多浪羊CNP基因克隆及其在初情期组织表达特性分析
2022-06-01张志帅隋志远张继虎李庆瑾
张志帅,隋志远,张继虎,李庆瑾,邢 凤
(1.塔里木大学动物科学学院,阿拉尔 843300;2.新疆生产建设兵团塔里木畜牧科技重点实验室,阿拉尔 843300)
初情期为雌性动物初次发情并发生排卵的时期,与下丘脑-垂体-性腺轴的调控,环境和遗传因素对黄体生成素(LH)和卵泡刺激素(FSH)的协调功能的相互作用有关[1-5]。动物初情期的启动是其从幼年发育到获得繁殖能力的标志,初情期出现的快慢与其繁殖性能直接相关[6]。在母羊中,初情期的启动受内部和外部因素共同控制,主要取决于动物的年龄、体重和代谢状态,其他因素还包括品种、营养和环境因素[7]。初情期发育的正常或紊乱主要由遗传因素决定[8-9]。环境和代谢因素的神经内分泌调节在性腺发育中起着重要作用,但事实上这些因素均在基因调控的基础上发挥其功能[6]。据报道,下丘脑组织中的神经肽Y和瘦素系统[10]、神经激素B系统[11]、γ-氨基丁酸系统[12]、Lin28系统[13]等均与初情期启动有关。利钠肽家族由心房利钠肽(ANP)、脑利钠肽(BNP)和C型利钠肽(CNP)3种结构相关的肽组成[14]。CNP基因在多种组织中表达,并可能通过利钠肽受体(GC-B)作为局部自分泌/旁分泌的调节剂[15]。很多研究表明,CNP基因在生殖和胎儿生长发育中发挥着重要作用。Cameron等[16]研究发现,在小鼠性交后10.5 d的胎盘组织中CNP基因开始大量表达;Stepan等[17]研究发现,在妊娠前小鼠的卵巢以及子宫组织中CNP基因的表达量最高;CNP基因在大鼠卵巢和子宫中的表达与初情期的启动有关[18]。CNP基因的表达量对发情周期的依赖是由于子宫内容物、液体内容物质还有雌激素含量的变化[19]。在大鼠卵巢中,CNP基因的表达在初情期启动过程中表现出时间依赖性[20]。在啮齿动物中,CNP基因及其受体在子宫和卵巢中均高表达,在发情周期中具有特异性调节作用[21]。CNP基因及其受体鸟苷环化酶-利钠肽受体2 (NPR2)是环磷酸鸟苷(cGMP)稳态的关键调节因子。CNP-NPR2-cGMP信号级联在卵母细胞减数分裂的进程中起重要作用,这对雌性哺乳动物的繁殖至关重要[22]。CNP被认为是维持卵母细胞内环磷酸腺苷(cAMP)浓度的重要因素,cAMP浓度则是保持减数分裂阻滞直到排卵的重要因素[23],排卵前LH增加降低了体细胞中CNP的释放和cGMP的产生,从而降低了卵母细胞cAMP水平,促使减数分裂恢复[24-25]。在绵羊中,CNP预处理对绵羊卵母细胞成熟过程中母源基因DNA甲基化酶1(DNMT1)、合子阻滞因子1(Zar1)、胚胎必要的母体抗原(Mater)和卵丘细胞扩展相关基因透明质酸合成酶2(HAS2)、前列腺素内过氧化物合成酶2(PTGS2)、正五聚蛋白3(PTX3)的表达均有影响,可暂时阻止绵羊卵母细胞减数分裂的恢复,并提高绵羊卵母细胞成熟质量及早期胚胎发育能力[26]。
多浪羊是新疆早熟绵羊品种的典型代表,在正常生态条件下,其初情期年龄为3~4个月[27],CNP基因与减数分裂相关,但其在初情期启动过程中的影响机制还不清楚。探究CNP基因在多浪羊初情期启动过程中的影响机制对提高多浪羊的繁殖性能具有重要意义。因此,本研究通过克隆多浪羊CNP基因序列并进行测序,检测其在初情期前、初情期、初情期后3个不同发育期下丘脑、垂体、输卵管、卵巢、子宫中的表达水平,以期进一步揭示CNP基因在多浪羊初情期启动过程中的作用,为缩短多浪羊的育种周期提供理论依据。
1 材料与方法
1.1 材料
1.1.1 试验动物及样品采集 试验选用同一饲养条件下的初情期前(50.33±2.08) d、初情期(127±2.52) d、初情期后(154.28±2.30) d各3只健康的多浪羊,屠宰后采集新鲜下丘脑、垂体、输卵管、卵巢、子宫5个部位的组织样品,所有样品均于-80 ℃保存备用。试验所用多浪羊均来自塔里木大学试验站。母羊发情判断标准:兴奋不安,对外界刺激非常敏感,频繁排尿,外阴红肿有黏液,静立并接受公羊爬跨。
1.1.2 主要试剂 Trizol试剂盒、实时荧光定量PCR试剂盒2×SYBR®Premix ExTaqTM、pMD19-T载体均购自TaKaRa公司;DNA聚合酶、大肠杆菌DH5α感受态细胞、DNA回收试剂盒均购自天根生化科技(北京)有限公司;反转录试剂盒购自Thermo Fisher公司。
1.2 方法
1.2.1 引物设计与合成 参考GenBank中绵羊的CNP基因序列(登录号:XM_027974523.1),使用Primer Premier 5.0软件设计该基因CDS区序列特异性引物以及实时荧光定量PCR引物,引物信息见表1。引物均由生工生物工程(上海)股份有限公司合成。

表1 引物信息Table 1 Primer information
1.2.2 总RNA提取、反转录 按照Trizol法提取各个组织的总RNA,用1.0%琼脂糖凝胶电泳检测RNA条带的完整性。用PrimeScriptTMRT Reagent Kit反转录试剂盒反转录合成cDNA,置于-20 ℃保存。
1.2.3 基因克隆及测序 以下丘脑组织的cDNA为模板,用表1中的引物CNP-1、CNP-2、CNP-3对多浪羊CNP基因进行扩增。 PCR反应体系25 μL:Premix 12.5 μL,10 μmol/L上、下游引物各1 μL,cDNA 1 μL,ddH2O 9.5 μL。PCR扩增程序:95 ℃预变性5 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30s,共38个循环;72 ℃延伸5 min;4 ℃保存。用1.0%琼脂糖凝胶电泳检测PCR产物大小。PCR产物使用琼脂糖凝胶DNA回收试剂盒(TIANgel Midi Purification Kit)进行回收,pMD19-T载体进行连接,把连接的产物转化大肠杆菌DH5α感受态细胞,在摇床中37 ℃震荡50 min,接种于含有氨苄西林(Amp)的LB固体培养基,在37 ℃隔水培养箱中倒置培养,长出菌落后,挑取阳性菌落,进行菌落PCR鉴定(体系及程序同上)后,取鉴定正确的菌液送生工生物工程(上海)股份有限公司进行测序。将克隆所得CNP基因序列拼接成完整序列。
1.2.4 生物信息学分析 用DNAMAN软件对人(Homosapiens,登录号:NP_149124.3)、鸡(Gallusgallus,登录号:NP_990381.1)、猪(Susscrofa,登录号:XP_003131468.2)、牛(Bostaurus,登录号:XP_005220718.1)、马(Equuscaballus,登录号:XP_014595720.1)、驴(Equusasinus,登录号:XP_014689090.1)、山羊(Caprahircus,登录号:XP_017920590.1)、多浪羊(Dolang sheep)进行相似性比对,利用表2中的在线软件对多浪羊CNP基因序列及其编码蛋白进行生物信息学分析。

表2 生物信息学分析软件Table 2 Bioinformatics analysis softwares
1.2.5CNP基因组织表达分析 以不同初情期阶段多浪羊下丘脑、垂体、输卵管、卵巢、子宫5个组织的cDNA为模板进行实时荧光定量PCR分析。β-actin作为内参基因来校正个体差异。PCR反应体系15 μL:ddH2O 5.5 μL,PerfectStart Green qPCR SuperMix (2×) 7.5 μL,cDNA 1 μL,上、下游引物各0.5 μL。PCR反应程序:95 ℃预变性15 s;95 ℃变性15 s,55 ℃退火15 s,68 ℃延伸20 s,共40个循环。采用2-△△Ct法计算相对表达量。
1.3 数据统计及分析
用SPSS 22.0软件对数据进行单因素方差分析分析,2组之间的差异显著性比较用独立样本t检验,P<0.05表示差异显著。
2 结 果
2.1 多浪羊CNP基因的克隆与测序
1.0%琼脂糖凝胶电泳检测表明,CNP-1、CNP-2和CNP-3引物扩增所得目的条带大小分别在914、863和890 bp左右(图1),与预期目的片段大小相符。对多浪羊CNP基因先克隆并测序,后将序列进行拼接,所得序列大小为2 227 bp,包括5′-UTR 50 bp、3′-UTR 914 bp和CDS区1 263 bp,与预期结果一致。
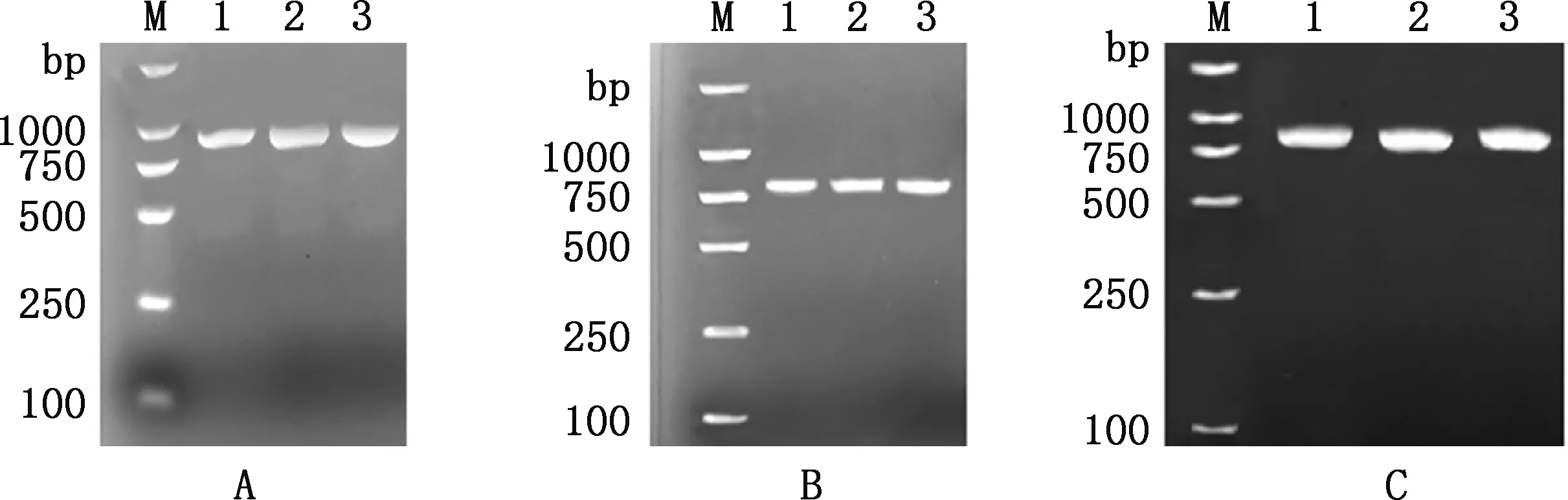
①A~C,分别代表由引物CNP-1、CNP-2、CNP-3所扩增的片段。②M,DL2000 DNA Marker;1~3,样品①A-C,Fragments amplified by primers CNP-1,CNP-2 and CNP-3,respectively.②M,DL2000 DNA Marker;1-3,Samples图1 多浪羊CNP基因PCR结果Fig.1 PCR results of CNP gene in Dolang sheep
2.2 CNP基因氨基酸序列相似性比较及系统发育树的构建
由图2可知,多浪羊CNP基因编码氨基酸序列与人、鸡、猪、牛、马、驴、山羊的相似性分别为70.7%、60.7%、90.0%、95.0%、87.9%、89.3%、97.9%。系统发育树结果显示,多浪羊与山羊、牛、猪的亲缘关系较近,与人、马、驴的亲缘关系次之,与鸡的亲缘关系最远(图3)。
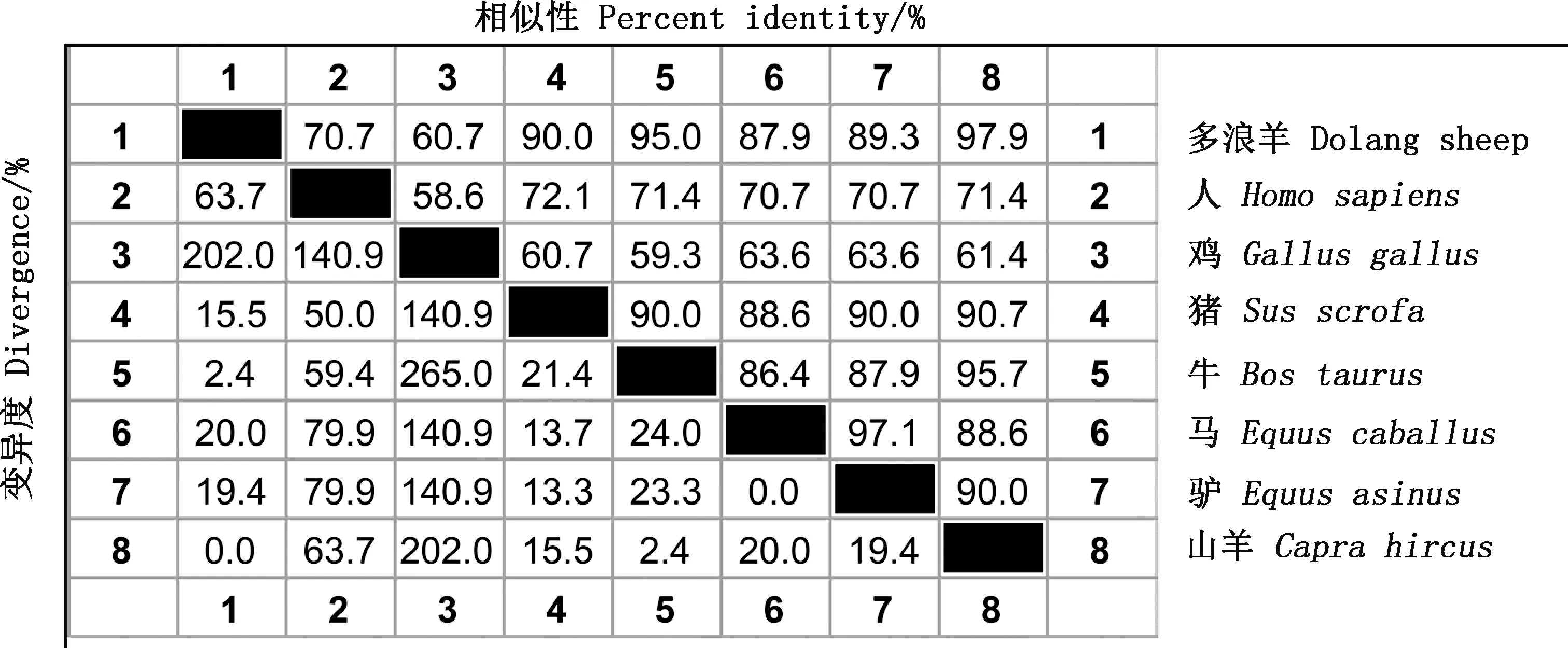
图2 多浪羊CNP蛋白氨基酸相似性比对分析Fig.2 Amino acid similarity alignment of CNP protein in Dolang sheep
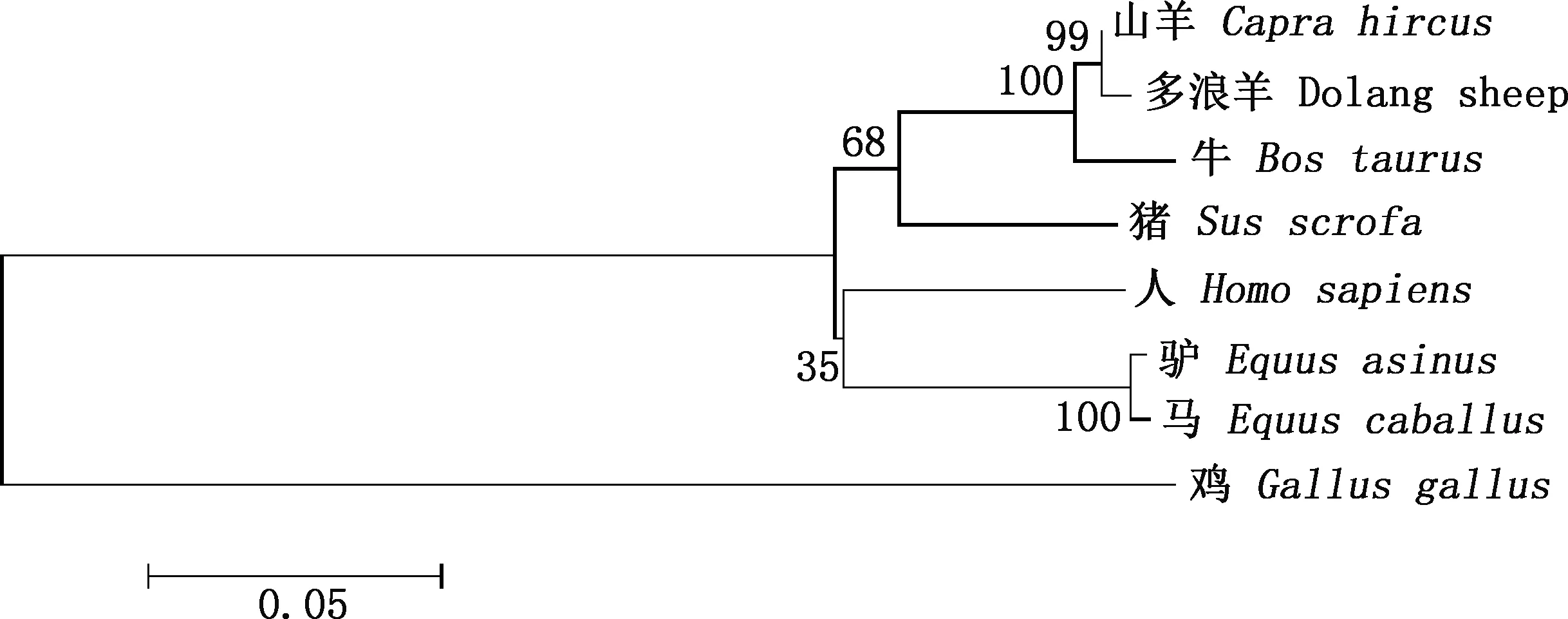
图3 基于CNP蛋白氨基酸序列的系统发育树Fig.3 Phylogenetic tree based on amino acid sequences of CNP protein
2.3 CNP蛋白的生物信息学分析
2.3.1 理化性质 通过在线分析软件ProtParam分析多浪羊CNP蛋白的理化性质显示,其分子质量为47 370.22 ku,理论等电点pI为9.13,分子式为C2114H3356N584O625S13,原子总数为6 692个,共编码420个氨基酸(表3),其中亮氨酸(L)有47个,比例最高,占11.2%,色氨酸(W)、Asn(N)、His(H)、Met(M)各有5个,比例最低,均占1.2%。带负电荷的氨基酸(Asp+Glu)残基总数为56,带正电荷的氨基酸(Arg+Lys)残基总数为68(表3);不稳定指数为43.02;利用在线分析软件ProtScale预测CNP蛋白的疏水性,由图4可知,得分小于0的区域比得分大于0的区域更密集,脂溶性系数为77.33;体外哺乳动物网织红细胞中半衰期为30 h,总平均亲水性(GRAVY)为-0.502,表明该蛋白为不稳定亲水碱性蛋白。
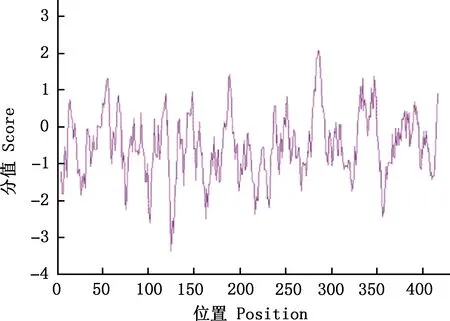
图4 多浪羊CNP蛋白质疏水性分析Fig.4 Hydrophobicity analysis of CNP protein in Dolang sheep

表3 多浪羊CNP蛋白氨基酸组成Table 3 Amino acid composition of CNP protein in Dolang sheep
用TMHMM软件对多浪羊CNP蛋白的跨膜结构域的预测显示,CNP蛋白质不含跨膜结构域(图5)。用在线软件SignalP 4.0预测多浪羊CNP蛋白质的信号肽,发现CNP蛋白不含有信号肽(图6)。用在线软件NetPhos 3.1对多浪羊CNP蛋白质的磷酸化位点进行预测,结果显示共有43个磷酸化位点,包括25个丝氨酸(S)位点、13个苏氨酸(T)位点、5个络氨酸(Y)位点(图7)。

图5 多浪羊CNP蛋白跨膜结构域预测Fig.5 Transmembrane domain prediction of CNP protein in Dolang sheep

图6 多浪羊CNP蛋白信号肽预测Fig.6 Signal peptide prediction of CNP protein in Dolang sheep

图7 多浪羊CNP蛋白磷酸化位点预测Fig.7 Phosphorylation sites prediction of CNP protein in Dolang sheep
2.3.2 二级结构及三级结构预测 用SOPMA软件对CNP蛋白二级结构的预测显示,该蛋白含有157个α-螺旋(37.38)、66个延伸链(15.71%)、18个β-折叠(4.29%)和179个无规则卷曲(42.62%);α-螺旋、延伸链和无规则卷曲贯穿于整个氨基酸链,β-折叠只有少量,散布在α-螺旋附近(图8)。使用SWISS-MODEL软件对CNP蛋白的三级结构预测显示,匹配到1个与CNP蛋白相似性为80.45%的同源序列2xmi.1.A,以它作为CNP蛋白的模板进行建模分析,结果显示,GMQE为0.45,QMEAN为-0.21,CNP蛋白无配体结构(图9)。
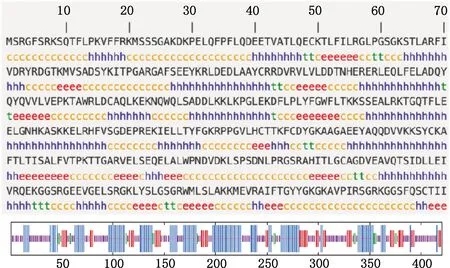
①h,α-螺旋;t,β-转角;c,无规则卷曲;e,延伸链。②线条按照从长到短依次代表α-螺旋、延伸链、β-转角和无规则卷曲① h,Alpha helix;t,Beta turn;c,Random coil;e,Extended chain.②The lines represent alpha helix,extended chain,beta turn and random coil,by the length,respectively图8 多浪羊CNP蛋白二级结构预测Fig.8 Secondary structure prediction of CNP protein in Dolang sheep

图9 多浪羊CNP蛋白三级结构预测Fig.9 Tertiary structure prediction of CNP protein in Dolang sheep
2.4 CNP基因在多浪羊不同初情期阶段不同组织中的表达分析
CNP基因在初情期启动的3个阶段下丘脑中表达量均显著高于在其他组织中对应发育阶段的表达量(P<0.05)。下丘脑中CNP基因在初情期的表达量显著低于初情期前和初情期后的表达量(P<0.05)。子宫中CNP基因在初情期前的表达量显著高于初情期和初情期后的表达量(P<0.05)(图10)。

①同一组织不同情期,肩标不同小写字母表示差异显著(P<0.05);肩标相同小写字母或无字母标注表示差异不显著(P>0.05)。②同一情期不同组织,肩标不同大写字母表示差异显著(P<0.01);肩标相同大写字母表示差异不显著(P>0.05)①Values of the same tissues, different period with different small letter superscripts mean significant difference (P<0.05); While with the same small letter or no letter superscripts mean no significant difference (P>0.05).②Values of the same period, different tissues with different capital letter superscripts mean significant difference (P<0.05); While with the same capital letter superscripts mean no significant difference (P>0.05)图10 多浪羊CNP基因在不同初情期阶段不同组织中的相对表达量Fig.10 The relative expression of CNP gene in different tissues at different puberty stages of Dolang sheep
3 讨 论
CNP基因与动物生殖系统功能关系密切,其具体作用机制是通过产生cGMP间接影响生殖细胞内cAMP的含量,从而影响生殖活动。CNP基因在雌性动物中的表达随发情周期的变化而变化,受多种内分泌激素及通路的影响[28]。CNP基因首次在猪脑提取物中被纯化和测序[29],是脑内高表达的利钠肽。本研究成功克隆了多浪羊CNP基因,并对其进行生物信息学分析。结果显示,CNP蛋白质的二级结构主要由α-螺旋和无规则卷曲组成,属于亲水性蛋白,没有信号肽和跨膜结构域,推测CNP蛋白不属于分泌蛋白,在合成后不发生转运。蛋白磷酸化是一种可逆的翻译后修饰,这种翻译后修饰可以改变蛋白的构象,进而使蛋白活化或者失活[30]。对CNP蛋白磷酸化位点进行预测发现了43个磷酸化位点,包括25个丝氨酸(S)位点、13个苏氨酸(T)位点、5个络氨酸(Y)位点。CNP蛋白氨基酸序列在不同物种间存在着高度保守性[31],对多浪羊CNP基因与其他物种的同源性分析以及构建系统进化树发现,多浪羊与山羊、牛、猪的亲缘关系较近,与人、马、驴的亲缘关系次之,与鸡的亲缘关系最远,与生物的进化特征相符,并且有广泛的同源性。
CNP基因是利钠肽家族的第3个成员,研究表明,CNP基因在小鼠和猪的卵母细胞减数分裂阻滞中起着维持作用[32],在排卵前的卵泡中,促黄体素诱导的由C型尿钠肽前体(natriuretic peptide precursor typeC,NPPC)编码的CNP基因表达下降是卵母细胞减数分裂恢复的先决条件[22]。研究表明,CNP基因在水牛卵母细胞周围的卵丘细胞中的表达量显著低于颗粒细胞[33];妊娠前的成年小鼠子宫和卵巢中CNP基因含量最高,超过前脑和脑干中CNP基因含量,在皮肤、舌头、心脏和肺脏中均有少量表达[17];在新生小鼠中,前脑和脑干中CNP浓度最高,在皮肤、舌头、心脏、肺脏、胸腺、骨骼肌、肝脏、肾脏、胃和颅骨中均有少量表达[17]。CNP基因在下丘脑和垂体前叶中也有表达[34]。本研究结果表明,初情期前下丘脑CNP基因表达量显著高于其他组织,这与在新生小鼠脑组织中的表达趋势一致。在大鼠发情期间,CNP基因参与了子宫中水与矿物质平衡及子宫肌层活动[35]。在山羊中,CNP基因可能参与调节卵泡发育和卵母细胞成熟[36]。本研究下丘脑和子宫中CNP基因在多浪羊初情期前到初情期的发育过程中表达量显著降低,证明CNP基因在多浪羊初情期启动过程中发挥着重要作用,这可能与CNP基因对减数分裂的抑制效果降低有关。综上所述,多浪羊CNP基因在下丘脑、垂体、输卵管、卵巢、子宫中均有表达,下丘脑组织中高表达,在初情期前子宫组织中也有较高的表达。这为进一步研究多浪羊CNP蛋白性质及其在多浪羊初情期启动中的调控机制奠定了理论依据。
4 结 论
本研究成功克隆了多浪羊CNP基因,其序列长度为2 227 bp,包括5′-UTR 50 bp,3′-UTR 914 bp和CDS区1 263 bp。CNP蛋白在进化中和物种间比较保守;多浪羊CNP基因主要在下丘脑和子宫中表达,且下丘脑中CNP基因表达量显著高于其他组织,在初情期前、初情期、初情期后的下丘脑组织中其表达量先降低再升高,提示其可能参与调控初情期的启动过程。
