南方大豆皱叶发生时叶片形态变化及其对产量性状的影响
2022-05-26陈文杰陈渊韦清源郭小红汤复跃赵团结梁江
陈文杰 陈渊 韦清源 郭小红 汤复跃 赵团结 梁江


摘要:【目的】研究南方大豆皺叶发生时叶片形态特征变化及皱叶对产量性状的影响,为明确皱叶对大豆叶片形态、农艺性状和产量相关性状的影响程度及揭示大豆皱叶发生机制提供数据支撑。【方法】利用皱叶症近杂合异质系材料GY_C(皱叶)和GY_N(正常),采用随机区组试验设计,研究大豆皱叶症级为4时,叶片形态、叶绿素含量、光合气体交换参数、农艺及产量相关性状的变化。【结果】皱叶发生后叶片主脉薄壁细胞、皱缩处叶片增厚,叶缘处叶片变薄。叶片皱缩处栅栏组织和海绵组织排列不规则,叶缘处出现多层薄壁细胞。皱叶发生时,大豆叶片鲜重和干重、叶柄鲜重和干重、叶柄长和粗、叶片长和宽、叶面积等形态指标均显著减少(P<0.05,下同)。皱叶的叶绿素含量分布不均,但对光合气体交换参数无显著影响(P>0.05)。大豆发生皱叶后成熟期平均提前3.33 d,茎秆变细,抗倒伏性变差,株高、底荚高、主茎节数等农艺性状也有减少趋势,但不同年份有所不同。皱叶导致大豆单株干物质重、单株荚数、单株粒数、单株粒重、百粒重等产量相关性状显著减少, GY_C较GY_N两年平均减产43.11%。皱叶发生时大豆籽粒蛋白含量有降低趋势、油分含量有增高趋势,但不同年份存在差异。【结论】南方大豆皱叶发生时形态上主要因叶缘不能正常伸展所致,皱叶(症级为4)发生后大豆叶面积减少,植株干物质积累变少,生育期缩短、抗倒伏性变差,严重影响单株粒重等产量相关性状。
关键词: 大豆;皱叶;形态;产量
中图分类号: S565.1 文献标志码: A 文章编号:2095-1191(2022)02-0460-09
Changes of leaf morphology during the occurrence of wrinkled leaves in southern soybean and its effects on yield traits
CHEN Wen-jie CHEN Yuan WEI Qing-yuan GUO Xiao-hong TANG Fu-yue ZHAO Tuan-jie LIANG Jiang
(1Cash Crops Research Institute,Guangxi Academy of Agricultural Sciences, Nanning 530007, China;
2Guangxi Crop Genetic Improvement and Biotechnology Laboratory, Nanning 530007, China;
3Nanjing Agricultural University, Nanjing 210095, China)
Abstract:【Objective】To investigate the effects of crinkle leaf on soybean leaf morphology and yield in South China, which could assist to clarify the impact of crinkle leaf on leaf morphology, agronomic characters and yield-related characteristics of soybean and reveal the occurrence mechanism of soybean crinkle leaf in South China. 【Method】A randomized block trial design was used to research effects on leaf morphology, chlorophyll content, photosynthetic gas exchange parameters, agronomic traits, yield and yield-related traits in two crinkle leaf heterozygous lines:GY_C (crinkle leaf line) and GY_N (normal line). 【Result】After the occurrence of crinkle leaf, the parenchyma of the main vein and the wrinkle spaces of the crinkle leaves thickened, but spaces at the leaf edge became thinner. Palisade tissue and spongy tissue were arranged irregularly at the wrinkled spaces and multi-layer parenchyma cells appeared at the leaf margin. With crinkle leaf occurrence, morphological indexes such as the fresh and dry weights of soybean leaves and petioles, petiole length and thickness, leaf length and width, as well as the leaf area decreased significantly (P<0.05 the same below). The distribution of chlorophyll content in wrinkled leaves was uneven, but had no significant effect on photosynthetic gas exchange parameters (P>0.05). With the occurrence of crinkle leaf, soybean matured in 3.33 days on average than those without it. Stems became thinner, with a decrease in lodging resistance and the agronomic characters such as plant height, bottom pod height and the number of main stem nodes also tended to decrease. However, these differences varied in different years of observation. Crinkle leaf resulted in the reduction of yield-related traits such as plant dry weight, pod and seed numbers per plant, seed weight per plant and 100 seeds weight. The average yield of GY_C was 43.11% lower than that of normal leaf material from GY_N in two years. Seed protein content of the crinkle leaf material tended to decrease and the seed oil content tended to increase, but with variation in different years. 【Conclusion】Morphologically, the main cause of soybean leaf wrinkle in South China is that the leaf margin cannot stretch normally. With the occurrence of wrinkled leaf, the soybean leaf area and the accumulation of plant dry matter decreases, with a shortening of the growth period and a decrease in lodging resistance. Crinkle leaf of soybean in South of China seriously affects the yield and yield-related traits such as grain weight.
Key words: soybean; crinkle leaf; morphology; yield
Foundation items: National Natural Science Foundation of China(32060490); Guangxi National Science Foundation(2019GXNSFAA185009); Modern Agricultural Industy Technology System Construction Project(CARS-04-CES30); Science and Technology Development Fund Project of Guangxi Academy of Agricultural Sciences(Guinongke 2021JM67)
0 引言
【研究意义】叶片是植物重要的光合作用器官,可为植物生长发育提供必要的能源物质。作物生长过程中若叶片受到逆境胁迫、病菌和病毒感染、害虫叮食、基因突变等因素影响,会导致叶片发育异常,不利于产量形成。广东、广西、福建和贵州等地在大豆生产中发现一种形态上类似大豆花叶病毒病(SMV)所致的皱叶症,该症状严重时,叶片皱褶不能展开,严重影响大豆的生长。因此,系统开展南方大豆皱叶症对叶片形态、产量及产量相关性状的影响,对科学认识此类型皱叶症及揭示皱叶发生机制具有重要意义。【前人研究进展】皱叶是大豆叶片发育异常的一种表现,皱叶有很多种类,如病毒病(Samertwanich et al.,2001;Campos et al.,2014;王大刚等,2018;Yang et al.,2019)、除草剂(Kelley et al.,2005;Robinson et al.,2013;Silva et al.,2018)、錳毒(Santos et al.,2017)、基因突变(Song et al.,2015;Wang et al.,2016)等因素均可引起皱叶,叶片皱缩直接或间接导致大豆减产。SMV侵染大豆后导致大豆叶片叶绿素含量下降,叶面积减少,植株光合面积和光合能力降低,致使大豆产量损失15%~35%,严重时损失可达70%以上,甚至绝产(刘志涛,2014)。Andersen等(2004)研究表明,56 g/ha的麦草畏处理2节期大豆,叶片皱缩严重,可导致大豆减产83%左右,112 g/ha的2,4-D丁酯处理2节期大豆可导致大豆减产28.7%左右。实际生产中,以前在我国南方等地大豆生产上发现的皱叶症多被误认为由病毒病引起。近些年,本课题组从大豆皱叶的发生特点和发生诱因等方面,对广东、广西、福建和贵州等地大豆发生的皱叶情况进行调查研究,发现南方地区发生的这种类型的皱叶能稳定遗传,且存在基因与环境互作(陈文杰等,2020),环境中的土壤因素是导致南方大豆叶片皱缩的最主要因素。前期利用皱叶和正常叶差异明显的材料衍生的皱叶杂合株系,初步调查了皱叶对大豆产量及其相关性状的影响,发现皱叶严重时(症级为4)可导致大豆减产20%~30%,说明南方大豆发生皱叶对其产量影响较大。【本研究切入点】本课题组前期对南方大豆皱叶现象发生的特点、诱因等进行了初步研究,但目前还缺乏用遗传背景纯合材料进行的皱叶发生对大豆产量及其相关性状影响的研究。【拟解决的关键问题】利用遗传背景较纯合的皱叶症近杂合异质系材料,采用随机区组试验,研究皱叶发生对叶片形态、叶片光合能力、产量相关指标及大豆籽粒品质等的影响,为明确皱叶对大豆叶片形态、农艺性状和产量相关性状的影响程度及揭示大豆皱叶发生机制提供数据支撑。
1 材料与方法
1. 1 试验材料
试验材料为桂春8号(正常叶,皱叶症级为0)和粤春2017-1(皱叶,皱叶症级为4)有性杂交衍生的近杂合异质系材料。创制方法:以桂春8号为母本,粤春2017-1为父本杂交获得F0种子;F0种子播种后根据皱叶性状去除假杂种得到F1单株;F1单株种子株行种植,并调查每个植株的皱叶症症级(陈文杰等,2020),成熟时每行单株收获F2种子;F2单株种子株行种植,并调查每个株行的皱叶症发生情况,株行全皱或全正常的视为纯合株系,同时有皱叶和正常叶的株系视为杂合株系;从杂合株系中选择植株皱叶症级为4的株系(陈文杰等,2020),单株收获皱叶植株获得F2∶3种子;F2:3株系单株株行种植,成熟时选择其中一个杂合株系单株收获皱叶植株获得F2:4种子;F2:4种子株行种植,成熟时选择其中一个杂合株系单株收获皱叶植株获得F2:5种子。按照此方法直至F2:7的株行,从F2:7的株行选择正常叶片株系和纯合皱叶株系混收,获得近杂合异质系材料GY_C(皱叶)和GY_N(正常)(图1)。
1. 2 试验方法
田间试验于2020—2021年在广西农业科学院明阳试验基地进行。2020年播种时间为7月19日,2021年播种时间为7月8日。试验采用单因素随机区组设计,重复3次,小区长5.0 m,宽1.2 m,每小区种植3行大豆,采取穴播方式,行距40 cm,穴距20 cm,播种时每穴播4~6粒,出苗后于1节期每穴定苗2株。播种前施入基肥复合肥(N∶P∶K=18∶18∶18)375 kg/ha,之后不再追肥,田间管理同正常大田。
1. 3 测定项目及方法
1. 3. 1 叶片形态结构观察 外观形态观测:参照Fehr和Caviness(1977)划分的生育时期测定方法,标记GY_C和GY_N盛花期长势相近的生长点幼芽,分别于标记后的第1、2、3、4和5 d采摘拍照观测皱叶和正常叶发育过程中的形态变化。显微镜观测:始粒期选择具有代表性植株倒3叶的中间叶片,于叶基向叶尖方向1/3处的叶脉和叶缘取材后,立即放入FAA标准固定液(Servicebio)固定保存,通过脱水—石蜡包埋—切片—脱蜡—染色等过程制成永久性石蜡切片,采用尼康ECLIPSE Ni型显微镜观察分析(杨虎彪等,2009),并参考戴志冲等(2009)的方法用ImageJ 2016计算叶脉薄壁组织厚度、叶脉基部叶片厚度、叶片皱缩处叶片厚度和叶缘处叶片厚度。每个切片测量4次,测量3个切片。
1. 3. 2 叶片形态指标测定 2021年8月25日,每小区取始粒期10株大豆倒3叶复叶,用直尺测量每片复叶的叶柄长,游标卡尺测量每片复叶的叶柄粗,天平分别称取复叶和叶柄的鲜重。利用HP Laser Jet MFP M227扫描仪对叶片进行扫描,参考戴志冲等(2009)的方法用ImageJ 2016计算各处理叶片的长、宽及叶面积。扫描后取出叶片和叶柄装入信封,置于烘箱108 ℃杀青30 min,80 ℃烘干至恒重,用天平分别称取叶片和叶柄的干重,计算比叶重(叶片干重/叶片面积),重复3次。
1. 3. 3 叶绿素含量测定 2021年8月25日,大豆长至始粒期时,取长势相近的GY_C和GY_N倒3叶,采用丙酮乙醇混合液法测定叶片不同部位的叶绿素a、叶绿素b及总叶绿素含量(张宪政,1986):用剪刀分别取叶片边沿(B)、叶脉到叶缘中间(M)、叶脉基部(V)部位叶片0.2 g,剪成细丝状放入50 mL的加有丙酮乙醇混合液的容量瓶,并定容至50 mL,然后放置黑暗的室温环境浸泡12 h,用WD-9403B型紫外分光光度计测定不同溶液在663和645 nm波长下的光密度值D663和D645。
叶绿素a含量=(12.7D663+2.69D645)×[V/1000×W]
叶绿素b含量=(22.9D645+4.68D663)×[V/1000×W]
叶绿素总含量=(20.2D645+8.02D663)×[V/1000×W]
式中,V为提取液体积(mL),W为叶片鲜重(g)。
1. 3. 4 光合生理指标测定 待大豆长至始粒期,于晴天时8:00—11:00用便携式光合仪LI-6400XT测定无虫咬的倒3叶中间叶的光合气体交换参数净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)(陈文杰等,2017)。光合仪用LED红蓝光源,测定参数:光强1200 μmol/(m2·s),流速500。每小区测4株。参考Fishcher和Turner(1978)的方法计算水分利用率(WUE),WUE =Pn/Tr。
1. 3. 5 农艺性状、产量性状及品质测定 参照邱丽娟和常汝镇(2006)的方法田间调查每小区的花期和始熟期,计算播种至开花天数和播种至成熟天数。以试验小区全部植株为调查对象,计算各处理的倒伏比率,倒伏比率(%)=小区内倒伏植株数(主茎与地面倾斜角度小于30°)/小区内所有大豆植株数×100。始熟期时每小区连续取5株植株地上部分,烘箱108 ℃杀青30 min,80 ℃烘干至恒重,称量各处理的单株干物质重。成熟后连续取10株进行室内考种,记录各小区的茎粗、株高、底荚高、分枝数、主茎节数、单株荚数、单株粒数、单荚粒重、单株粒重、百粒重和容重等指标。小区收获脱粒称重后折算公顷产量,用TPKZ-2型作物考种设备(浙江拓普云农业科技股份有限公司)测量种子的长、宽和籽粒面积(平行子叶),用DA7200近红外谷物分析仪测定各处理的种子蛋白和油分含量。
1. 4 统计分析
利用Excel 2019进行数据整理及表格制作,利用SAS 8.0进行数据的方差分析。
2 结果与分析
2. 1 大豆皱叶形态结构变化
由图2可看出,GY_C材料皱叶发生时,幼小起初叶脉处不表现皱缩,叶尖和靠近葉尖处叶缘有干枯状,随着叶片的生长,叶缘处不能正常伸展致使叶脉不能正常伸长,导致叶脉呈S形生长,叶片沿着叶脉呈波浪形皱褶;无论幼叶或成熟叶片,皱缩发生时均在靠近叶缘处出现黄斑(图2-A)。而CY_N叶片则在幼小初期叶尖正常,随着叶片的生长,叶缘正常展开,最后表现为叶面平整(图2-B)。
图3为显微镜观察结果,与正常叶相比,皱叶发生时叶片皱缩处叶脉薄壁组织增厚(图3-A和图3-B),栅栏组织和海绵组织排列不规则,栅栏组织由垂直表皮排列变平行表皮排列,叶片下表皮细胞变褶皱(图3-C和图3-D)。正常叶叶缘的表皮细胞下即为叶肉,而皱叶的叶缘表皮皮下有较厚的薄壁细胞(图3-E和图3-F)。由表1可知,皱缩叶片的叶脉薄壁组织较正常叶显著增厚202.62%(P<0.05,下同),叶脉基部叶片厚度差异不显著(P>0.05,下同),皱缩处叶片厚度较正常叶显著增厚73.95%,但叶缘处叶片厚度却较正常叶显著减少38.44%。表明皱叶发生时,叶脉变粗,叶片薄厚不均匀,叶缘处变薄,皱缩处叶片增厚。
2. 2 皱叶发生时大豆叶片形态指标的变化
由表2可知,皱叶发生时,大豆叶片鲜重和干重、叶柄鲜重和干重、叶柄长和粗、叶片长和宽、叶面积等形态指标与正常叶相比均显著减少,降幅分别为38.77%、48.57%、43.88%、50.00%、22.55%、20.69%、42.36%、30.36%和59.23%;皱叶较正常叶的比叶重显著增加17.52%。
2. 3 皱叶发生时大豆叶片叶绿素含量及光合指标的变化
由表3可知,皱叶发生时,大豆叶片整体上变得浓绿,但叶绿素含量分布不均匀,叶绿素a和叶绿素b含量均在靠近皱叶叶片边沿处降低,而叶脉到叶缘中间和叶脉基部的叶绿素含量均显著增加,其中叶脉基部叶绿素a和叶绿素b含量分别增加48.10%和60.23%,叶脉到叶缘中间的叶绿素a和叶绿素b含量分别增加23.18%和36.19%。由表4可知,与正常叶相比,皱叶发生时大豆叶片的Pn、Gs、Ci、Tr和WUE均略有升高,但均无显著差异。
2. 4 皱叶发生时大豆农艺性状的变化
由表5可知,皱叶症级为4时,GY_C的播种至开花天数与GY_N无差异,但成熟时间提前,2020和2021年GY_C和GY_N的播种至成熟天数差异显著,两年平均成熟时间提前3.33 d。皱叶后植株倒伏比率显著增加,抗倒伏性变差。皱叶发生后大豆茎粗变细,主茎节数减少,茎粗平均减少19.27%,主茎节数平均减少12.45%。不同年份皱叶对大豆株高影响不同,2020年皱叶导致大豆株高显著降低27.50%,而2021年则无显著变化。皱叶对大豆分枝数无显著影响。
2. 5 皱叶发生时大豆产量性状和籽粒性状的变化
由表6可知,皱叶对大豆的产量相关性状有显著影响。皱叶导致大豆单株干物质重、单株荚数、单株粒数、单株粒重和百粒重减少,两年平均分别显著减少46.22%、28.87%、31.11%、38.78%和11.15%。但皱叶对大豆单荚粒数无显著影响。
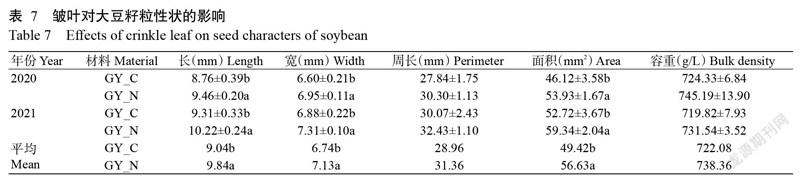
由表7可知,皱叶还影响到籽粒性状,皱叶症级为4时大豆籽粒的长、宽、面积均显著减少,两年平均减少8.13%、5.47%和12.73%。但皱叶对大豆籽粒的周长和容重无显著影响。
2. 6 皱叶发生时大豆产量及品质变化
由表8可知,皱叶对大豆产量有显著影响。2020年皱叶材料较正常叶材料减产40.70%,2021年减产45.00%,两年平均减产43.11%。皱叶还有降低大豆籽粒粗蛋白含量、增加油分含量的趋势,籽粒蛋白含量两年平均降低1.53%,油分含量平均增加0.99%。
3 讨论
利用遗传背景纯合的差异性状材料是研究作物某一性状对产量性状影响的最佳途径之一(刘萍等,2018;郭清云等,2020;韩玉洲等,2021)。本课题组利用皱叶症差异较大的材料有性杂交后,不断分离杂合单株后从F2:7的株行选择正常叶片株系和纯合皱叶株系混收,获得近杂合异质系材料GY_C和GY_N,理论上两者遗传背景纯合率已达99.22%。利用2个材料研究皱叶发生后大豆叶片的形态、光合气体交换参数、农艺性状及产量相关性状的变化,结果较可靠。
植物的叶片发育过程中有多种代谢途径参与(郑梦迪等,2019)。参与叶片发育途径的基因发生突变便有可能发生畸形叶,如拟南芥中参与编码质体包膜蛋白结构的基因CRL发生突变会影响细胞分裂、分化及质体分化,从而导致拟南芥叶片出现皱缩症状(Asano et al.,2004;Chen et al.,2009;?imková et al.,2012)。在大豆上也有相关基因突变所致叶片皱缩的报道(Song et al.,2015;Wang et al.,2016),但在形态上与南方大豆叶片皱缩类型完全不同。此外,外界因素如病毒病(王大刚等,2018;Yang et al.,2019)、除草剂(Kelley et al.,2005;Robinson et al.,2013;Silva et al.,2018;Scholtes et al.,2019)也能导致大豆出现皱叶现象。因病毒引起皱缩的叶片在电镜下可看到“风轮状内含体”等病毒结构(李学湛等,1991;马娜等,2016),但南方大豆皱缩叶片在电镜下观察不到病毒结构;麦草畏等除草剂引起叶片皱缩呈“杯状叶”(Weidenhamer et al.,1989),而南方大豆皱叶叶片多呈“勾手”状(陈文杰等,2020)。此外,本研究通过观察大豆叶片发育过程中叶片皱缩的特点,发现南方大豆皱叶可能是由于叶缘干枯无法正常延伸使得叶脉无法正常伸长所致,同锰毒等其他因素所致的皱叶症状也不同(Santos et al.,2017)。南方大豆叶片皱缩类型形态上不同于已报道的因基因突变、病毒、除草剂所致的皱叶类型,但大豆叶片皱缩形态上的差异也有可能因使用的品种、气候等差异导致。因此,若要探明南方大豆皱叶的遗传及分子机制还需尽快找出导致南方大豆皱叶的诱因。
前人研究表明,光合效率同大豆产量呈显著正相关(Zhu et al.,2002)。本研究发现,大豆皱叶发生时,局部叶片增厚,叶绿素含量增加,但光合速率等光合气体交换参数并未显著提高,可能是由于皱叶发生时大豆叶片处于病态,光合通路中其他代谢过程受到影响所致。作物的经济产量同光合能力、光合叶面积、光合时间呈正相关关系(杜维广等,1999),皱叶导致大豆叶面积减少,生育期缩短,在光合能力不变的情况下,皱叶发生后光合面积和光合时间减少,大豆植株“源”减少,进而导致大豆植株干物质积累量减少,后期表现为大豆单株干重、单株荚数、单株粒数、单株粒重、百粒重等产量相关性状减少,最终导致减产。皱叶癥级为4时,大豆两年试验平均减产43.11%,说明皱叶发生对南方大豆产量影响明显。
不利的外界环境不仅影响植物叶片生长,还会影响根、花等组织的生长。如铝毒(刘鹏等,2004)、镉胁迫(刘晓庆等,2017)、低磷(张文献等,2020)等均会影响大豆根系的正常生长。南方大豆皱叶症是南方土壤中某种致病因子同大豆基因互作导致大豆出现的一种病态表现,干物质积累、产量及产量相关性状受到的影响可能为致病因子诱发大豆叶片产生畸形后而产生的次级影响。本研究主要关注大豆的地上部分,外观上皱叶发生后大豆花、茎秆等组织形态并未发生明显改变,皱叶影响最严重的为大豆叶片。因此,为探明南方大豆皱叶现象的发生机制,后续还需利用遗传背景纯合的皱叶差异材料通过嫁接技术研究皱叶诱因是否对大豆根系生长产生影响。
4 结论
我国南方大豆皱叶发生时在形态上主要因叶缘不能正常伸展所致,皱叶(症级为4)发生后大豆叶面积减少,植株干物质积累变少,生育期缩短、抗倒伏性变差,严重影响单株粒重等产量相关性状,导致大豆显著减产。
参考文献:
陈文杰,陈渊,韦清源,郭小红,汤复跃,叶万典,杨萌,梁江. 2020. 一种大豆皱叶症发生特性及材料症级鉴定[J]. 大豆科学,39(3):431-441. [Chen W J,Chen Y,Wei Q Y,Guo X H,Tang F Y,Ye W D,Yang M,Liang J. 2020. Occurrence characteristics and identification of resistant materials for soybean crinkle leaf disease[J]. Soybean Science,39(3):431-441.] doi:10.11861/j.issn.1000-9841. 2020.03.0431.
陳文杰,郭小红,汤复跃,韦清源,陈渊,梁江. 2017. 不同生育时期大豆叶片光合特性变化及相关性研究[J]. 中国油料作物学报,39(3):360-365. [Chen W J,Guo X H,Tang F Y,Wei Q Y,Chen Y,Liang J. 2017. Changes and relationship of photosynthetic traits of soybean leaves at different growth stages[J]. Chinese Journal of Oil Crop Sciences,39(3):360-365.] doi:10.7505/j.issn.1007-9084. 2017.03.010.
戴志聪,杜道林,司春灿,林英,郝建良,孙凤. 2009. 用扫描仪及Image J软件精确测量叶片形态数量特征的方法[J]. 广西植物,29(3):342-347. [Dai Z C,Du D L,Si C C,Lin Y,Hao J L,Sun F. 2009. A method to exactly measure the morphological quantity of leaf using scanner and Image J software[J]. Guihaia,29(3):342-347.] doi:10. 3969/j.issn.1000-3142.2009.03.013.
杜维广,张桂茹,满为群,栾晓燕,陈怡,谷秀芝. 1999. 大豆光合作用与产量关系的研究[J]. 大豆科学,18(2):61-66. [Du W G,Zhang G R,Man W Q,Luan X Y,Chen Y,Gu X Z. 1999. Study on relationship between soybean photosynthesis and yield[J]. Soybean Science,18(2):61-66.]
郭清云,蒯婕,汪波,刘芳,张椿雨,李根泽,张云云,傅廷栋,周广生. 2020. 感抗油菜近等基因系混播对根肿病发病率的影响[J]. 作物学报,46(9):1408-1415. [Guo Q Y,Kuai J,Wang B,Liu F,Zhang C Y,Li G Z,Zhang Y Y,Fu T D,Zhou G S. 2020. Effect of mixed-sowing of near-isogenic lines on the clubroot disease controlling efficiency in rapeseed[J]. Acta Agronomica Sinica,46(9):1408-1415.] doi:10.3724/SP.J.1006.2020.04074.
韩玉洲,张勇,杨阳,顾正中,吴科,谢全,孔忠新,贾海燕,马正强. 2021. 小麦株高QTL Qph.nau-5B的效应评价[J]. 作物学报,47(6):1188-1196. [Han Y Z,Zhang Y,Yang Y,Gu Z Z,Wu K,Xie Q,Kong Z X,Jia H Y,Ma Z Q. 2021. Effect evaluation of QTL Qph.nau-5B controlling plant height in wheat[J]. Acta Agronomica Sinica,47(6):1188-1196.] doi:10.3724/SP.J.1006.2021.01053.
李学湛,Kim K S,Sctt H A. 1991. 复合病毒感染对大豆叶片细胞超微结构的影响[J]. 电子显微学报,(1):10-15. [Li X Z,Kim K S,Sctt H A. 1991. Ultrastructral affect of compound viruses infection to soybean leaf cells[J]. Journal of Chinese Electron Microscopy Society,(1):10-15.]
刘鹏,Yang Y S,徐根娣,朱申龙. 2004. 铝胁迫对大豆幼苗根系形态和生理特性的影响[J]. 中国油料作物学报,26(4):51-56. [Liu P,Yang Y S,Xu G T,Zhu S L. 2004. The effect of aluminum stress on morphological and physiological characteristics of soybean root of seedling[J]. Chinese Journal of Oil Crop Sciences,26(4):51-56.] doi:10.3321/j.issn:1007-9084.2004.04.010.
刘萍,董文汉,王明君,尹元萍,董蓉娇,张慧,梁泉. 2018. 低磷胁迫条件下大豆磷高效近等基因系主要农艺性状分析[J]. 西南农业学报,31(8):1553-1558. [Liu P,Dong W H,Wang M J,Yin Y P,Dong R J,Zhang H,Liang Q. 2018. Analysis of main agronomic characters of soybean inbred lines with high phosphorus efficient at low phosphorus conditions[J]. Southwest China Journal of Agricultural Sciences,31(8):1553-1558.] doi:10.16213/j.cnki. scjas.2018.8.001.
刘晓庆,陈华涛,张红梅,张智民,陈新. 2017. 不同品种大豆幼苗对镉胁迫的响应[J]. 江西农业学报,29(3):14-17. [Liu X Q,Chen H T,Zhang H M,Zhang Z M,Chen X. 2017. Response of seedlings of different soybean varieties to cadmium stress[J]. Jiangxi Agricultural Science,29(3):14-17.] doi:10.19386/j.cnki.jxnyxb.2017.03.03.
刘志涛. 2014. 植物保健剂对大豆生长和产量以及大豆花叶病毒抗性的影响[D]. 南京:南京农业大学. [Liu Z T. 2014. Influence of plant health agents on soybean growth,yield and the resistance to soybean mosaic virus[D]. Nanjing:Nanjing Agricultural University.]
马娜,栾鹤翔,沈颖超,赵琳,刘志涛,张锴,王成坤,智海剑. 2016. 大豆花叶病毒侵染大豆抗感近等基因系后叶片超微结构变化的比较[J]. 大豆科学,35(2):280-284. [Ma N,Luan H X,Shen Y C,Zhao L,Liu Z T,Zhang K,Wang C K,Zhi H J. 2016. Cellular ultrastructure analysis in soybean resistant lines and susceptible lines of NILs infected by SMV[J]. Soybean Sciences,35(2):280-284.] doi:10.11861/j.issn.1000-9841.2016.02.0280.
邱麗娟,常汝镇. 2006. 大豆种质资源描述规范和数据标准[M]. 北京:中国农业出版社:69-73. [Qiu L J,Chang R Z. 2006. Descriptors and data standard for soybean(Glycine spp.)[M]. Beijing:China Agriculture Press:69-73.]
王大刚,李凯,智海剑. 2018. 大豆抗大豆花叶病毒病基因研究进展[J]. 中国农业科学,51(16):3040-3059. [Wang D G,Li K,Zhi H J. 2018. Progresses of resistance on soybean mosaic virus in soybean[J]. Scientia Agricultura Sinica,51(16):3040-3059.] doi:10.3864/j.issn.0578-1752.2018.16.002.
杨虎彪,李晓霞,罗丽娟. 2009. 植物石蜡制片中透明和脱蜡技术的改良[J]. 植物学报,44(2):230-235. [Yang H B,Li X X,Luo L J. 2009. An improved clearing and de-waxing method for plant paraffin sectioning[J]. Chinese Bulletin of Botany,44(2):230-235.] doi:10.3969/j.issn.1674-3466.2009.02.013.
张文献,李增强,胡亚丽,梁志辰,罗登杰,卢海,唐美琼,陈鹏. 2020. 不同浓度磷胁迫对大豆幼苗生长及根系DNA甲基化水平的影响[J]. 中国农业大学学报,25(12):9-18. [Zhang W X,Li Z Q,Hu Y L,Liang Z C,Luo D J,Lu H,Tang M Q,Chen P. 2020. Effects of different concentrations of phosphorus stresses on soybean(Glycine max L.) seedling growth and DNA methylation in root system[J]. Journal of China Agricultural University,25(12):9-18.] doi:10.11841/j.issn.1007-4333.2020.12.02.
张宪政. 1986. 植物叶绿素含量测定——丙酮乙醇混合液法[J]. 辽宁农业科学,(3):26-38. [Zhang X Z. 1986. Determination of plant chlorophyll content—Acetone ethanol mixture method[J]. Liaoning Agricultural Sciences,(3):26-38.]
郑梦迪,王春阳,张寒,张彦,汪兴军. 2019. 植物叶缘和叶脉发育调控的研究进展[J]. 生物资源,41(1):22-27. [Zheng M D,Wang C Y,Zhang H,Zhang Y,Wang X J. 2019. Advances in plant leaf margin and venation pattern regulation mechanism[J]. Biotic Resources,41(1):22-27.] doi:10.14188/j.ajsh.2019.01.004.
Andersen S M,Clay S A,Wrage L J,Matthees D. 2004. Soybean foliage residues of dicamba and 2,4-D and correlation to application rates and yield[J]. Agronomy Journal,96:750-760. doi:10.2134/agronj2004.0750.
Asano T,Yoshioka Y,Kurei S,Sakamoto W,Machida Y,Sodmergen. 2004. A mutation of the CRUMPLED LEAF gene that encodes a protein localized in the outer envelope membrane of plastids affects the pattern of cell division,cell differentiation,and plastid division in Arabidopsis[J]. Plant Journal,38(3):448-459. doi:10.1111/j.1365-313X. 2004.02057.x.
Campos R E,Bejerman N,Nome C,Laguna I G,Pardina P R. 2014. Bean yellow mosaic virus in soybean from Argentina[J]. Journal of Phytopathology,162(5):322-325. doi:10. 1111/jph.12185.
Chen Y L,Asano T,Fujiwara M T,Yoshida S,Machida Y,Yoshioka Y. 2009. Plant cells without detectable plastids are generated in the crumpled leaf mutant of Arabidopsis thaliana[J]. Plant and Cell Physiology,50(5):956-969. doi:10.1093/pcp/pcp047.
Fehr W R,Caviness C E. 1977. Stages of soybean development. Special report 80,cooperative extension service,agriculture and home economic experiment station[M]. Iowa:Iowa State University:1-11.
Fishcher R A,Turner N C. 1978. Plant productivity in the arid and semiarid zones[J]. Annual Review of Plant Physio-logy,29(1):277-317. doi:10.1146/annurev.pp.29.060 178. 001425.
Kelley K B,Wax L M,Hager A G,Riechers D E. 2005. Soybean response to plant growth regulator herbicides is affected by other post emergence herbicides[J]. Weed Science,53:101-112. doi:10.1614/WS-04-078R.
Robinson A P,Simpson D M ,Johnson W G. 2013. Response of glyphosate tolerant soybean yield components to dicamba exposure[J]. Weed Science,61(4):526-536. doi:10.1614/WS-D-12-00203.1.
Samertwanich K,Kertipakorn K,Chiemsombat P,Ikegami M. 2001. Complete nucleotide sequence and genome organization of soybean crinkle leaf virus[J]. Journal of phtopathology,149:333-336. doi:10.1046/j.1439-0434.2001. 00629.x.
Santos E F,Santini J M K,Paix?o A P,Júnior E F,Lavres J,Campos M,dos Reis A R. 2017. Physiological highlights of manganese toxicity symptoms in soybean plants:Mn toxicity responses[J]. Plant Physiology and Biochemistry,113:6-19. doi:10.1016/j.plaphy. 2017.01.022.
Scholtes A B,Sperry B P,Reynolds D B,Irby J T,Eubank T W,Barber L T,Dodds D M. 2019. Effect of soybean growth stage on sensitivity to sublethal rates of dicamba and 2,4-D[J]. Weed Technology,33(4):555-561. doi:10.1017/wet.2019.39.
Silva E D N,Aguiar A C M,Novello B D P,Silva ? A A,Basso C J. 2018. Rift of 2,4-D and dicamba applied to soybean at vegetative and reproductive growth stage[J]. Ciencia Rural,48(8):1-7. doi:10.1590/0103-8478cr2018 0179.
?imková K,Kim C,Gacek K,Baruah A,Laloi C,Apel K. 2012. The chloroplast division mutant caa33 of Arabidopsis thaliana reveals the crucial impact of chloroplast homeostasis on stress acclimation and retrograde plastid-to-nucleus signaling[J]. The Plant Journal:for Cell and Molecular Biology,69(4):701-712. doi:10.1111/j.1365-313X.2011.0 4825.x.
Song X F,Wei H C,Cheng W,Yang S X,Zhao Y X,Li X,Luo D,Zhang H,Feng X Z. 2015. Development of INDEL markers for genetic mapping based on whole genome resequencing in soybean[J]. Genes Genomes Genetics,5(12):2793-2799. doi:10.1534/g 3.115.022780.
Wang Y Q,Chen W,Zhang Y,Liu M F,Kong J J,Yu Z P,Jaffer A M,Gai J Y,Zhao T J. 2016. Identification of two duplicated loci controlling a disease-like rugose leaf phenotype in soybean[J]. Crop Science,56(4):1611-1618. doi:10.2135/cropsci2015. 09.0580.
Weidenhamer J D,Triplett G B,Sobotka F E. 1989. Dicamba injury to soybeans[J]. Agronomy Journal,81:637-643. doi:10.2134/agronj1989.00021962008100040017x.
Yang X D,Niu L,Zhang W,He H L,Yang J,Xing G J,Guo D Q,Zhao Q Q,Zhong X F,Li H Y,Li Q Y,Dong Y S. 2019. Increased multiple virus resistance in transgenic soybean overexpressing the double-strand RNA-specific ribonuclease gene PAC1[J]. Transgenic Research,28:129-140. doi:10.1007/s11248-018-0108-8.
Zhu G J,Jiang G M,Hao N B,Liu H Q,Kong Z H,Du W G,Man W Q. 2002. Relationship between ecophysiological features and grain yield in different soybean varieties[J]. Acta Botanica Sinica,44(6):725-730. doi:10.1127/0340-269X/2002/0032-0317.
(責任编辑 王 晖)
