草地贪夜蛾V-ATPase亚基E、F、G和H基因的克隆与表达分析
2022-06-01邱琪琪王聪珂张阳王梦珂赵特周琳汪梅子
邱琪琪,王聪珂,张阳,王梦珂,赵特,周琳,汪梅子
(河南农业大学植物保护学院/河南省新型农药创制与应用重点实验室/河南省绿色农药创制工程技术研究中心,河南 郑州 450002)
液泡型ATP酶(vacuolar-type proton ATPase,V-ATP酶)定位在不同的膜中,能与离子通道和转运体结合维持适合所有真核细胞内膜隔室生化功能的pH环境,并激活许多不同的传输过程[1]。维持pH稳态对许多生理生化过程至关重要,包括细胞内的膜运输、激素原处理和神经递质运输,以及许多病毒进入细胞等。V-ATP酶是多亚基复合酶,由胞质内介导ATP水解的V1结构域和结合在膜上介导质子转移的V0结构域组成[2]。外周V1结构域包含至少8种分子质量为13~70 kD的不同亚基(亚基A~H),V0结构域包含6个分子质量17~100 kD的不同亚基(亚基a、c、c’、c”、d和e)[3]。V-ATP酶具有高度保守性,在真核细胞中通常位于质膜或酸性细胞器的膜中[4]。在许多昆虫的上皮细胞中,V-ATP酶分子大量存在于顶膜中产生一个电化学质子梯度,用于细胞外空间的酸化或碱化、离子和液体的分泌或重吸收、营养物质的输入、以及各种其他细胞活动[5]。
昆虫中活跃的跨上皮阳离子转运最初是在马氏管中被发现的,随后也在其他上皮中发现,如唾液腺、唇腺、中肠和感觉器,参与调节pH稳态[6]。昆虫的马氏管是排泄器官,作为运输组织当受到刺激时可以用比任何其他已知组织都要高的速度运输水和离子,这是由位于管腔细胞膜上的V-ATP酶驱动的[7]。V-ATP酶在昆虫其它组织中可能还发挥着其它重要作用。在黑腹果蝇Drosophilamelanogaster体内,位于复眼光感受器的V-ATP酶对两类板间神经元L1和L2的大小和形状进行了昼夜节律的周期性调节[8]。除此之外,V-ATP酶很可能参与了一部分信号通路的运行。在果蝇中,V-ATP酶不仅可以促进溶酶体中Notch配体的降解,而且促进核内体中Notch信号的激活[9]。在经过siRNAs靶向V-ATP酶的2个亚基(ATP6V1C2和ATP6V0C)处理的细胞中,Wnt信号通路的受体LRP6的磷酸化被抑制,这些结果表明Wnt信号通路的激活需要V-ATP酶的活性[10]。因其重要性,V-ATP酶被看作害虫防治的潜在分子靶标。
草地贪夜蛾属于世界性重大农业害虫,目前已有100多个国家受到了严重危害。草地贪夜蛾具有寄主范围广泛、多虫态共存、迁飞速度快、繁殖能力强和抗药性强等特点[11-12]。中国玉米种植广泛,草地贪夜蛾可能随着气候变迁及玉米播种时期不同进行迁移危害[13]。为探索草地贪夜蛾V-ATP酶V1结构域E、F、G和H亚基对草地贪夜蛾生长发育的影响,本研究采用PCR技术克隆了草地贪夜蛾V-ATP酶V1结构域的4个亚基(E~H)基因序列,通过实时荧光定量PCR技术测定了其在草地贪夜蛾不同发育阶段的表达动态。
1 材料和方法
1.1 材料
供试昆虫:草地贪夜蛾在温度(26 ± 1)℃、光照周期L/D=14 h/10 h、相对湿度60%~85%的室内条件下人工饲养。幼虫饲料的主要成分为玉米粉及玉米叶粉,成虫用10%蜂蜜水补充营养。
试剂:TRIzol试剂,美国Invitrogen公司;2×Es Taq Master Mix、大肠杆菌EscherichiacoliDH5α感受态细胞,北京庄盟国际生物基因科技有限公司;质粒DNA提取试剂盒,北京索莱宝科技有限公司;DNA凝胶回收试剂盒、PrimeScriptTMII 1st Strand cDNA Synthesis Kit,日本TaKaRa公司;PowerUpTMSYBRTMGreen Master Mix,赛默飞世尔科技(中国)有限公司。其他试剂均为国产分析纯。
仪器:DYY-6C电泳仪,北京六一仪器厂;In-GeniusLHR凝胶成像系统,美国Syngene公司;QuantStudio 3 Real-Time PCR System,美国ABI公司。
1.2 方法
1.2.1 总RNA提取与cDNA合成 收集草地贪夜蛾卵、1~6龄整头幼虫、雌蛹、雄蛹、雌成虫、雄成虫备用,每个时期1个生物学重复,所用的草地贪夜蛾数量分别为卵100粒,1龄40头,2龄30头,3龄15头,4龄、5龄各5头,6龄、雌雄蛹和雌雄成虫各3头。所有样品均设3次生物学重复。采用Trirol法提取各个龄期的RNA,取用适量RNA分别检测浓度,并通过琼脂糖凝胶电泳检测是否降解,剩余放置在-80 ℃冰箱保存备用。cDNA模板的合成参考PrimeScriptTMII 1st Strand cDNA Synthesis Kit说明书进行。
1.2.2 RT-PCR反应 从草地贪夜蛾转录组中获得V-ATP酶亚基的核苷酸序列,利用Primer Pre-mier 6.0软件设计出V-ATP酶V1结构域E、F、G和H亚基基因的4对引物(表1)。PCR采用50 μL体系:1.1×T3 Super PCR Mix 44 μL,10 μmol·L-1正向引物和反向引物各2 μL,cDNA模板2 μL。反应条件为:94 ℃预变性2 min;94 ℃变性10 s,60 ℃退火10 s,72 ℃延伸20 s,30个循环; 72 ℃延伸2 min;4 ℃保存。根据凝胶回收试剂盒说明书回收纯化PCR产物,将收集得到的PCR产物按照pClone007 Versatile Simple Vector Kit说明书进行目的片段连接,随后将连接产物转化到大肠杆菌DH5α感受态细胞,筛选阳性克隆并提取质粒DNA测序。引物合成及测序工作均委托郑州擎科生物有限公司完成。

表1 RT-PCR及qRT-PCR反应所用引物序列
1.2.3 序列分析和系统发育树的构建 对测序结果进行分析拼接后获得全长序列。使用NCBI的BLAST程序对V-ATP酶E~H亚基基因在不同物种间的同源性进行对比分析;用DNAMAN软件分析各个亚基基因的一致性;用MEGA X软件分析各个亚基与其他物种之间的进化关系;利用在线软件ExPASy-ProtParam tool进行各个亚基分子量、等电点等理化性质的分析;利用在线软件ExPASy-ProtScale tool进行各个亚基蛋白质的亲水性和疏水性分析。利用Pfam在线预测软件(http://pfam.janelia.org/search)对各个亚基蛋白结构域进行分析。
1.2.4 荧光定量RT-PCR分析 根据河南省新型农药创制与应用重点实验室已克隆出的草地贪夜蛾V-ATP酶E~H 4个亚基基因的序列全长设计出符合要求的4对特异性引物(表1)。为了消除样本量差异,以GAPDH基因作为内参基因[14]。
荧光定量PCR采用20 μL体系,各反应成分的含量为:PowerUpTMSYBRTMGreen Master Mix (2X) 10 μL,10 μmoL·L-1的正向和反向引物各2 μL,cDNA模板1 μL,ddH2O 5 μL。反应条件为:50 ℃ 2 min,95 ℃2 min;95 ℃ 15 s,60 ℃ 1 min,40个循环;溶解曲线95 ℃ 15 s,60 ℃ 1 min,0.15 ℃·s-1升温至95 ℃持续15 s。每个样品设置3个技术重复,反应结束后收集Ct值。采用2-△△Ct法计算相对表达量,以表达量最低的样品为基准进行定量分析。
1.3 数据分析
采用SPSS 19.0软件对试验数据进行统计分析,应用HSD法进行差异显著性检验。
2 结果与分析
2.1 核苷酸序列分析
通过特异性引物PCR扩增出编码草地贪夜蛾V-ATP酶V1结构域E~H 4个亚基的开放阅读框。E亚基的开放阅读框长度为681 bp,编码226个氨基酸,预测分子质量和等电点分别为55.5 kD和5.11,GenBank登录号为MT707618。E亚基基因编码的蛋白不稳定指数为40.97,属不稳定蛋白,蛋白亲疏水性预测显示为亲水性。该蛋白仅有一个保守结构域,即位于18~216的V-ATP酶E亚基结构域(图1)。

注:黑色表示高度保守的氨基酸,图2~4同。序列上方横线标注的是保守结构域:V-ATP酶E亚基结构域(FIE至ALF)。草地贪夜蛾:MT707618;斜纹夜蛾:XP_022815239.1;棉铃虫:XP_021191658.1;粉纹夜蛾:XP_026735678.1;家蚕:NP_001040451.1;夏威夷红蛱蝶:XP_026497418.1;野桑蚕:XP_028025362.1;烟草天蛾:XP_030031453.1;亚洲玉米螟:XP_028179329.1。
F亚基开放阅读框序列长375 bp,编码124个氨基酸,预测分子质量和等电点分别为30.8 kD和5.24,GenBank登录号为MT707619。F亚基基因编码的蛋白不稳定指数为53.33,属不稳定蛋白,蛋白亲疏水性预测显示为亲水性。该蛋白仅有一个位于12~113的V-ATP酶F亚基的保守结构域(图2)。

注:序列上方横线标注的是保守结构域:V-ATP酶F亚基结构域(ISV至LRR)。草地贪夜蛾:MT707619;斜纹夜蛾:XP_022825205.1;偏瞳蔽眼蝶:XP_023948003.1;夏威夷红蛱蝶:XP_026496172.1;家蚕:NP_001040448.1;脐橙螟:XP_013184521.1;粉纹夜蛾:XP_026737139.1;玉带凤蝶:NP_001298649.1;亚洲玉米螟:XP_028157779.1。
G亚基基因开放阅读框序列长354 bp,编码117个氨基酸,预测分子质量和等电点分别为27.7 kD和5.26,GenBank登录号为MT707620。G亚基基因编码的蛋白不稳定指数为44.68,属不稳定蛋白,蛋白亲疏水性预测显示为亲水性。该蛋白仅有一个保守结构域,即位于3~107的V-ATP酶G亚基结构域(图3)。

注:序列上方横线标注的是保守结构域:V-ATP酶G亚基结构域(SQT至DIK)。草地贪夜蛾:MT707620;斜纹夜蛾:XP_022818379.1;棉铃虫:XP_021188551.1;亚洲玉米螟:XP_028164911.1;夏威夷红蛱蝶:XP_026494470.1;柑橘凤蝶:NP_001299749.1;家蚕:NP_001040287.1;阿芬眼蝶:XP_034825880.1;野桑蚕:XP_023934297.1。
H亚基基因开放阅读框序列长1 452 bp,编码483个氨基酸,预测分子质量和等电点分别为118.2 kD和4.96,GenBank登录号为MT707621。H亚基基因编码的蛋白不稳定指数为41.01,属不稳定蛋白,蛋白亲疏水性预测显示为亲水性。H亚基蛋白具有2个保守结构域,分别是位于28~328和334~450的V-ATP酶F亚基N端结构域和C端结构域(图4)。

注:序列上方横线标注的是保守结构域:N端结构域(VLQ至ERL)和C端结构域(LSS至RNW)。草地贪夜蛾:MT707621;斜纹夜蛾:XP_022822595.1;粉纹夜蛾:XP_026726220.1;烟草天蛾:XP_030040379.1;大蜡螟:XP_026756478.1;家蚕NP_001040488.1;亚洲玉米螟:XP_028168669.1;夏威夷红蛱蝶:XP_026497418.1;柑橘凤蝶:XP_013180033.1;菜粉蝶:XP_022129728.1。
2.2 氨基酸序列同源性分析
草地贪夜蛾V-ATP酶E亚基与其他8种昆虫E亚基的氨基酸序列同源性分析结果显示,草地贪夜蛾V-ATP酶E亚基与斜纹夜蛾S.litura和棉铃虫H.armigera的V-ATP酶E亚基氨基酸同源性分别为97.8%和91.6%。通过多重序列比对发现不同物种间该基因编码氨基酸序列的差异少且分散。系统发育树结果显示(图5),草地贪夜蛾与斜纹夜蛾、棉铃虫聚集在同一进化分支上,有较高同源性。

图5 不同物种间V-ATP酶E亚基系统发育进化树
草地贪夜蛾V-ATP酶F亚基与其他8种昆虫F亚基的氨基酸序列同源性分析结果显示,草地贪夜蛾V-ATP酶F亚基与斜纹夜蛾的V-ATP酶F亚基氨基酸源性高达98.4%。通过多重序列比对发现不同物种间该基因编码氨基酸序列的差异不明显。系统发育树结果显示(图6),草地贪夜蛾与斜纹夜蛾聚集在同一进化分支上,有较高同源性。

图6 不同物种间V-ATP酶F亚基系统发育进化树
草地贪夜蛾V-ATP酶G亚基与其他8种昆虫G亚基的氨基酸序列同源性分析结果显示,草地贪夜蛾V-ATP酶G亚基与斜纹夜蛾、棉铃虫的V-ATP酶G亚基氨基酸同源性高达99.1%、96.6%。通过多重序列比对发现不同物种间该基因编码氨基酸序列前70位完全保守,差异主要存在于中间80~100位。系统发育树结果显示(图7),草地贪夜蛾与斜纹夜蛾、棉铃虫聚集在同一进化分支上,有较高同源性。

图7 不同物种间V-ATP酶G亚基系统发育进化树
草地贪夜蛾V-ATP酶H亚基与其他8种昆虫H亚基的氨基酸序列同源性分析结果显示,草地贪夜蛾V-ATP酶H亚基与斜纹夜蛾的V-ATP酶H亚基氨基酸同源性高达96.8%。通过多重序列比对发现不同物种间该基因编码氨基酸序列的差异少且分散。系统发育树结果显示(图8),草地贪夜蛾与斜纹夜蛾聚集在同一进化分支上,有较高同源性。
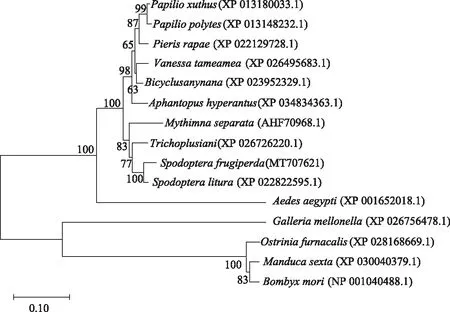
图8 不同物种间V-ATP酶H亚基系统发育进化树
2.3 草地贪夜蛾不同发育阶段V-ATP酶E~H亚基表达量分析
V-ATP酶E亚基基因在草地贪夜蛾整个发育阶段均有表达(图9)。其中在雄蛹中表达量最低,在雄成虫中高量表达,为雄蛹的12.44倍。另外,在1龄幼虫、4龄幼虫和雌性成虫中都有较高量的表达,这三者之间无显著差异。在雌蛹中表达量为雄蛹的1.27倍,两者无显著差异,均处于低水平。不同发育阶段中的相对表达量由高到低依次为雄成虫>1龄幼虫>6龄幼虫>4龄幼虫>2龄幼虫>雌成虫>3龄幼虫>5龄幼虫>卵>雌蛹>雄蛹。

注:数据为平均值±标准差。不同字母表示经HSD法检验在P<0.05水平差异显著。下同。
以表达量最低的雌蛹作为对照,草地贪夜蛾体内V-ATP酶F亚基基因的表达量在不同龄期存在差异(图10)。V-ATP酶F亚基基因在卵至3龄幼虫的表达量为雌蛹表达量的2.46~3.01倍之间,上下浮动不明显,无显著差异。从5龄开始表达量逐渐升高,直至6龄时期达到顶峰,为雌蛹表达量的6.82倍。雄蛹表达量明显高于雌蛹,两者之间差异显著。不同发育阶段中的相对表达量由高到低依次为6龄幼虫>雄成虫>雄蛹>5龄幼虫>雌成虫>卵>1龄幼虫>3龄幼虫>2龄幼虫>4龄幼虫>雌蛹。
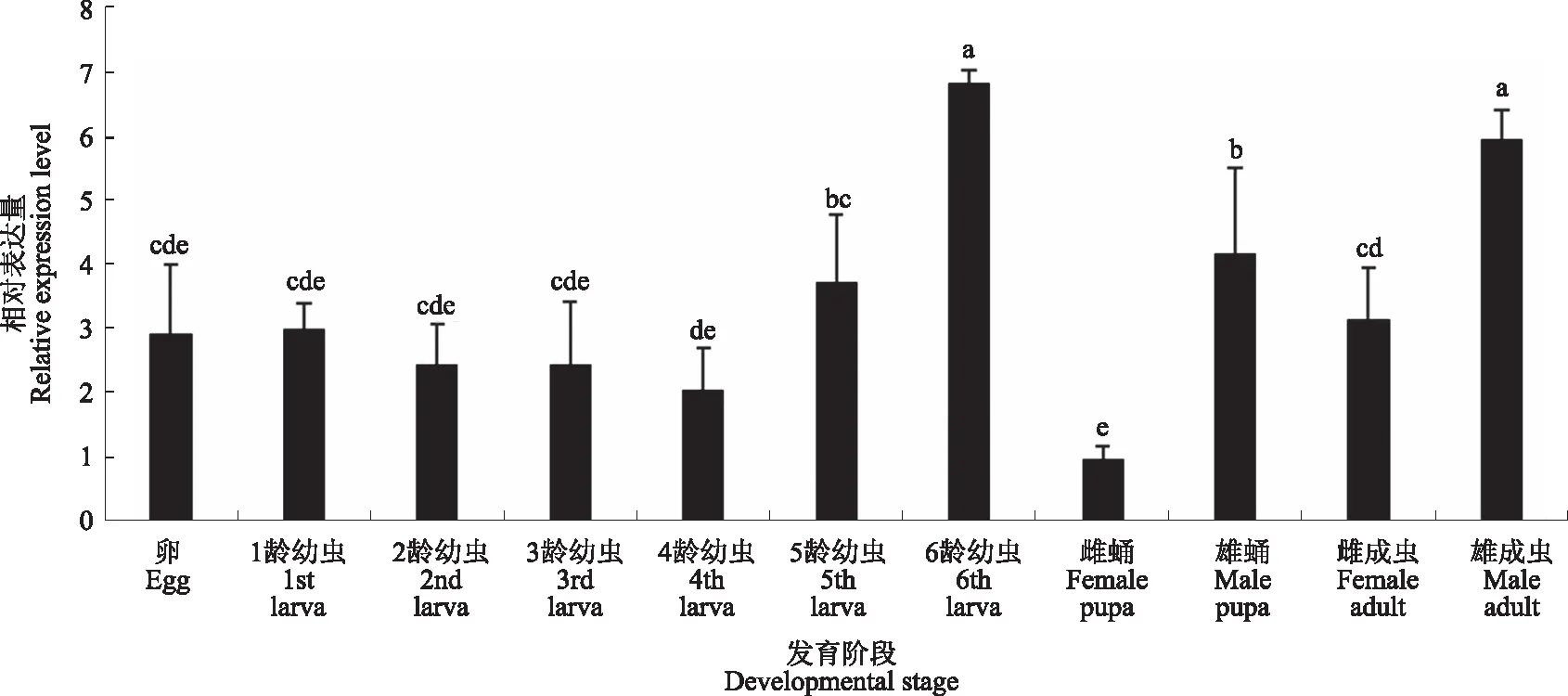
图10 V-ATP酶F亚基基因在草地贪夜蛾不同发育阶段的相对表达量
以表达量最低的雌蛹作为对照,对草地贪夜蛾不同发育阶段V-ATP酶G亚基基因的表达量进行分析(图11)。其中,在雄成虫中的表达量显著高于其他龄期,为雌蛹的18.28倍;在6龄幼虫中表达量也较高,为雌蛹的9.42倍。不同发育阶段中的相对表达量由高到低依次为雄成虫>6龄幼虫>1龄幼虫>雌成虫>3龄幼虫>2龄幼虫>5龄幼虫>4龄幼虫>卵>雄蛹>雌蛹。以表达量最低的雌蛹作为对照,草地贪夜蛾体内V-ATP酶H亚基基因的表达量在不同发育阶段存在差异(图12)。H亚基基因在卵和雄蛹中的表达量低于与其他龄期中的表达量,分别为雌蛹的2.22和2.26倍。雄成虫中H亚基基因的表达量明显高于其他龄期,为雌蛹表达量21.77倍,且与其他龄期表达量相比差异显著。幼虫期1龄至6龄的表达量之间均无显著差异。不同发育阶段中的相对表达量由高到低依次为雄成虫>6龄幼虫>雌成虫>3龄幼虫>1龄幼虫>4龄幼虫>2龄幼虫>5龄幼虫>雄蛹>卵>雌蛹。

图11 V-ATP酶G亚基基因在草地贪夜蛾不同发育阶段的相对表达量
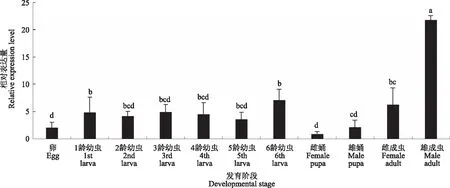
图12 V-ATP酶H亚基基因在草地贪夜蛾不同发育阶段的相对表达量
3 讨论
V-ATP酶的V1结构域被划分为几个子域:A3B3柱体、中心茎和外围茎,该结构域主要功能是水解ATP[15]。单一拷贝的F亚基通过其C端与D亚基结合从而组成中心茎,并且起到连接V1和V0结构域的作用[16]。外围茎由C、E、G、H亚基在中心茎外包裹形成。研究表明,H亚基可以在V-ATP酶V1和V0结构域的可逆解离过程中抑制V1结构域的催化活性[17]。昆虫的V-ATP酶亚基功能的研究主要依靠RNAi的方式进行,V-ATP酶的敲除对多种物种均有致死作用[18-20]。除此之外,V-ATP酶亚基的RNAi还会导致多种发育异常,如玉米蜡蝉(Peregrinusmaidis)的繁殖力降低和卵母细胞形成异常以及西方蜜蜂(Apismellifera)化蛹失败[21-22]。因此,V-ATP酶V1结构域的各个亚基很可能在昆虫的各种生理生化过程中承担着各不相同的重要责任。
本研究从草地贪夜蛾中成功克隆获得了V-ATP酶的E、F、G和H亚基基因。多序列比对和系统进化树的分析表明草地贪夜蛾中的V-ATP酶E~H亚基与其他物种的V-ATP酶E~H亚基具有极高的同源性。不同发育阶段V-ATP酶E~H亚基表达量分析结果表明,V-ATP酶E~H 4个亚基在草地贪夜蛾整个生长发育期均有表达,且都表现出在雄成虫中高量表达的特点。有研究表明,V-ATP酶在雄性灰翅夜蛾Spodopteramauritia输精管上皮细胞中参与调节管腔酸碱环境以维持精子正常发育[23]。V-ATP酶维持精子健康发育环境的功能不仅在昆虫中被发现,在哺乳动物、线虫中也得到了证实[24-25]。研究发现,桔小实蝇雄虫生殖节中的G亚基mRNA含量是雌虫生殖节中的6.04倍[26],也可以为此结果提供论据。E~H亚基在卵期和蛹期均表现为低量的表达,其原因在于处于这2个时期的个体飞行、进食等生命活动均不再进行,V-ATP酶只需维持低量运转就可以为孵化和羽化积蓄力量[27]。除此之外,个别亚基在6龄幼虫中也表现出较高的表达量,其中F和G亚基最为明显。6龄时期处于幼虫阶段末期,此时即将进入预蛹期各项生命活动都会降低,而F和G亚基却表现出较高的表达量,表明这2个亚基可能与化蛹相关。综上所述,V-ATP酶E、F、G和H均在昆虫的各种生理生化过程中承担着不可或缺的责任。
目前,V-ATP酶在昆虫的研究方向上还有许多空白,其与昆虫生长、发育和繁殖相关的调控机制尚需进一步明确。下一步拟将该V-ATP酶作为新型农药研发的候选靶标,通过深入了解V-ATP酶的作用方式,筛选出合理的作用靶点或开发相应的抑制剂,从而建立创新型农业害虫研究和防治手段。
