密度和滴灌量对高产春玉米叶片生理指标的影响
2022-06-09陈江鲁王群蒋兴星张小伟
陈江鲁 王群 蒋兴星 张小伟
摘要:以先玉335为研究对象,探究不同种植密度下滴灌量对春玉米叶片生理指标的影响,为新疆高产春玉米节水灌溉提供理论和试验依据。采用2因素裂区试验设计,主处理密度(D)共4个水平,即D1(4.5万株/hm2)、D2(7.5万株/hm2)、D3(10.5万株/hm2)、D4(13.5万株/hm2),以滴灌量(I)为副区,设置3个不同灌量(6 000、4 800、3 600 m3/hm2),在玉米生长的主要生育时期,测定叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性,以及丙二醛(MDA)、脯氨酸(Pro)、可溶性蛋白(SP)和可溶性糖(SS)含量。结果发现,滴灌量降低会使保护性酶活性略有升高或变化不显著;但灌浆后部分密度受水分亏缺时,酶活性降低。MDA、Pro含量在前期均不随滴灌量降低有显著差异;灌浆初期D3密度下,MDA含量随滴灌量减少有升高趋势;D4密度和乳熟期的MDA含量均随滴灌量减少有降低趋势。SS既是分解的产物,也是最初的同化产物;SP含量随滴灌量减少有降低趋势或差异不显著。总体来看,即使在高密度群体内,先玉335的叶片对轻中度水分胁迫仍有一定弹性,即灌水可以缓解植株群体密度增加的压力,以保证玉米叶片正常的生理特性,减轻水分胁迫对细胞代谢过程的伤害。
关键词:高产春玉米;种植密度;滴灌量;保护性酶;渗透调节物质
中图分类号:S513.04;S513.07 文献标志码: A
文章编号:1002-1302(2022)09-0055-07
玉米在粮食作物栽培过程中需水量大,干旱胁迫严重影响我国玉米生产。干旱胁迫会导致农作物体内的活性氧产生与清除机制失衡,从而造成作物器官活性氧累积增加,对农作物蛋白质、膜脂等其他细胞成分构成伤害[1]。Bacelar等研究发现,灌溉可以提高橄榄树叶片抗氧化能力,降低氧化损伤[2]。耐密型玉米具有较高的清除活性氧的能力和抗逆性,能够促进干物质积累,有利于延缓其衰老,是获得高产的重要生理基础[3]。研究表明,干旱条件下作物叶片内在保护酶系统活性被诱导,减轻了膜脂过氧化程度[4-6],植物内源保护酶活性的变化会因植物材料、不同胁迫方式与胁迫程度而异[1]。干旱可使玉米叶片及根系的有机渗透调节物质上升[7],且干旱程度越严重,其有机渗透调节物质上升幅度越大,明显改变了作物的渗透压,使土壤水难以运输到作物体内,降低其蒸腾速率[8]。因此,协调灌溉量和密度的促控功效,对塑造高效作物个体和合理群体具有重要意义[9]。前人关于干旱胁迫、种植密度对玉米生理指标影响的研究较多,但多集中在低密度、较大水分胁迫程度下,本研究在前人的基础上研究提高种植密度、降低水分胁迫程度,以及水密互作下保护性酶活性和渗透调节物质含量在不同处理下的差异及变化规律,进一步探究不同密度下滴灌量对玉米功能叶生长发育的促控作用,为春玉米高产节水栽培研究提供理论依据。
1 材料与方法
1.1 试验地点和试验设计
试验于2020年在新疆维吾尔自治区奇台农场(89°46′01″E,43°50′41″N)高产试验田进行,海拔750 m,太阳总辐射量≥ 546.3 kJ/cm2,年大于10 ℃积温3 000~3 200 ℃,年日照时数2 500~2 800 h,无霜期130~156 d,年均降水量180~480 mm。试验于2016年开展,选用先玉335玉米杂交种为试验材料,采用裂区设计,密度為主区,滴灌量为副区,密度设置为D1(4.5万株/hm2)、D2(7.5万株/hm2)、D3(10.5万株/hm2)、D4(13.5万株/hm2),滴灌量水平为I1(6 000m3/hm2)、I2(4 800 m3/hm2)、I3(3 600 m3/hm2),3次重复,小区面积60 m2,宽窄行(70 cm+40 cm)种植。4月 18日播种,采用膜下滴灌,10 月 13日收获。V10期设置不同的灌溉水平,灌溉量用水表计量控制,灌溉间隔为10 d。其他管理与大田生产相同。
1.2 测定项目及方法
在玉米大喇叭口期,于各处理选取具有代表性的玉米,取下最后1张展开叶;吐丝期、灌浆初期、乳熟期,取穗位叶,置于超低温冰箱中保存,用于生理指标测定。氮蓝四唑法测定超氧化物歧化酶(SOD)活性、考马斯亮蓝G-250法测定可溶性蛋白(SP)含量、硫代巴比妥酸法测定丙二醛(MDA)含量,以上方法均参照李合生的 《植物生理生化实验原理和技术》[10];紫外分光光度法[11]测定过氧化氢酶(CAT)活性的相对大小(K值);愈创木酚法测定过氧化物酶(POD)活性、蒽酮比色法测定可溶性糖(SS)含量,参照高俊凤的《植物生理学实验指导》[12];脯氨酸(Pro)含量测定参照张宪政的《作物生理研究法》[13]。
1.3 数据处理
数据通过DPS 7.05软件进行SSR检验和Duncans多重比较法(α=0.05)进行统计分析,图表使用Microsoft Excel 2010制作。
2 结果与分析
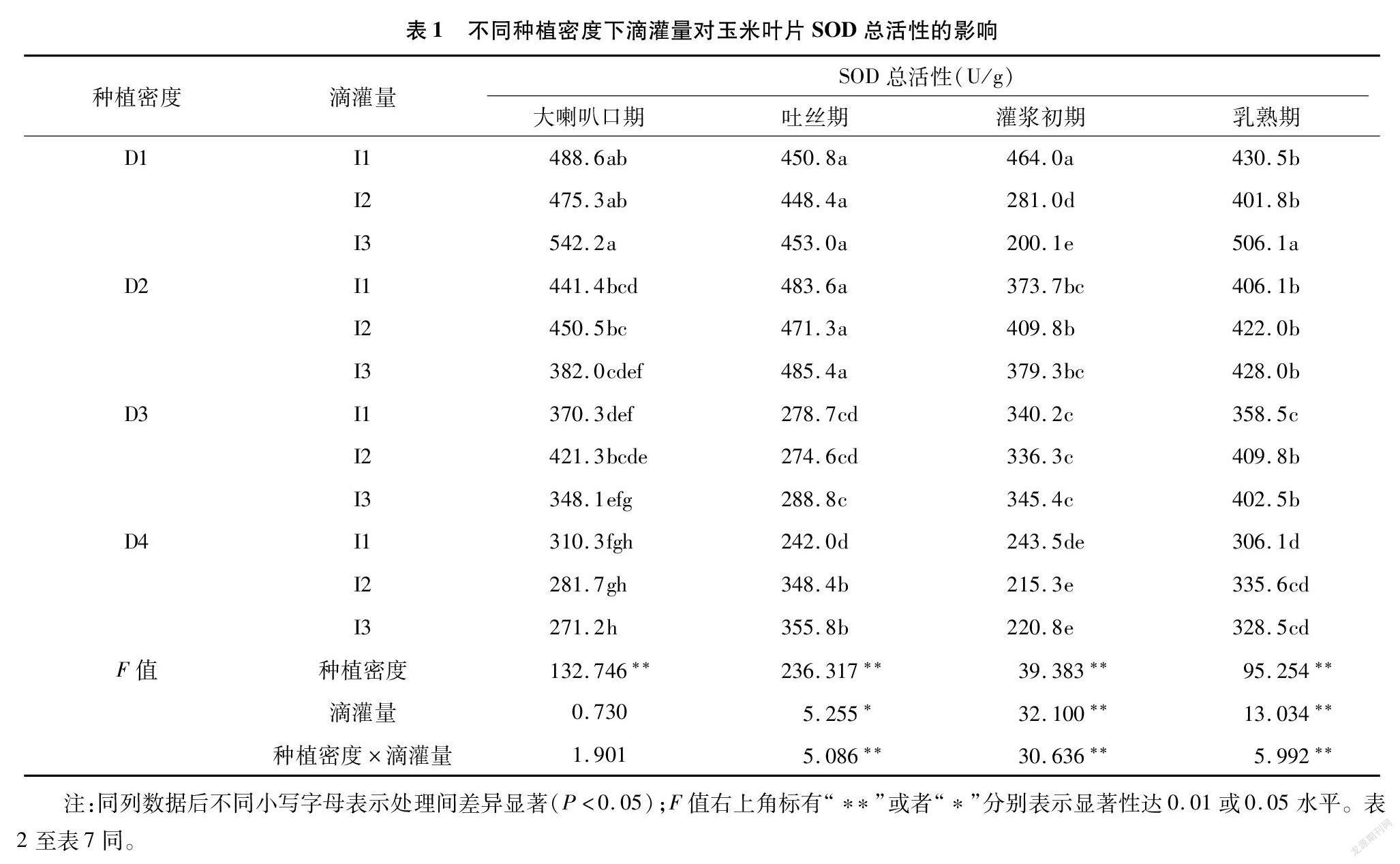
2.1 不同种植密度下滴灌量对玉米叶片超氧化物歧化酶(SOD)活性的影响
从表1可以看出,各生育时期内不同种植密度间的SOD活性均达到极显著水平,除大喇叭口期外,其他生育时期的SOD活性在不同滴灌量间均达到显著或极显著水平,且随种植密度增大SOD活性总体呈降低趋势。在吐丝期的D4密度下,SOD活性随滴灌量减少有升高趋势,I2、I3处理间差异不显著。进入灌浆初期,除D1处理下SOD活性随滴灌量的减少而降低外,其他密度下酶活性均不随滴灌量下降有显著变化。D2密度下乳熟期的SOD活性随滴灌量减少有增加趋势,且D2、D4下各灌量处理间差异不显著。
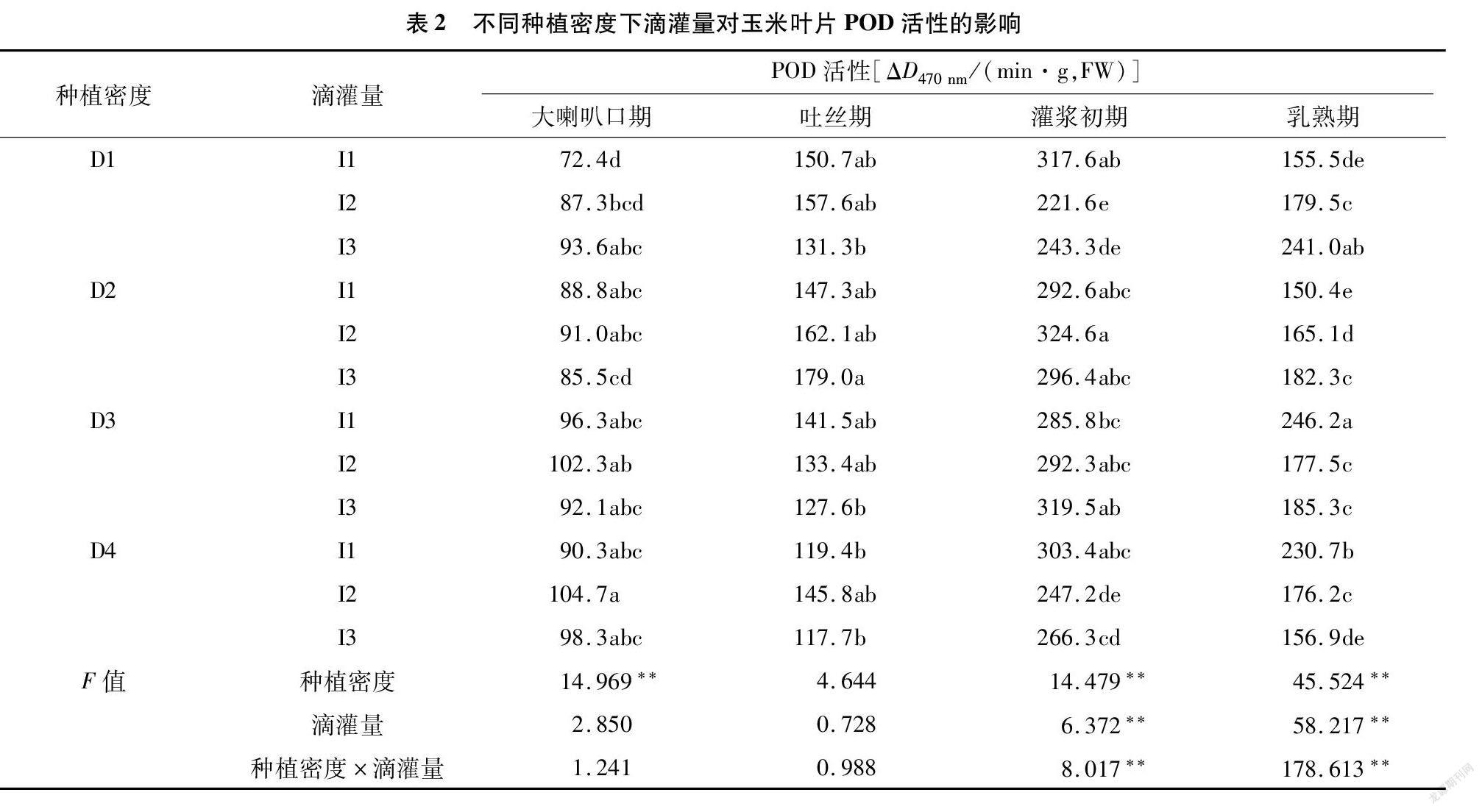
2.2 不同种植密度下滴灌量对玉米叶片过氧化物酶(POD)活性的影响5A43E19C-6A53-4052-BC07-6D590CC13758
从表2可以看出,除吐丝期外不同密度间的POD活性均达到极显著水平; 灌浆初期和乳熟期的POD活性在不同滴灌量间均达到极显著水平。在大喇叭口期和吐丝期的各密度下,POD活性均不随滴灌量减少有显著变化。进入灌浆初期,表现为D1密度下I2和I3的POD活性显著低于I1;D4密度下I2的POD活性显著低于I1,I3和I1差异不显著;D2、D3处理下差异不显著。乳熟期的D1、D2密度处理下,随滴灌量减少POD活性有升高趋势;D3密度下I1与I2和I3差异显著,随着滴灌量的降低,POD活性呈先降低后升高趋势;D4密度下POD活性随滴灌量降低呈下降趋势,I1、I2、I3之间差异显著。
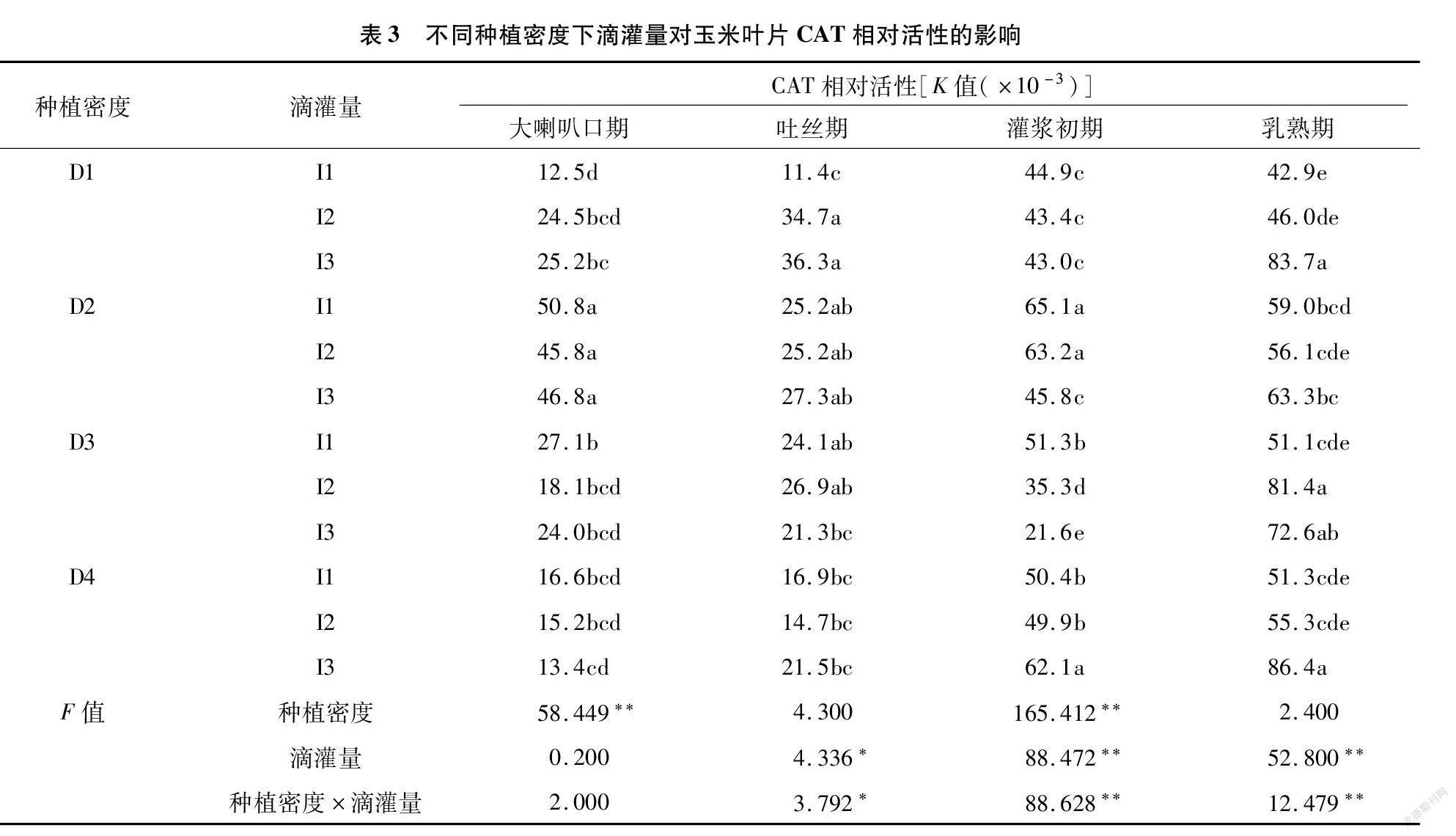
2.3 不同种植密度下滴灌量对玉米叶片过氧化氢酶(CAT)活性的影响
从表3可以看出,在大喇叭口期和灌浆初期的不同密度间CAT活性有极显著差异;除大喇叭口期外,不同滴灌量间的差异均达到显著或极显著水平。在吐丝期的D1密度下,I1处理的CAT活性显著低于I2、I3。灌浆初期,D1~D3密度下,随滴灌量减少CAT活性有降低趋势,密度越大,不同灌量间差异越显著;而D4密度下,酶活性则有先降后升趋势,且I1、I2处理间差异不显著。进入乳熟期时,D1和D4密度下,随滴灌量减少CAT活性有升高趋势;D2密度下,各灌量处理的CAT活性差异不显著;D3密度下,随滴灌量减少CAT活性呈先升后降趋势。
2.4 不同种植密度下滴灌量对玉米叶片丙二醛(MDA)含量的影响
从表4可以看出,MDA含量在各生育时期的不同密度处理间差异均达到显著或极显著水平;在生育后期,不同滴灌量处理间的差异均达到极显著水平。灌浆初期D3密度下,I3灌量的MDA含量显著高于I1、I2处理。到乳熟期时,随滴灌量减少MDA有降低趋势。
2.5 不同种植密度下滴灌量对玉米叶片脯氨酸(Pro)含量的影响
从表5可以看出,除大喇叭口期外,不同密度条件下的Pro含量差异均达显著或极显著水平;仅灌浆初期各滴灌量处理下Pro含量有极显著差异。灌浆初期的D1~D3密度下,随滴灌量减少Pro含量升高;D4密度下,随滴灌量减少Pro呈先微降再上升趋势。
2.6 不同种植密度下滴灌量对玉米叶片可溶性糖(SS)含量的影响
从表6可以看出,在不同生育时期,不同密度下的SS含量差异均达到显著或极显著水平;灌浆初期和乳熟期的不同灌量间SS含量差异分别达到显著和极显著水平。灌浆初期的D3、D4密度下,随滴灌量减少SS含量有升高趋势。乳熟期,D1~D3处理下,随滴灌量减少SS含量有降低趋势,且I1、I2处理间差异不显著。
2.7 不同种植密度下滴灌量对玉米叶片可溶性蛋白(SP)含量的影响
从表7可以看出,除乳熟期的不同密度间差异不显著外,其他生育时期的SS含量均在不同密度间存在显著或极显著差异;仅灌浆初期SP含量在不同的滴灌量下有极显著差异。在D4密度下,随滴灌量降低SP含量有下降趋势,且大喇叭口期和灌浆初期I2、I3处理间差异不显著,吐丝期和乳熟期不同灌量间无显著差异。灌浆初期D1密度下,各滴灌量间SP含量差异不显著;D2~D4密度下,均随滴灌量减少SP含量呈下降趋势。
3 讨论与结论
3.1 不同滴灌量对玉米叶片超氧化物歧化酶(SOD)活性的影响
Tahi等指出,初期水分亏缺会增加叶片SOD活性[14]。本试验发现,灌浆初期D1密度下,SOD活性随滴灌量减少而降低; D2密度下, 随滴灌量减少呈先升后降趋势,D3、D4密度下,随滴灌量减少呈先降后升趋势。由此可以推断,在低密度条件下,灌浆初期的水分胁迫并不会促使SOD合成相关基因的表达而产生过多的SOD。
3.2 不同滴灌量对玉米叶片过氧化物酶(POD)活性的影响
本试验发现POD活性呈单峰曲线变化,峰值出现在灌浆初期,与前人研究结果[1,15]一致。在灌浆初期D4密度和乳熟期D3密度下POD活性随滴灌量减少呈先降后升趋势,即生育后期高密度条件下的水分胁迫使叶片内膜系统受损,不能产生较多的POD酶,与马秀芳等结果[16]一致;而乳熟期D1、D2密度下的POD活性却随滴灌量减少而增加,从而保证叶片的功能,这是先玉335后期叶片衰老慢,保持绿叶面积时间多,叶片功能期长的主要原因,因而具有较强的光合时间和能力,也是超高产玉米具有较高产量潜力的主要原因[17]。灌浆初期D1处理下的变化原因与SOD活性相同。
3.3 不同滴灌量对玉米叶片过氧化氢酶(CAT)活性的影响
Zhang等对匍匐翦股颖的研究发现,轻度干旱胁迫可以增加CAT活性[18]。本试验发现在D1密度下,CAT活性均随滴灌量减少有升高趋势或差异不显著,乳熟期和灌浆初期的D4密度下也发现了相似的规律;与闫素辉等对干旱胁迫下小麦旗叶CAT活性的研究结果[19]类似,可能是植株对干旱逆境的适应性反应,在一定程度上可减少自由基的积累,缓解伤害,但这种适应能力是很有限的。本试验在灌浆初期,在D1~D3密度条件下也发现土壤水分胁迫加剧会使叶片中的CAT活性降低,与石岩等研究结果[20]一致。
3.4 不同滴灌量对玉米叶片丙二醛(MDA)含量的影响
本试验发现在灌浆初期的D3密度,随滴灌量减少MDA含量有升高趋势,与前人研究结果[1,16,21]一致,本研究结果,对应处理下的SOD、POD活性差异不显著,而CAT活性有降低趋势,表明保护酶活性的下降与MDA积累密切相关,可能互为因果。一方面由于SOD、CAT活性下降,使有害自由基积累乃至超过伤害值,直接或间接启动膜脂过氧化反应,使MDA含量增加;CAT和POD是蛋白质,MDA 的聚集和增加,势必造成它们的活性下降,清除自由基的能力减弱[22]。另一方面,MDA的积累又反过来抑制保护酶活性,促使膜系统受损加重[3]。MDA含量在灌漿前并不随滴灌量减少发生显著变化,仅在乳熟期和灌浆初期的D4密度下,随滴灌量减少有降低趋势,而此时期叶片的抗氧化酶活性均保持较高水平,表明MDA含量低与耐密型品种玉米清除活性氧能力较强有关,即膜脂过氧化水平低、膜系统稳定性好、抗衰老能力强。5A43E19C-6A53-4052-BC07-6D590CC13758
3.5 不同滴灌量对玉米叶片脯氨酸(Pro)含量的影响
孙彩霞指出,渗透调节相关基因表达受环境胁迫诱导,如proβA基因编码Pro合成酶系统,其功能是促进Pro在细胞内的合成和累积;干旱条件下玉米叶片的饱和渗透势下降,Pro含量增加明显,以提高原生质的渗透压,防止水分的过多散失,对叶片正常生理代谢起到一定的保护作用[21]。本试验发现,仅在叶片代谢活动较旺盛的灌浆初期,D1~D3密度条件下,随滴灌量减少Pro含量呈增加趋势,以提高了原生质的渗透压,防止水分的过多散失;除乳熟期D1密度下I1和I2差异显著,其他时期和密度处理下滴灌量减少对Pro含量均没有显著影响。王静等指出,随水分胁迫强度增强,叶片游离Pro含量成倍增加,但不同抗旱性品种间的变化规律不明显[23];说明水分胁迫条件下叶片Pro含量不宜作为品种抗旱性鉴定指标,但可作为水分胁迫的征兆作用,不排除Pro的累积参与渗透调节、有利抗旱的作用。Wassom等通过测定不同基因型玉米叶片的游离Pro含量提出,Pro含量似乎并不是一个预测玉米对水分胁迫耐受性可靠的指标[24]。
3.6 不同滴灌量对玉米叶片可溶性糖(SS)含量的影响
在水分胁迫下一些植物可在一定的水势变化范围内通过渗透势的改变来维持膨压,如增加SS含量来抵御干旱的影响[23],从而维持细胞的正常功能[25],SS不仅参与细胞的渗透调节作用,而且是适应环境的信号物质[23]。孙彩霞发现,干旱处理时SS含量下降,并认为SS可能既是分解的产物,也是最初的同化产物[21]。王静等指出,一方面水分胁迫时光合作用受抑制,同化产物减少,致使玉米叶片SS减少;另一方面水分胁迫时有机物分解大于合成,叶片SS又有增加来抵御水分胁迫的趋势[23]。本试验也发现,在不同的处理下SS含量可能随灌溉量减少而有增加或减少的趋势。
3.7 不同滴灌量對玉米叶片可溶性蛋白(SP)含量的影响
本试验发现,水分胁迫会使玉米叶片的SP含量降低,与前人研究结果[26]一致,但此规律仅在灌浆初期和D4密度下表现明显。本试验还发现,随着功能期推进,叶片SP含量呈下降趋势,与马超等试验结果[27]相同,即随着功能期推进,叶片同化功能降低,衰老加剧,密度较高时早衰现象严重,对光合作用过程有重要贡献的二氧化碳固定酶也随着叶片的老化,被迅速分解,这是老化过程中叶片光合作用机能迅速减退的重要原因[28];前期SP含量高说明光合速率有利于生产与积累较多的光合产物,有较充足的源为籽粒灌浆提供重要的物质基础。
由于保护酶的反应是一种被动的适应,不能完全避免逆境伤害,而且在不同的时期和环境条件下,这种适应能力也不相同。总体来看,即使在高密度群体内,先玉335的叶片对轻中度水分胁迫仍有一定弹性,即使在高密度群体内,降低滴灌量,春玉米仍可通过株体内一系列生理生化反应来减轻或分散水分胁迫对细胞代谢过程的伤害,即在生育后期仍保持一定的抗氧化酶活性,MDA含量相对较低,且有一定的渗透调节物质维持细胞的正常渗透压,以保证玉米叶片正常的生理特性,减轻水分胁迫对细胞代谢过程的伤害。本研究仅对保护性酶类以及渗透调节相关物质在水密互作时随水分和密度变化的规律进行了初步探索,具体原因还有待于基因表达和物料合成方面的进一步深入研究阐明。
参考文献:
[1]王光涛,冯素伟,丁位华,等. 根际pH值对冬小麦叶片抗氧化酶活性和内源激素含量的影响[J]. 江苏农业科学,2021,49(10):71-75.
[2]Bacelar E A,Santos D L,Moutinho-Pereira J M,et al. Physiological behaviour,oxidative damage and antioxidative protection of olive trees grown under different irrigation regimes[J]. Plant and Soil,2007,292(1/2):1-12.
[3]黄智鸿,李秀娟,梁煊赫,等. 超高产玉米品种叶片保护酶活性及膜脂过氧化作用研究[J]. 江苏农业科学,2008,36(2):29-32.
[4]Hu T T,Yuan L N,Wang J F,et al. Antioxidation responses of maize roots and leaves to partial root-zone irrigation[J]. Agricultural Water Management,2010,98(1):164-171.
[5]Islam M R,Xue X Z,Mao S S,et al. Effects of water-saving superabsorbent polymer on antioxidant enzyme activities and lipid peroxidation in oat (Avena sativa L.) under drought stress[J]. Journal of the Science of Food and Agriculture,2011,91(4):680-686.
[6]Nie L X,Jiang X Y,Wu S H,et al. Effects of DA-6 on leaf photosynthetic carboxylase and protective enzyme activities and grain yield of high-yielding summer maize[J]. Journal of Applied Ecology,2010,21(10):2558-2564.
[7]蔡昆争,吴学祝,骆世明,等. 抽穗期不同程度水分胁迫对水稻产量和根叶渗透调节物质的影响[J]. 生态学报,2008,28(12):6148-6158.5A43E19C-6A53-4052-BC07-6D590CC13758
[8]周 磊,田军仓,沈 晖,等. 抽雄期水分胁迫对膜下滴灌玉米的影响研究[J]. 灌溉排水学报,2014,33(2):86-89.
[9]王俊峰,贾 涛,刘生荣. 灌水量和密度对转基因抗虫棉生育特性的影响[J]. 干旱地区农业研究,2007,25(5):64-68.
[10]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[11]王 群,刘朝巍,徐文娟. 紫外分光光度法测定玉米过氧化氢酶活性新进展[J]. 中国农学通报,2016,32(15):159-165.
[12]高俊鳳. 植物生理学实验指导[M]. 北京:高等教育出版社,2006.
[13]张宪政. 作物生理研究法[M]. 北京:农业出版社,1992.
[14]Tahi H,Wahbi S,El Modafar C,et al. Changes in antioxidant activities and phenol content in tomato plants subjected to partial root drying and regulated deficit irrigation[J]. Plant Biosystems,2008,142(3):550-562.
[15]申 林,黄智鸿,孙 刚,等. 超高产玉米与普通玉米叶片生理生化特性变化比较[J]. 安徽农业科学,2008,36(20):8453-8455.
[16]马秀芳,沈秀瑛,杨德光,等. 不同耐旱性玉米品种对干旱的生理生化反应[J]. 沈阳农业大学学报,2002,33(3):167-170.
[17]梁烜赫. 超高产玉米与普通玉米保护酶活性的比较研究[D]. 长春:吉林农业大学,2006:17-18.
[18]Zhang L J,Zhong T X,Xu L X,et al. Water deficit irrigation impacts on antioxidant metabolism associated with freezing tolerance increepingbentgrass[J].JournaloftheAmerican Society for
Horticultural Science,2015,140(4):323-332.
[19]闫素辉,李文阳,杨安中,等. 灌溉地和雨养地小麦旗叶生育后期抗氧化酶活性变化的比较[J]. 华北农学报,2011,26(1):162-166.
[20]石 岩,于振文,位东斌,等. 土壤水分胁迫对小麦根系与旗叶衰老的影响[J]. 西北植物学报,1998,18(2):196-201.
[21]孙彩霞. 玉米抗旱性鉴定指标体系及抗旱鉴定指标遗传特性的研究[D]. 沈阳:沈阳农业大学,2001.
[22]张玉荣,周显青,张 勇. 储存玉米膜脂过氧化与生理指标的研究[J]. 中国农业科学,2008,41(10):3410-3414.
[23]王 静,杨德光,马凤鸣,等. 水分胁迫对玉米叶片可溶性糖和脯氨酸含量的影响[J]. 玉米科学,2007,15(6):57-59.
[24]Wassom J,Reese R,Schumacher T,et al. Proline accumulation in response to dehydration and diurnal hydration cycles varies among maize genotypes[J]. Maydica,2000,45(4):335-343.
[25]汤章城. 植物对水分胁迫的反应和适应性:Ⅱ植物对干旱的反应和适应性[J]. 植物生理学通讯,1983,19(4):1-7.
[26]葛体达,隋方功,白莉萍,等. 水分胁迫下夏玉米根叶保护酶活性变化及其对膜脂过氧化作用的影响[J]. 中国农业科学,2005,38(5):922-928.
[27]马 超,黄晓书,李鹏坤,等. 种植密度对夏玉米果穗叶生理功能衰退的影响[J]. 玉米科学,2010,18(2):50-53.
[28]陈少裕. 膜脂过氧化与植物逆境胁迫[J]. 植物学通报,1989,24(4):211-217.5A43E19C-6A53-4052-BC07-6D590CC13758
