草甸MBC 和MBN 含量均显著高于高寒草原(P<0.01),且两种草地类型数据分布特点不同(图1)。其原因在于两种草地类型的主导限制因子不同,导致MBC 和MBN 含量有所差异。高寒草甸MBC 和MBN 含量高于高寒草原可能是因为高寒草甸比高寒草原具有更充沛的降水量,且物种丰富度和生物量较高,从而导致高寒草甸土壤有机质积累较多,而高寒草原土壤有机质相对较少,而有机质的含量与土壤微生物量密切相关[28]。例如,在祁连山地区,MBC 含量高寒草原比高寒草甸中高40%以上,0~5 cm 土层的MBN 含量,高寒草甸比高寒草原高29%[40]。本研究结果中两种草地类型MBN 含量的差异与其相似,而MBC 含量与其相反,究其原因,可能是本研究包括了青藏高原大部分高寒草地MBC 和MBN 含量的测定,而在祁连山地区开展的研究是在相对较小的尺度上开展[37],局部地区可能有不同的小生境以及小气候所致。也有相关研究表明[41−42],高寒草甸和高寒草原土壤微生物量碳氮含量均达显著差异,这是由于高寒草甸主要以嵩草属和薹草属为主要建群种,而高寒草原则以禾本科和杂类草为主,高寒草甸地上生物量高于高寒草原,并且嵩草属植物根系要比禾本科植物发达,从而使得植物地下部分比较发达,根际微生物量含量较高,这是高寒草甸土壤微生物量碳氮含量高于高寒草原的主要原因。除此之外,与高寒草甸相比,高寒草原土壤含水量低,适宜的土壤水分含量会增加土壤微生物量,大多数微生物在低湿条件下由于干旱胁迫而分解死亡,因此导致土壤微生物量下降。
3.2 土壤微生物量碳氮含量对水热条件的响应
目前,已经有诸多研究表明降水或水分对土壤微生物量碳氮含量有很大的影响[43−44]。相关分析结果表明,本研究区两种不同草地类型土壤MBC 和MBN 含量受生长季降水量(GSP)影响显著(P<0.01)(图3 和表1)。这是由于青藏高原整体海拔较高,年平均温度较低,降水量的不同形成了高寒草甸和高寒草原两种不同的草地类型[27]。随着降水量的增加能够促进生态系统对土壤有效氮和碳含量的积累,从而有利于土壤养分的矿化和积累[45],土壤中有效养分含量的增加使得土壤微生物可以积累更多的微生物量碳氮含量,在本研究中土壤微生物生物量碳氮含量随着降水的增加而呈现上升的趋势。因此,生长季降水是控制两种草地类型MBC 和MBN 含量的关键气候因子,这与前人的研究结果一致[19,46]。在本研究中,高寒草原生长季普遍降水量低,因此土壤微生物量碳氮含量受降水限制性较强,使得青藏高原不同区域高寒草原MBC 和MBN 含量差异不大,而高寒草甸区域跨度较大,降水条件不均匀,导致土壤微生物量碳氮含量差异较大,数据分布相对离散(图1)。
在不同研究区域,温度对土壤微生物量的影响有所差异,相关分析结果表明,本研究区仅高寒草原MBN 含量与生长季均温(GST)呈显著负相关(图3 和表1)(P<0.01)。年均温可以通过改变凋落物的质量和数量从而使分解者食物网的结构和功能受到影响,温度升高引起的植物群落结构的改变,进而影响土壤生物的养分组成,最终对土壤微生物产生影响。高寒草原降水条件有限,土壤含水量较低,植物群落多样性低,温度升高使得土壤的干旱程度增加,使土壤微生物活动受到限制,因此,土壤微生物量随着生长季均温的升高而减少。由此可见,降水是控制该地区土壤微生物量的主要因素,而并非温度。
3.3 土壤微生物量碳氮含量对群落和土壤因子的响应差异
土壤微生物量与地上植被以及土壤因子的关系十分密切[47],不同的草地类型由于地上植被和土壤营养含量的差异,导致土壤微生物量也有一定的差异。结构方程模型分析发现,土壤全氮(STN)对高寒草甸MBC 和MBN 含量具有显著正效应(P<0.05),而高寒草原MBC 和MBN 含量受土壤有机碳(SOC)影响显著(P<0.05)(图5)。植被类型不同将导致土壤全氮和有机碳的含量不同[48],与高寒草甸相比,高寒草原植被盖度降低,植物种类减少,植物枯落物少,从而导致地表裸露面积大于高寒草甸,且土壤保持水分的能力降低,SOC 的含量也随之下降。SOC 含量下降主要是因为高寒草原草地植被发生变化,导致返还土壤的凋落物的数量减少,使得植物群落中碳素的输出大于输入,显著影响了土壤有机碳的积累[49]。因此,高寒草原SOC 含量普遍较低,导致土壤微生物量碳氮含量差异不大。与高寒草甸相比,高寒草原为土壤微生物提供的凋落物较少,从而影响到土壤微生物能源物质来源及数量,导致土壤微生物对SOC 变化极为敏感,SOC 是调控MBC 和MBN 含量的主要因素,这与前人的研究结果一致[50]。氮素是限制草地生产力的一个重要因素,速效养分的多少决定了土壤氮素的有效性,虽然高寒草甸土壤氮素储量丰富,但有效性很低,因此影响其生态系统生产力的因素一直被认为是受到氮素的限制[51],但是在不同区域的高寒草甸,可能由于降水等因子的控制,土壤氮素含量不同,从而导致MBC 和MBN 含量差异较大。因此,高寒草甸受氮素影响的敏感度要大于高寒草原,使得高寒草甸土壤微生物生物量碳氮含量主要受土壤全氮的调控[52]。
4 结论
综合研究5 种植物群落因子、8 种土壤因子以及气候因子的关系,揭示了生物因子和非生物因子对青藏高原高寒草甸和高寒草原两种草地类型MBC 和MBN 含量的重要影响。研究发现,不同草地类型间的MBC 和MBN含量差异显著,表现为高寒草甸MBC 和MBN 含量高于高寒草原,并且土壤全氮(STN)是高寒草甸土壤微生物碳氮含量变化的主要调控因子,而高寒草原土壤微生物量碳氮含量则主要受土壤有机碳(SOC)的调控。本研究可为青藏高原区域尺度土壤微生物量研究提供一定的参考。
