贵州都匀下泥盆统蟒山群Zosterophyllum australianum(澳大利亚工蕨)新材料及其古地理意义*
2022-05-30周志强赵景宇王嘉树吴成洋
周志强 赵景宇 王嘉树 吴成洋 肖 彪 黄 璞
1宿州学院资源与土木工程学院,安徽宿州 234000 2北京大学地球与空间科学学院,北京 100871 3中国科学院南京地质古生物研究所,江苏南京 210008
Zosterophyllum(工蕨属)是由Penhallow于1892年依据苏格兰下泥盆统中的标本材料建立的一个化石植物属,最早出现在志留纪晚期,繁盛于早泥盆世,是早期陆生植物群落中的优势和代表性类群(Kenrick and Crane,1997;Kotyketal.,2002;Hao and Xue,2013)。该属广布全球,在中国、哈萨克斯坦、欧洲、北美和澳大利亚等国家和地区均有着丰富的化石记录(Lang and Cookson,1930;Jurina,1969;Edwards,1975;Kotyketal.,2002;Hao and Xue,2013;Gossmannetal.,2021)。尽管对Zosterophyllum的鉴别特征存在不同的认识(Kotyketal.,2002;Edwards,2006),但其重要的特征包括:营养枝光滑,形成H型或K型分枝;球形、椭圆形至肾形的孢子囊侧生于裸轴之上,并形成孢子囊穗;孢子囊沿远端开裂带分裂成2瓣等(Hao and Xue,2013)。根据孢子囊在茎轴上的排列方式,Croft和Lang(1942)将Zosterophyllum划分为Euzosterophyllum(真工蕨亚属)和Platyzosterophyllum(扁工蕨亚属)。Euzosterophyllum(后改为Zosterophyllum)以孢子囊螺旋着生于茎轴为特征,而Platyzosterophyllum的孢子囊则着生于茎轴一侧(Hueber,1972)。目前为止,全球已经命名和描述该属20多个种,其中大部分种被认为是客观有效的(Hao and Xue,2013;Gossmannetal.,2021)。Zosterophyllum具有世界广布、时间延限相对较短等特点,是研究泥盆纪生物地层学(特别是非海相地层)和古地理学的重要实证材料。
Zosterophyllumaustralianum(澳大利亚工蕨)最早发现于澳大利亚维多利亚州(Lang and Cookson,1930),之后Cookson(1935)报道了该州另一个剖面中的Z.australianum。Croft和Lang(1942)描述了英国下老红砂岩中的Zosterophyllumcf.Z.australianum(澳大利亚工蕨相似种)。Cookson(1949)在澳大利亚维多利亚州发现有该植物的孢子囊穗和分散保存的孢子囊,并认为该孢子囊也属于Z.australianum。Jurina(1969)报道并描述了哈萨克斯坦的Z.australianum化石。中国的Z.australianum材料则最早发现于云南文山坡松冲组中(李星学和蔡重阳,1977)。Hao(1992)对产自云南曲靖徐家冲组的Z.australianum进行了详细研究,并获取了该植物的原位孢子。Hao和Gensel(1998)根据华南板块与澳大利亚共有Z.australianum、Hedeia和Baragwanathia等植物属种的证据,结合一些古地理重建中这两个地区在早泥盆世相距较近的观点,提出“东北冈瓦纳古植物地理区系”(“Northeastern Gondwana Phytogeographic Unit”)这一概念。郝守刚和王德明(2000)详细描述了采自云南文山坡松冲植物群中保存较好的Z.australianum,并进一步阐述东北冈瓦纳古植物地理区系的特征,认为在该植物区系中Zosterophyllum亚属占优势地位,而Platyzosterophyllum亚属的成员则多见于劳俄大陆。

图1 早泥盆世全球古地理图(A)(修改自Huang et al.,2018)及贵州都匀包阳剖面交通位置图(B,C)Fig.1 The Early Devonian paleogeographic map(A)(modified from Huang et al.,2018)and traffic map of Baoyang section in Duyun,Guizhou(B,C)
文中描述了采集自贵州都匀包阳剖面蟒山群中的Zosterophyllumaustralianum标本。该化石保存较为完整,清晰地展示了孢子囊穗的整体形态和孢子囊特征。笔者将其与其他产地的Z.australianum的形态学特征进行了对比和总结,并依据该种的延限为蟒山群下段的时代提供约束。另外,结合华南同时代多个植物群的资料,通过网络分析(Network Analysis)手段,讨论了Z.australianum在生物古地理学方面的意义。
1 材料与方法

图2 贵州都匀包阳剖面蟒山群岩性柱状图及化石产出层位Fig.2 Lithological column of the Mangshan Group at Baoyang section in Duyun,Guizhou,showing fossil-bearing beds
文中的Zosterophyllumaustralianum新材料采自贵州都匀包阳剖面蟒山群,GPS点位为 26°6′44″N、107°26′44″E(图1)。依据戴传固等(2017)对本区域地层的划分观点及野外观察,文中将包阳剖面地层划分为志留系翁项组、泥盆系蟒山群和独山组鸡泡段。其中,蟒山群以灰白色石英砂岩为主,夹灰白色、灰黑色泥岩和粉砂岩。包阳剖面包括A、B共2个亚剖面,相距约200 m,分别位于背斜的两翼。根据相对位置和延展方向,推测亚剖面A可能位于亚剖面B之下(图2)。包阳剖面产出丰富的动植物化石,动物化石包括:鱼类Duyunolepissp.(都匀鱼未定种)(图3-A)、存疑的胴甲鱼及鱼碎片(图3-C,3-D)、腕足动物“Lingula”(舌形贝)(图3-B)、节肢动物碎片(图3-E,3-F,3-G)及遗迹化石Bifungitesisp.(双菌迹未定种)(图4-A)。这些鱼类、腕足动物及节肢动物通常被解释为海洋动物。含植物化石Z.australianum的层位位于亚剖面B的下段。植物化石围岩为灰色泥岩,部分围岩由于浸染氧化等原因,呈灰红色团块(图4)。化石以压型形式保存。所有标本均保存于中国科学院南京地质古生物研究所标本馆中。
由于植物繁殖枝的部分茎轴埋藏在围岩之中,使用气修针除去周围的围岩。使用数码相机对化石标本进行拍照,孢子囊穗的细节照片使用体式显微镜Axio Zoom V16进行拍照。所有照片仅调节对比度和亮度,以增大标本与围岩之间的反差。使用绘图软件CorelDRAW绘制孢子囊穗线条图。使用软件ImageJ 1.44测量化石的相关性状(表1)。
为了分析华南早泥盆世几个重要组段中植物属种的古地理分布特征,文中使用软件Gephi 0.9.2进行网络分析(具体原理及方法请参见王骞和黄冰,2020)。涉及的植物群包括:云南文山早泥盆世坡松冲植物群和曲靖徐家冲植物群、四川江油平驿铺植物群及文中研究的蟒山群植物组合。分析所用数据来源于耿宝印(1992a,1992b)、Wang 等(2002)、Hao和Xue(2013)、Edwards等(2016)、Edwards和Li(2018a,2018b)、Xue等(2018)等相关文献,其中相似属种和未定种均不纳入本次统计和分析范畴(数据参见表 2和表3)。网络分析是一种新的数据可视化途径,用以呈现复杂系统中组成分子之间的关系模式(王骞和黄冰,2020),近几年来在古生物地理学相关研究中逐渐受到重视。文中引入这种方法,对华南早泥盆世植物群展开分析。

A—Duyunolepis sp.(都匀鱼未定种),标本号为NIGP179710,比例尺为5 mm;B—“Lingula”(舌形贝),标本号为NIGP179711,比例尺为2 mm;C—与“Lingula”同层保存的存疑的鱼类碎片,标本号为NIGP179712,比例尺为5 mm;D—与Zosterophyllum australianum同层保存的存疑的胴甲鱼碎片,标本号为NIGP179713,比例尺为10 mm;E-G—与Z. australianum同层保存的节肢动物碎片,标本号为NIGP179714-179716,比例尺分别为5 mm(E、F)和2 mm(G)图3 贵州都匀包阳剖面蟒山群中的动物化石Fig.3 Animal fossils from the Mangshan Group at Baoyang section in Duyun,Guizhou
2 系统描述与比较
新标本的繁殖枝较为完整,长度可达141.0 mm(含顶端的孢子囊穗),未见分枝。茎轴光滑,向上逐渐弯曲,最宽处约为2.0 mm(图4-A,4-B)。孢子囊穗着生于繁殖枝顶端,长约16.2 mm,宽可达4.5 mm。孢子囊穗由穗轴和螺旋排列的孢子囊组成,可以观察到至少22个孢子囊紧密螺旋排列(图4-C,4-D;图5-G)。孢子囊呈椭圆形或扇形,宽2.6~4.1 mm,高可达3.9 mm(图4-C,4-D,4-E,4-H,4-I)。囊穗顶端的孢子囊明显变小(图4-E),推测该孢子囊尚未成熟。孢子囊加厚带明显,宽可达0.6 mm(图4-H,4-I),沿远端分裂(图4-F)。孢子囊具短柄(图4-G),柄长0.7 mm、宽约1.1 mm,以45°~53°斜插于中央穗轴之上。

A,B—Zosterophyllum australianum的正反面标本,孢子囊穗着生于茎轴顶端。数字1至8所指为遗迹化石Bifungites isp.,标本号为PB24570,比例尺为10 mm;C,D—为A、B中孢子囊穗的放大,箭头e-i分别放大于图E-I,比例尺为2 mm;E—孢子囊穗顶端的孢子囊,比例尺为1 mm;F—孢子囊顶端加厚带开裂,比例尺为500 μm;G—孢子囊具短柄,比例尺为1 mm;H—具明显的孢子囊加厚带,比例尺为1 mm;I—孢子囊,底部箭头所指为孢子囊的一瓣,上部箭头所指为加厚带,比例尺为500 μm图4 贵州都匀包阳剖面蟒山群中的Zosterophyllum australianumFig.4 Zosterophyllum australianum from the Mangshan Group at Baoyang section in Duyun,Guizhou

表1 华南板块Zosterophyllum australianum 测量数据对比表Table 1 Measurements of Zosterophyllum australianum from South China Block

表2 华南板块上扬子地区早泥盆世植物大化石各属与产地Table 2 Genera and localities of plant macrofossils of the Early Devonian from the Upper Yangtze in South China Block

表3 华南板块上扬子地区早泥盆世植物大化石各种与产地Table 3 Species and localities of plant macrofossils of the Early Devonian from the Upper Yangtze in South China Block
尽管不少学者认为Zosterophyllum的大部分种是有效的,但是各个种之间的区分仍有一定难度。区别和鉴定Zosterophyllum各个种的特征,包括营养枝的分枝形式、孢子囊的排列形式、孢子囊着生角度、孢子囊形态大小、孢子囊柄长度、加厚带有无等。由于文中报道的化石材料显示出孢子囊呈螺旋排列这一典型特性,毫无疑问其隶属于Zosterophyllum亚属,而与Platyzosterophyllum亚属的成员在孢子囊排列方式上截然不同。文中材料的孢子囊排列紧密,这与孢子囊松散或较为松散排列的Z.bifurcatum、Z.deciduum、Z.dushanense、“Z.longhuashanense”、Z.mi ̄nor ̄sta ̄chyum、Z.myretonianum、Z.ova ̄tum、Z.ra ̄mosum、Z.spa ̄thula ̄tum、Z.shengfengense、Z.sinense、Z.tene ̄rum、Z.yun ̄nanicum不同(徐仁,1966;Edwards,1975;李星学和蔡重阳,1977;Gerrienne,1988;郝守刚和王德明,2000;Xue,2009;Haoetal.,2010;Hao and Xue,2013;Edwards and Li,2018b;汪瑶和徐洪河,2018)。尽管Zosterophyllum的部分种也显示出孢子囊紧密排列的习性,如Z.confertum和Z.minifertillum,但其与这些植物在分枝形式、孢子囊穗形态和大小、孢子囊大小、加厚带宽度等方面仍有所区别(Hao and Xue,2013;Gossmannetal.,2021)。
与文中描述的标本最为接近的物种是Zosterophyllumaustralianum,特别是郝守刚和王德明(2000)描述的产自云南坡松冲植物群中的该种标本。Lang和Cookson(1930)最初描述Z.australianum时,认为该植物“bearing around its upper portion the spirally arranged and closely-crowded appendages…the sporangia are apparently terminal on short stalks”,也就是说该植物具有孢子囊螺旋排列、形成紧密的孢子囊穗、孢子囊具短柄等特征(如图5-A)。这些特征被认为是鉴别Z.australianum的关键特征,但囿于该种的原始标本保存较差,且相关营养结构并不清楚,也为后期准确鉴定Z.australianum制造了较大困难,有关认识也多有分歧。Edwards(1975)认为Jurina(1969)中的大部分标本(如No.48/490)的化石(图5-F)与Z.myretonianum相似。Hao(1992)认为Cookson(1935)、Croft和Lang(1942)、Jurina(1969)中的部分标本与原始标本相比,在孢子囊大小、孢子囊柄的尺寸等方面有所不同(如图5-B,5-C,5-D,5-F),推测这些标本很可能不属于Z.australianum。以前在中国报道的Z.australianum材料同样存在争议,即Hao(1992)认为李星学和蔡重阳(1977)中描述的Z.australianum和“Z.contiguum”均属于Z.australianum,但后者被Li和Edwards修订,建立为新组合Demersathecacontigua(Li and Edwards,1996),而前者中部分标本也被修订为D.contigua(Wang Yetal.,2020)。本文通讯作者对馆藏于中国科学院南京地质古生物研究所的这批标本(标本号:PB6467-6475)进行观察,赞同将李星学和蔡重阳(1977)描述的部分Z.australianum处理为D.contigua这一观点。其余各地报道的Z.australianum标本,保存状态均较差,无法鉴别,如Cookson(1949)记述的一些分散孢子囊(图5-E)。
产生上述问题的根源在于Zosterophyllumaustralianum的原始标本信息较为有限,亟待对原始标本开展更为详细的观察和研究,并采集更多原始产地的新标本来丰富分类学上的信息。与原始标本形成鲜明对比的是,云南下泥盆统徐家冲组和坡松冲组中的标本则展示出了更为全面的信息,特别是营养结构。云南文山菖蒲塘剖面坡松冲组的Z.australianum茎轴长达110.0 mm,宽1.0~2.8 mm,等二歧或不等二歧分枝;孢子囊紧密螺旋排列形成孢子囊穗,穗长13.0~15.2 mm、宽4.3~6.8 mm;孢子囊前视观为椭圆形或扇形,成熟时孢子囊高2.6~3.6 mm、宽3.5~4.5 mm;孢子囊柄长0.6~0.8 mm,宽0.6~1.4 mm;孢子囊顶端具有明显的加厚带,可达0.9 mm宽(郝守刚和王德明,2000;表 1)。文中描述的标本除营养枝的分枝特征尚不清楚、孢子囊穗的长度略有差距外,其余性状均与坡松冲组的标本非常接近。与坡松冲组中的标本显示出多次等二歧或不等二歧分叉情况不同,文中描述的Z.australianum的繁殖枝未见分叉,长度可达141.0 mm,这显示出了该植物的繁殖枝在分枝形式上的多样化。
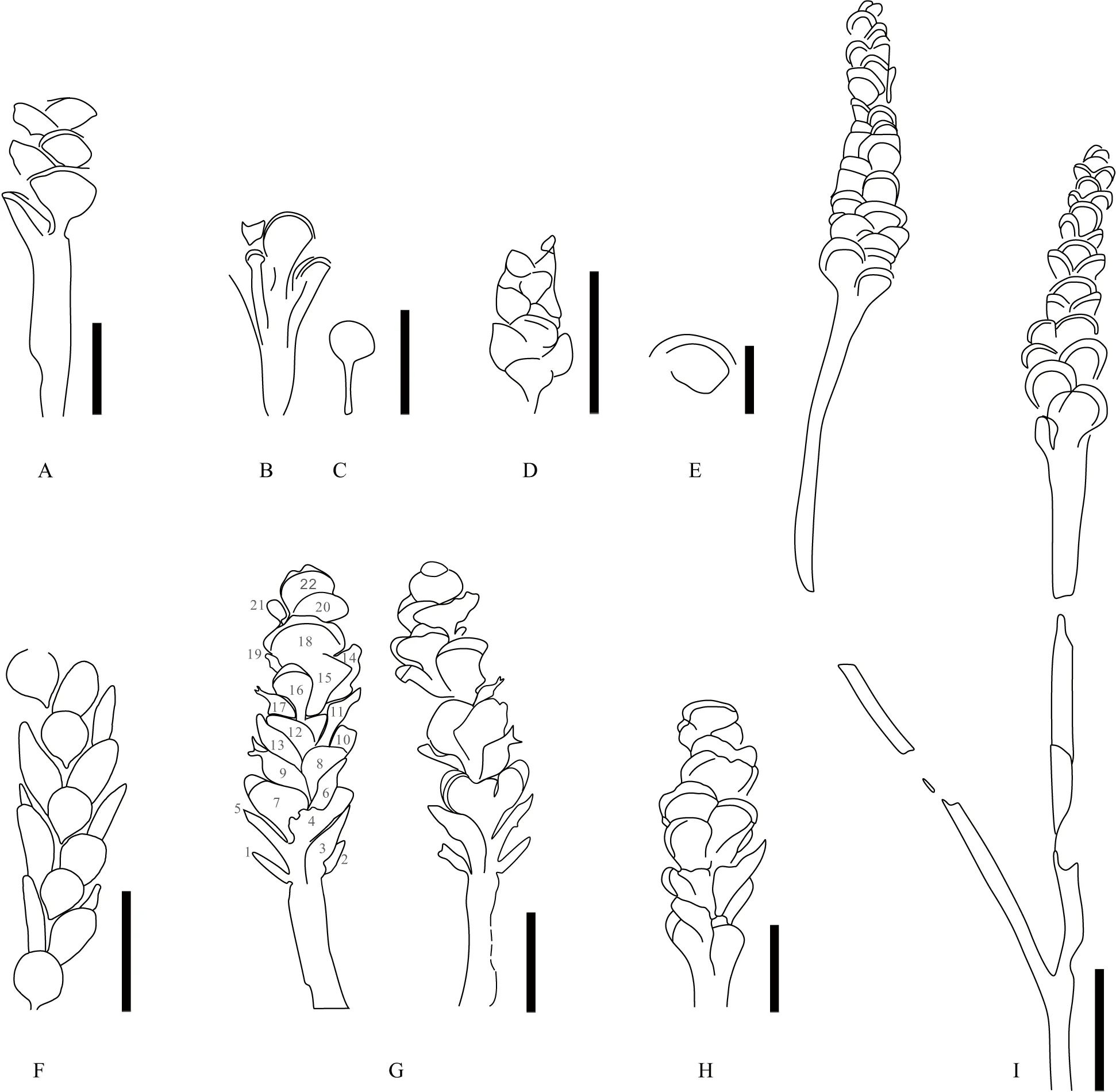
A—澳大利亚维多利亚州North Road石矿中的Zosterophyllum australianum的线条图,依据Lang and Cookson(1930)图版12中的 图16 绘制,比例尺为10 mm;B,C—澳大利亚维多利亚州Pleasant山中的Z. australianum的线条图,依据Cookson(1935)图版10中的图10、11绘制,比例尺为 5 mm;D—英国Llanover石矿中的Z. cf.Z. australianum,依据Croft和Lang(1942)图版10中的 图30 绘制,比例尺为10 mm;E—澳大利亚维多利亚州Hull Road的Z. australianum的分散孢子囊,依据Cookson(1949)图版Ⅳ中的图7 绘制,比例尺为5 mm;F—哈萨克斯坦的Z. australianum,依据Jurina(1969)插 图7 的线条图绘制,比例尺为10 mm;G—本文标本(贵州都匀包阳剖面),据图4-C和4-D,比例尺为5 mm;H—云南文山坡松冲组中的Z. australianum,依据郝守刚和王德明(2000)的图1A绘制,比例尺为5 mm;I—云南曲靖徐家冲组中的Z. australianum,依据Hao(1992)的 图32 绘制,比例尺为10 mm图5 不同产地Zosterophyllum australianum的线条图Fig.5 Line drawings of Zosterophyllum australianum from different localities
3 讨论
3.1 蟒山群的时代及沉积环境
贵州都匀、独山一带的泥盆系序列较为完整,是中国泥盆系研究的重点区域之一(廖卫华等,1978;戴传固等,2017;郄文昆等,2020)。相较于海相泥盆系的研究,非海相地层学的研究相对较弱,部分典型剖面的地层划分仍存在较大争议,有关时代的认识尚不统一。以贵州都匀包阳剖面为例,地层划分的方案也因各家认识不同而有所争议,目前对于包阳剖面不整合于志留系之上的碎屑岩主要有3种划分方案:第1种方案将这套砂岩与邦寨砂岩进行对比,认为属于(上)邦寨组(胡炎坤,1964;地质部贵州省地质局区域地质队,1965);第2种方案是根据贵州独山县猴儿山的典型剖面,将包阳剖面的下泥盆统划分为丹林组(群)和舒家坪组,认为它们的时代为布拉格期到埃姆斯期(刘向,内部资料;高联达和侯静鹏,1975;沈志达和曾羽,1988;陈代钊和陈其英,1994);第3种方案将该地区志留系之上的泥盆系碎屑岩全部划入蟒山群(组)中(戴传固等,2017)。第1种方案中采用的邦寨组的命名剖面的具体位置并不清楚,且划分界限不明,而后来认定的典型剖面中的邦寨组又存在断层,造成认识分歧较大(王钰和俞昌民,1962;王钰等,1964;廖卫华等,1978;侯鸿飞等,2000)。第2种划分方案曾被众多学者所采纳,但是在包阳剖面中划分丹林组和舒家坪组颇为不易。基于以上原因,目前本文采用第3种方案。
蟒山群最初由乐森璕(1929)根据都匀城外的蟒山石英砂岩而命名,并依据其他地区剖面中相当层位中的化石证据,推测其时代为早、中泥盆世(江能人,1958;王钰和俞昌民,1962;刘向,内部资料)。黄璞等(待刊出)根据在包阳剖面蟒山群下段发现的Adoketophytonsubverticillatum,认为蟒山群下段的时代很可能为布拉格期。Zosterophyllumaustralianum在华南主要分布在云南坡松冲组和徐家冲组。坡松冲组的时代根据孢粉组合被认为是布拉格期(Cascales-Mianaetal.,2019)。尽管徐家冲组上部的时代为布拉格期至最早埃姆斯期,但Z.australianum发现于该组的底部(Wellmanetal.,2012)。综合以上证据,笔者认为蟒山群下段的时代为早泥盆世布拉格期。
当前材料的孢子囊细节保存欠佳,表明其经历了一定距离的搬运。孢子囊随陆源碎屑搬运的过程为滨海或河口地区提供了丰富的营养物质,大量以悬浮物为食的生物富集于此,这也是大量Bifungitesisp.与Z.australianum同层保存的原因。同时,这些以悬浮物为食的生物吸引了大量的捕食者,比如一些鱼类和节肢动物,共同构成了复杂的生态链(图3;图4-A)。因此,蟒山群植物化石层附近层位的沉积环境可能为滨海或海陆过渡环境(如河口湾)。
3.2 古地理学意义
华南板块具有丰富的下泥盆统非海相地层,其中已报道的含植物化石的产地至少有21处(蔡重阳和王怿,1995),但从古植物学角度深入研究过的化石主要集中于云南早泥盆世坡松冲植物群和徐家冲植物群、四川平驿铺植物群以及广西石桥苍梧组中的植物(李星学和蔡重阳,1977;耿宝印,1992a,1992b;Wangetal.,2002;Hao and Xue,2013;Edwardsetal.,2016;Edwards and Li,2018a,2018b;Xueetal.,2018;Wang Yetal.,2020)。目前云南坡松冲植物群中已报道28属陆生植物(不含相似属),远超同一时期其他板块特定组段或化石点所报道的化石植物数目(Guoetal.,2012;Hao and Xue,2013;表 2),云南曲靖徐家冲植物群、四川平驿铺植物群中分别发现陆生植物7属和16属(表 2)。这些化石材料表明,华南板块是早期陆生植物多样性发育的中心之一。同时,这些植物显示出了浓重的地方性色彩,在属级、种级分类单元上,地方性分子远远多于全球广布分子(Xiongetal.,2013;Xueetal.,2018),这可能是因为华南板块在当时处于泛大洋,缺少与其他板块之间的直接联系,物种传播受到阻隔所致。华南板块与其他板块的植物化石在属一级上,仅共有Baragwanathia、Distichophytum、Drepanophycus、Estinnophyton、Hedeia、Psilophyton、Oricilla和Zosterophyllum等,而在种一级上,则少有共同种。以Zosterophyllum为例,尽管Zosterophyllum在早泥盆世广布于全球,从志留纪晚期一直延续到了埃姆斯期,在华南也有广泛的分布,但华南板块与其他板块(澳大利亚)仅共有Z.australianum和Z.ramosum(Kotyketal.,2002;Hao and Xue,2013;Gossmannetal.,2021)。
多项研究表明,Zosterophyllum中的不同种在古板块上的分布存在差异(Hao and Gensel,1998;郝守刚和王德明,2000;Hao and Xue,2013)。华南的Zosterophyllum以孢子囊螺旋着生为主,而劳俄大陆上的Zosterophyllum则存在孢子囊单侧着生的习性,后者代表性的物种包括Z.llanoveranum和Z.divaricatum等。尽管其他板块上也存在一些存疑的Z.australianum,但是该种在华南与澳大利亚的分布最为广泛。除此之外,华南与澳大利亚还同时分布有Z.ramosum、Hedei、Baragwanathia等属种,这一现象被解释为二者在早泥盆世期间相隔较近,在物种交流上保持了较强的联系,形成特有的植物区系(东北冈瓦纳古植物地理区系)。文中在贵州都匀包阳剖面发现的Z.australianum进一步支持了东北冈瓦纳古植物地理区系这一概念。但必须指出,由于澳大利亚的Z.australianum原始标本较为破碎,研究程度不够,亟待开展进一步研究。

A—华南板块早泥盆世上扬子地区古地理复原图(修改自Edwards等,2016);浅绿色、黄色和蓝色部分分别代表陆地、河口和浅海;B,C—华南板块早泥盆世代表性植物群的属级、种级分类单元的网络图。图中的大圆代表植物化石产出的组段,包括:PSC,坡松冲组;XJC,徐家冲组;PYP,平驿铺群;MS,蟒山群。图中的小圆代表不同的植物属(B)或种(C);2个或多个组段共有的植物属、种(小圆),位于各组段的中间位置,且与不同组段的大圆相连。*表示该分类单元存疑图6 华南板块上扬子地区古地理复原图(A)及早泥盆世代表性陆生植物群的生物地理网络图(B,C)Fig.6 Palaeogeography map of the Upper Yangtze in South China Block(A)and biogeographic network graph of the Early Devonian representative floras(B,C)
值得注意的是,在华南板块内部,几个早泥盆世植物群的组成面貌也存在着巨大差别,共有分子较少(图6;表 2,表 3)。以研究最为详细、物种多样性最高的云南坡松冲植物群为例,在属一级上,该植物群与徐家冲植物群仅共有Guangnania、Hedeia、Huia及全球广布属Zosterophyllum,与平驿铺植物群仅共有Adoketophyton、Guangnania、Zosterophyllum及存疑的Psilophyton(图6-B)。在这几个重要的化石产地中,Zosterophyllum是唯一共有的属,这也反映出Zosterophyllum在早泥盆世是优势类群、在全球广泛分布等特性。除了Zosterophyllum,分布较为广泛的植物为Adoketophyton和Guangnania,Adoketophyton在云南坡松冲植物群、四川平驿铺植物群和贵州包阳剖面蟒山群中均有发现,而Guangnania则在云南坡松冲植物群、徐家冲植物群和四川平驿铺植物群中可见到其身影。其他诸如Drepanophycus、Hedeia、Huia也显示出较广的分布范围(图6-B;表 2)。在种一级上,各植物群之间联系则更为稀少,云南坡松冲植物群甚至与四川平驿铺植物群之间毫无共有分子。Z.australianum是少数几种广泛分布在云南坡松冲植物群、徐家冲植物群和蟒山群的物种,显示出Z.australianum在华南的广布性,加之蟒山群中还发现有A.subverticillatum(黄璞等,待刊出),表明蟒山群中的植物组合与坡松冲植物群之间的联系更为紧密。
随着华南早期陆生植物资料的积累,华南各个植物群之间的差异逐渐被学者们注意到,Hao和Xue(2013)认为这些差别主要是由不同的生存环境所致。坡松冲组的含植物化石层被认为属于河道、泛滥平原、近海沼泽等多种环境下的产物,受海洋影响的证据来自于植物化石层中保存的舌形贝、板足鲎等海相动物化石(Hao and Xue,2013),而徐家冲组则呈现出典型的河流沉积特征,广泛发育红层古土壤(Xueetal.,2016)。Edwards等(2016)认为坡松冲植物群、徐家冲植物群和平驿铺植物群之间存在海拔低于3000 m的山脉(图6-A),但此高度不足以阻碍孢子的传播。需要指出的是,Edwards等(2016)引用的古地理复原图值得商榷,目前学术界对古海拔的认识更多的是推测,缺乏扎实的证据。从沉积环境角度来看,蟒山群的沉积环境与坡松冲组较为接近,而与徐家冲组有所区别,这或许可以解释蟒山群中的植物组成与坡松冲植物群的关系可能更为亲密的原因。未来尚需开展更深入的系统古植物学工作,以及结合精时格架下的沉积古地理学工作,以便更好地理解华南早泥盆世植物群的古地理分布格局。
4 结论
文中记述了贵州都匀包阳剖面蟒山群下段的Zosterophyllumaustralianum新材料,这是该种在贵州地区的首次报道。在形态学特征如孢子囊形状、度量等方面,包阳剖面的Z.australianum与云南坡松冲植物群中的同种材料极为相似。结合Z.australianum及最近在蟒山群中发现的Adoketophytonsubverticillatum的时代延限,可将蟒山群下段的时代限定为早泥盆世布拉格期。Z.australianum在蟒山群中的发现为贵州蟒山群植物组合与云南坡松冲植物群之间的联系提供了新的证据。
致谢中国科学院南京地质古生物研究所王怿研究员、黄家园同学对本文提出有益的意见,2位评审专家对本文提出了宝贵的修改意见。中国科学院古脊椎动物与古人类研究所盖志琨研究员鉴定鱼类化石,中国科学院南京地质古生物研究所徐洪河研究员提供文献资料,郭文博士鉴定腕足化石,汤晶晶工程师、张后蕊女士协助拍摄标本。宿州学院资源与土木工程学院张久东同学、桂林理工大学地球科学学院阮伟民同学协助野外工作。在此一并感谢。
