解淀粉芽胞杆菌不同整合位点对外源碱性蛋白酶表达的影响
2022-06-09牛馨张莹王茂军刘文龙路福平李玉
牛馨 张莹 王茂军 刘文龙 路福平 李玉
(1.工业发酵微生物教育部重点实验室 天津科技大学生物工程学院,天津 300457;2.山东隆科特酶制剂有限公司,沂水 276400)
蛋白酶被广泛的应用于食品[1]、洗涤[2]、医药[3]以及皮革[4]等行业,其中碱性蛋白酶所占比例达到40%[5],并且市场对碱性蛋白酶的需求量还在逐年增加,但目前产量依然无法满足工业上的需求[6]。在利用微生物发酵生产碱性蛋白酶的过程中,存在生产菌株性能不稳定、酶的表达量无法控制等现象,因此寻求高效稳定的表达策略,对碱性蛋白酶实现工业化生产具有重大意义。
为了解决生产菌株的遗传稳定性问题,可将外源基因整合到表达菌株基因组上以达到稳定遗传的目的,而整合基因的表达水平往往取决于它们在基因组上的整合位置[7]。Bryant等[8]通过将 Plac-gfp表达盒整合至大肠杆菌K-12基因组的不同位点,表明整合位点在染色体上的位置会影响基因的表达水平。DNA复制起始于基因组上的固定位置(复制起点Ori)[9],基因剂量随着与Ori的接近程度增加而增加[10],因此选择在Ori附近整合外源蛋白基因是提高外源蛋白表达量的有效手段。Zhou等[11]在Ori附近整合了aprE基因,提高了碱性蛋白酶的酶活。
另外,在细菌中大部分的胞外酶都通过Sec途径分泌至胞外[12],异源蛋白通过相同的分泌机制输出到胞外可能会引起细胞膜的堵塞[13],因此在细菌基因组胞外酶位点整合外源蛋白基因可以减少菌体自身分泌的胞外酶对资源的利用,有利于外源蛋白的表达。Ren等[14]在枯草芽胞杆菌基因组胞外蛋白酶位点(epr和bpr)以及淀粉酶位点amyE整合了外源基因Bgal1-3使整合菌株的胞外酶活比原始菌株提高了87%,并且整合菌株在150代以后依然能够稳定遗传。Watzlawick等[15]将ganA基因整合到枯草芽胞杆菌基因组amyE和nprB等5个位点,使β-半乳糖苷酶活性达到8.36 U/mg,这些研究表明在胞外蛋白酶位点以及胞外淀粉酶位点整合外源蛋白是有效提高外源蛋白表达量的一种方法。
本研究以实验室已有的生产菌株解淀粉芽胞杆菌Bacillus amyloliquefaciens TCCC 111018为研究对象,结合其基因组测序结果和数据分析,选择在OriC附近、胞外蛋白酶以及胞外淀粉酶位点进行外源碱性蛋白酶基因(aprE)的整合,以此探究基因组上不同位点对外源碱性蛋白酶表达的影响,为利用基因组整合方式构建表达不同外源蛋白的生产菌株奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 研究使用的菌株和质量见表1。

表1 研究中使用的菌株和质粒Table 1 Bacterial strains and plasmids used in this study
1.1.2 酶与试剂 DNA聚合酶、限制性核酸内切酶购自TaKaRa公司;无缝克隆和反转录试剂盒购自北京全式金公司;RNA提取试剂盒购自天根公司;质粒提取试剂盒、DNA 切胶回收试剂盒购自Omega公司;福林酚、酪蛋白底物、三氯乙酸等购自上海生工公司。
1.1.3 培养基 种子培养基(LB培养基):1% 蛋白胨,1% NaCl,0.5% 酵母提取物;发酵培养基:6.4%玉米粉,4% 豆粕粉,0.4% 磷酸氢二钠,0.03% 磷酸二氢钾,0.07% 高温淀粉酶。
1.2 方法
1.2.1 胞外蛋白酶缺失菌株以及整合菌株的构建 通过基因组序列分析和比对,确定了TCCC 111018基因组上至少有6个胞外蛋白酶:Mpr、Vpr、NprE、Epr、Apr和Bpr,采用同源重组的方法使6个胞外蛋白酶基因缺失,缺失过程以vpr基因为例。使用表2所示的引物扩增相应的目的片段,采用无缝克隆方式构建敲除质粒pWH-T2-vpr,将pWH-T2-vpr化转至 EC135 pM.Bam[16];再将经过甲基化修饰的质粒电转至解淀粉芽胞杆菌,筛选正确的转化子进行单、双交换,共构建了6株蛋白酶缺失菌株,并将pLY-2质粒转化至缺失菌株中。

表2 vpr基因缺失的引物设计Table 2 Primers for vpr gene deletion
基因整合的方法与敲除方法一样,在nprE、yaah以及amyE位点整合了aprE基因,与敲除方法不同的是在左右同源臂之间加入了一段整合基因片段,构建示意图如图1所示。

图1 基因整合构建示意图Fig.1 Schematic diagram of gene integration construction
1.2.2 胞外蛋白酶缺失菌株生长曲线的测定 将不同菌株在LB培养基中摇瓶发酵32 h,每隔2 h取样并使用酶标仪在600 nm的波长下测其吸光度值,以时间为横坐标,以OD值为纵坐标绘制生长曲线。
1.2.3 碱性蛋白酶表达量以及酶活的测定 将6株带有pLY-2质粒的蛋白酶缺失菌株以及3株整合菌株在发酵培养基中摇瓶发酵48 h,取发酵48 h后的上清液采用福林法(GB/T23527-2009)测定其酶活,并对TCCC 111018发酵上清液进行SDS-PAGE检测。
1.2.4 整合菌株转录水平的测定 提取3株整合菌株的RNA进行实时荧光定量PCR检测,使用引物如表3所示。

表3 用于实时荧光定量PCR的引物Table 3 Primers for real-time fluorescent quantitative PCR
1.2.5 数据分析 本研究所有实验均重复至少3次,并且每次设置3个平行,采用Origin8.0进行数据处理和分析。
2 结果
2.1 yaah整合位点的选择
基因的表达受基因与复制起始点OriC距离的影响,越靠近复制起始点基因的表达水平越高,因此通过Ori-Finder(http://tubic.tju.edu.cn/doric/)预测了TCCC 111018基因组的复制起始位点OriC,结果表明TCCC 111018的OriC总长度为1 021 bp,起始于基因组上的1 904 168 bp,终止于1 905 188 bp。比较距离OriC位点较近的几个基因中,Yaah是芽胞萌发的相关蛋白[17],芽胞可以帮助细菌度过恶劣的环境,而解淀粉芽胞杆菌不产生芽胞,所以该基因的缺失不会影响菌体的生长,因此可以在该位点整合aprE基因。
2.2 nprE整合位点的选择
2.2.1 胞外蛋白酶缺失菌株的构建 为确定更多的整合位点,首先探究了原始菌株B.amyloliquefaciens TCCC 111018基因组上胞外蛋白酶对外源碱性蛋白酶aprE基因表达的影响。利用同源重组技术分别缺失了6个胞外蛋白酶基因(epr、vpr、mpr、apr、bpr和nprE),并将6株缺失菌株分别命名为:18-ΔE、18-ΔV、18-ΔM、18-ΔA、18-ΔB 和 18-ΔN,各敲除位点在基因组上的位置示意图如图2所示。

图2 敲除位点在TCCC 111018基因组的位置图Fig.2 Diagram showing the location of the knockout sites in the TCCC 111018 genome
2.2.2 胞外蛋白酶缺失菌株生长曲线的测定 为了探究胞外蛋白酶的缺失对菌体生长是否有影响,对6株缺失菌株进行了生长曲线的测定,结果如图3所示。18-ΔV在14 h之前生长状况与原始菌株18基本一致,但14 h之后生长情况却不如原始菌株,说明vpr基因的缺失可能会导致菌体的自溶速度加快[18];其它5株缺失菌株与原始菌株的生长状况没有明显的差别,说明mpr、nprE、epr、apr和bpr基因的缺失对菌体的生长没有显著影响。

图3 胞外蛋白酶缺失菌株的生长曲线图Fig.3 Growth curve of extracellular protease-deficient strain
2.2.3 胞外蛋白酶缺失菌株对外源碱性蛋白酶的影响 为了探究胞外蛋白酶的缺失对外源碱性蛋白酶表达的情况,将质粒pLY-2转化至原始菌株B.amyloliquefaciens TCCC 111018及6株蛋白酶缺失菌株,分别将其命名为:CK、ΔE、ΔV、ΔM、ΔA、ΔB和ΔN,经发酵后测定了碱性蛋白酶的酶活,如图4所示。原始菌株CK的酶活为11 937.08 U/mL,缺失菌株ΔN的酶活为13 231.93 U/mL,比原始菌株提高了7.5%;ΔV酶活最低,比原始菌株降低了约15%;而mpr、epr、apr和bpr基因缺失菌株酶活与原始菌株无明显差异,表明nprE基因的缺失对外源碱性蛋白酶的表达有促进作用,因此选择在该位点整合aprE基因。
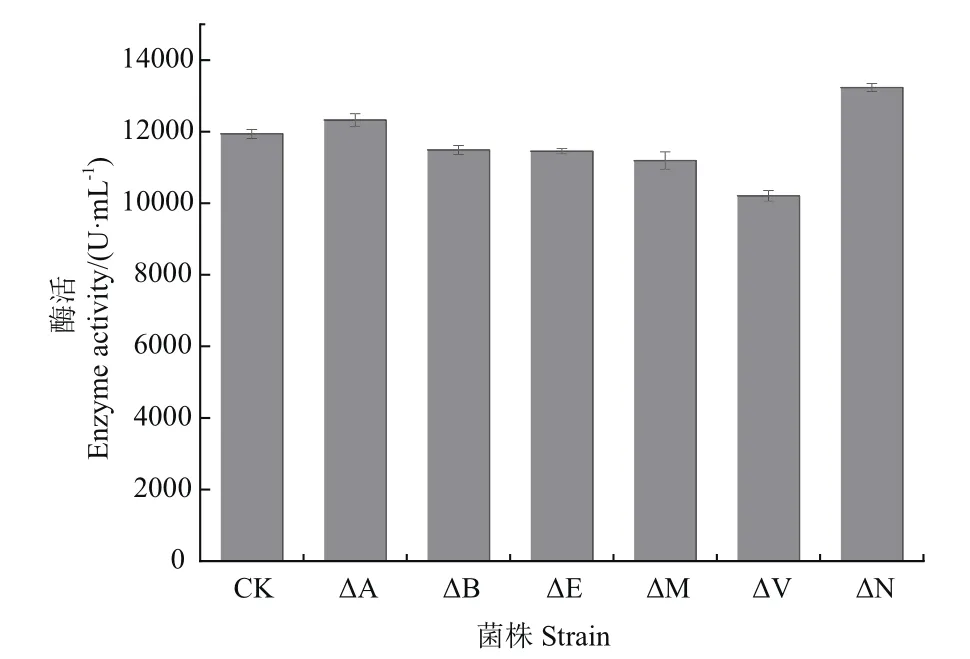
图4 胞外蛋白酶缺失菌株酶活力Fig.4 Enzyme activity of extracellular protease-deficient strains
2.3 amyE整合位点的选择
对原始菌株B.amyloliquefaciens TCCC 111018的发酵上清液进行了SDS-PAGE检测,结果如图5所示,在分子量为45.0-66.2 kD之间的蛋白条带更明显,表明该蛋白的表达量相对较高。

图5 TCCC 111018发酵液上清SDS-PAGE检测Fig.5 SDS-PAGE detection of TCCC 111018 fermentation broth supernatant
随后将图5中表达量最高的条带回收并进行质谱鉴定,质谱分析结果如表4所示,表明该蛋白为α-淀粉酶,说明在原始菌株TCCC 111018的发酵液上清中α-淀粉酶的表达量最高。

表4 质谱结果分析Table 4 Analysis of mass spectrometry results
由于α-淀粉酶与碱性蛋白酶均为胞外酶,分泌途径可能相同,因此针对二者的氨基酸序列利用 SignalP-5.0 Server(http://www.cbs.dtu.dk/services/SignalP/)软件进行信号肽预测分析。结果表明二者的信号肽均为Sec途径,由此可以推断两个胞外酶可能均由Sec途径分泌至胞外,Sec转运系统包括信号肽识别颗粒、分子伴侣、信号肽酶及通道蛋白SecYEG等蛋白分子[19],而α-淀粉酶的高表达量可能会占据这些资源,推测在该位点整合aprE基因有利于碱性蛋白酶的表达。
2.4 构建表达外源碱性蛋白酶的整合菌株
为了获得稳定的表达外源碱性蛋白酶菌株,根据以上研究结果,选择了在yaah、nprE以及amyE基因位点进行基因aprE的整合,构建的3株整合菌 株 分 别 命 名 为 18-ΔY∷aprE、18-ΔN∷aprE 和18-Δα∷aprE。
2.4.1 整合菌株生长曲线以及酶活的测定 为探究不同整合位点的aprE对菌株生长的影响以及碱性蛋白酶的酶活水平,对3株整合菌株进行了生长曲线以及酶活的测定,结果如图6、图7所示。18-ΔN∷aprE在12 h前比原始菌株生长速率快,但在16 h后生长却明显慢于原始菌株,出现这种现象的原因可能是18-ΔN∷aprE的酶活显著提高(图7)。由于碱性蛋白酶可以水解培养基中的蛋白物质为菌体的生长提供营养,该酶的高水平表达利于菌体的快速生长,而外源碱性蛋白酶的高表达会给菌株造成生长的负担,因此在生长后期菌体生长状况不如原始菌株。18-ΔY∷aprE在6-10 h 生长速率比原始菌株快,但在10 h后与原始菌株生长速率基本一致,原因可能是18-ΔY∷aprE比原始菌株的酶活提高了约15%利于菌体的前期生长。18-Δα∷aprE与原始菌株的生长状况基本没有差别,图7也表明18-Δα∷aprE酶活水平没有原始菌株高,因此对菌株生长无显著影响。

图6 整合菌株的生长曲线图Fig.6 Growth curve of integrated strains
综合上述分析,结合图7的结果,18-ΔN∷aprE比原始菌株的酶活提高了近89%,说明在nprE位点整合aprE基因可以显著提高碱性蛋白酶的酶活力;而18-Δα∷aprE酶活水平却明显低于了原始菌株,原因可能是将外源基因整合到该位点的同时也破坏了该基因,说明α-淀粉酶基因的缺失不利于外源碱性蛋白酶的表达,可能是由于α-淀粉酶基因缺失后菌体对培养基中碳源的利用有影响。

图7 整合菌株的酶活力分析Fig.7 Analysis of enzyme activity of integrated strains
2.4.2 整合菌株转录水平及碱性蛋白酶表达情况的分析 为探究不同整合位点的aprE的转录水平以及碱性蛋白酶的表达情况,对3株整合菌株进行了转录水平的测定以及SDS-PAGE凝胶电泳的检测,结果如图8和图9所示。由图8可知,18-ΔN∷aprE的相对转录水平最高;图9也显示出18-ΔN∷aprE的碱性蛋白酶表达量最高,因此在nprE位点整合amyE能够有效提高碱性蛋白酶的表达量可能是提高了碱性蛋白酶的转录水平,推测nprE位点的启动子转录水平较强;图8显示出18-Δα∷aprE和18-ΔY∷aprE在转录水平上两株菌相差不大,且显著低于18-ΔN∷aprE。由图9可知这两株菌的碱性蛋白酶表达量均与对照18无明显差别,且均没有18-ΔN∷aprE的碱性蛋白酶表达量高,表明yaah和amyE位点可能没有nprE位点的启动子转录水平高。

图8 整合菌株aprE的转录水平Fig.8 Transcription level of aprE in the integrated strains

图9 整合菌株AmyE的SDS-PAGE检测Fig.9 SDS-PAGE detection of AmyE in the integrated strains
3 讨论
本研究采用基因组整合的策略使碱性蛋白酶表达量比原始菌株提高了89%,Zhang等[20]也通过将Mglu基因整合到枯草芽胞杆菌基因组上使谷氨酰胺酶在5 L发酵罐中最大酶活达到357.6 U/mL;Mo等[21]将碱性蛋白酶apr4基因整合到嗜碱芽胞杆菌的基因组上使碱性蛋白酶的活力提高了111.3%。这些研究表明了基因组整合是有效提高外源蛋白表达的一种方法。
本研究在确定nprE基因整合位点的研究中发现中性蛋白酶nprE基因的缺失较其他蛋白酶基因能够提高外源碱性蛋白酶的酶活力。Wang等[22]也通过缺失解淀粉芽胞杆菌基因组上的中性蛋白酶Banpr基因显著提升了异源蛋白的产量,这与我们得到的结果一致。Kawamura等[23]通过敲除枯草芽胞杆菌基因组中的中性蛋白酶基因(nprE)和碱性蛋白酶基因(aprE),发现敲除菌株的胞外蛋白酶酶活比原始菌株减少了96%以上,说明枯草芽胞杆菌分泌的胞外蛋白酶主要是碱性蛋白酶和中性蛋白酶[24-25],因此,缺失中性蛋白酶基因可能有利于宿主菌中更多的资源用于碱性蛋白酶的表达及分泌。同时,细菌自身分泌的胞外蛋白酶能水解胞外的蛋白从而为菌体的生长提供营养[26],所以胞外蛋白酶会降解外源蛋白而影响目标蛋白的产量。在本研究中,可能是由于中性蛋白酶缺失菌株减少了对外源碱性蛋白酶的降解量从而提高了碱性蛋白酶的表达量。虽然其它蛋白酶都有降解作用,但由于在枯草芽胞杆菌中分泌蛋白主要是中性蛋白酶和碱性蛋白酶,而解淀粉芽胞杆菌与枯草芽胞杆菌的亲缘性极高[22],所以我们在解淀粉芽胞杆菌中缺失中性蛋白酶基因能显著减少碱性蛋白酶的损失而使碱性蛋白酶的表达量提高。
本研究在原始菌株发酵上清液中表达量最高的α-淀粉酶位点整合了外源碱性蛋白酶基因,结果发现整合菌株的酶活力没有原始菌株高。研究表明,淀粉酶是一类能够水解淀粉分子内的糖苷键从而得到葡萄糖或糊精等产物的一类酶[27]。由于发酵培养基中碳源的主要成分为淀粉,当淀粉酶基因缺失后对培养基中碳源的利用率降低[28],可能是这些因素导致了在淀粉酶位点整合碱性蛋白酶基因后,碱性蛋白酶的表达量并没有提升反而有所下降。
为获得表达外源蛋白的高产菌株,许多研究开始倾向于精简基因组,删除表达菌株中的冗余基因,构建底盘微生物。Reuβ等[29]通过删除了B.subtilis基因组36%的基因,使细胞内的资源利用更为合理,为构建高效表达外源蛋白的宿主菌有着重要的意义。本研究通过删除6个胞外蛋白酶发现对菌体的生长并无显著影响,并且发现删除了中性蛋白酶基因后使外源碱性蛋白酶的表达量有所提升,为构建高效表达外源蛋白的底盘微生物提供了一定的参考。
4 结论
本研究通过在Bacillus amyloliquefaciens TCCC 111018基因组上不同位点整合了外源碱性蛋白酶基因aprE,获得了一株高效表达碱性蛋白酶的菌株18-ΔN∷aprE,其相对酶活提高了近89%,提升了TCCC 111018作为工业菌株的价值,并为构建稳定表达外源蛋白的工程菌提供了一种思路方法。
