两栖蔊菜种群及其入侵群落特征
2022-05-23周凯华吴锁龙柯杰瀚
周凯华,吴锁龙,柯杰瀚,王 艳
(沈阳师范大学生命科学学院, 辽宁 沈阳110034)
据《中国入侵植物名录》[1]记载,截至2013年,我国入侵植物有806种。另外,据闫小玲等[2]对我国外来入侵植物等级的划分,恶性入侵类(一级)有34种,严重入侵类(二级)有69种,局部入侵类(三级)有84种。随着时间的推移,入侵能力强的入侵植物会降低当地植物的适合度、丰度和多样性,破坏生态平衡,甚至导致其他物种灭绝[3],从而造成巨大的经济和生态损失[4]。例如我国北方常出现的三裂叶豚草(Ambrosia trifida)入侵农田等各种生境,造成减产,并对人体健康造成重大威胁[5];光荚含羞草(Mimosa sepiaria)入侵导致原生态系统中特有的动植物丧失[6]。
种群空间分布格局一直是植物种群基本数量特征和种群生态学最活跃的研究领域之一,针对生物入侵植物已开展了很多种群分布格局的研究[7]。作为种群自身的生物学特性,种群的空间分布格局是指种群中的个体在生活空间上的位置状态或布局的状况[8]。种群空间分布格局既是种群适应环境后对空间占有格局的反映,也是对与其他种群相互作用关系的反映[9]。种群分布格局的研究对于了解生物入侵植物种群特征和种间关系有重要意义[10],它既可以定量描述种群的水平结构,也可以揭示植物群落的特征和格局的成因,进而阐明种群和群落的动态变化,从而为制定相应的生态对策提供基础科学依据[11]。入侵植物通常具有较强的适应能力[12],与当地植物竞争时常处于优势,且多呈聚集分布。如有研究发现入侵海南的飞机草(Eupatorium odoratum)在堤埂、林缘和荒地中呈聚集分布[13];入侵云南的肿柄菊(Tithonia diversifolia)在高速公路旁多呈聚集分布[14]。
植物群落的物种组成与结构是群落生态学研究的基础[15-16]。植物群落是指生活在一定区域内所有植物的集合[17],它是植物与其生存的非生物环境和生物环境之间经过长期的相互作用形成的。关于植物群落的研究可以揭示群落特征及分布规律、环境对植物的影响以及植物对环境的适应能力,因此开展关于入侵植物群落特征的研究,对于揭示其入侵能力及机制具有重要意义。据研究非生物因素对入侵植物分布的影响主要表现在大尺度上[18-19],一般随纬度的增加,入侵植物的分布减少[20]。较小尺度上,土壤成分、海拔高度以及水分等都影响植物群落的分布格局。有研究发现随水分的减少[21]、海拔的升高[22],入侵植物的分布逐渐减少。生物因素对入侵植物的分布同样造成一定的影响。生物因素对入侵植物的空间分布的影响主要是在小尺度上显现的[18-19]。随着人类活动对自然环境影响的加剧,植物入侵也随之加剧,其空间分布也随人类活动程度的不同而有差异。在对入侵种牛膝菊(Galinsoga parviflora)的研究[23]中发现,交通运输过程中的无意携带也使得其沿道路集中分布。滇缅公路薇甘菊(Mikania micrantha)危害状况的调查也说明了入侵植物沿着道路方向呈聚集分布[24]。贾桂康[25]研究了环境因子对入侵植物飞机草的影响,经主成分分析发现样地植物总株数、灌木数、林型、空气湿度、水源距离5个因子对飞机草生境生态变量的影响较大,该研究说明了生物因素和非生物因素对于入侵植物具有综合影响。因此,研究群落特征及其与生物与非生物环境因素的关系,对于揭示植物入侵机制具有重要意义。
两栖蔊菜(Rorippa amphibia)为十字花科蔊菜属多年生草本植物,原产欧洲[26],1981年在加拿大出现报道[27]。2003年加拿大学者对来自欧亚大陆的两栖蔊菜采用比例曲线法识别出了其快速且明显入侵的3个时期,分别为1929-1934年、1934-1952年、1971-1979年,但由于当时关于两栖蔊菜的生物学信息很少,因此学者们无法将这3个膨胀阶段与水位波动或任何其他类型的干扰联系起来,最终未解释其可能的入侵机制[28]。我国首次报道在2009年[26],如今该种出现于辽宁省多数地区,主要分布于路边、草坪等生境,据分析它随着草坪草的引入侵入的可能性最大[29]。两栖蔊菜具有很强的营养繁殖和有性繁殖能力,使其能在入侵的草坪等群落中成为优势物种,有爆发性增长的趋势,对入侵地的生态系统有极大破坏力[30]。本研究针对两栖蔊菜经常出现的沈阳部分高校校园草坪绿地开展调查,研究不同生境条件下其种群分布格局等种群特征;采用聚类分析[31-32]和主成分分析(principal components analysis,PCA)[33-34]方法对其入侵的植物群落结构类型及其与环境因子的生态关系进行分析,以揭示两栖蔊菜在入侵不同生境条件下草坪绿地的种群特点和群落特征,探讨影响其分布及入侵能力的主要环境因子,从而在一定程度上揭示其入侵机制,并为控制两栖蔊菜的进一步扩张提供一定的理论基础。
1 研究方法和数据来源
1.1 研究区概况
沈阳(41.8° N、123.4° E)位于辽宁省中部,以平原为主,平均海拔30~50 m,属温带半湿润大陆性气候,全年气温-35~36 ℃,平均气温8.3 ℃,年降水量530~680 mm,全年无霜期183 d。受季风影响,降水集中、温差较大、四季分明。
1.2 研究方法
于2020年9月上旬选取沈阳部分高校(沈阳师范大学、沈阳航空航天大学、沈阳工程学院、辽宁大学蒲河校区)建植3年以上的草坪绿地进行调查,根据绿地所处小环境特点不同大致可分为全遮阴、半遮阴、无遮阴3类生境。采取样方法进行调查,样方面积1 m × 1 m。调查记录内容主要包括样方总盖度、样方中各种植物的高度、分盖度,其中盖度采用目测估计,高度用米尺测量每种植物的冠层的自然平均高度,并记录每个样方中两栖蔊菜植株的数量。共调查73个样方,无遮阴、半遮阴和全遮阴分别采集了13、49、11个样方。同时,用温湿度计(AR 837)、照度计(TES-1339)分别记录各样地距地表20 cm处空气温度、空气湿度和光照强度,并用土钻采集0 -5 cm的土样,在实验室用电热鼓风干燥箱(WG-200)烘干测定土壤含水量。针对上述各项环境因子,均集中选择晴天在09:00 - 10:00测定,连续测定10 d取平均值,而且期间均无降雨影响,以排除随机气候因素引起的土壤水分差异。
1.3 数据分析
1.3.1两栖蔊菜的种群分布格局等种群特征
种群分布格局是指综合利用物种的生物学特性、种间关系和生存环境条件等因素,对其种群个体水平空间配置和分布状态做出的定量描述。种群空间分布格局的测定方法有很多[35-36],为避免不同指数之间存在的片面性,本研究主要采用具有相同本质或较大相似性的7种方法综合分析研究两栖蔊菜的分布格局[37-38],具体如下:
1)扩散系数(diffusion coefficient,C)

若C= 0,属于均匀分布;0 <C< 1,属于规则分布;C= 1,泊松分布(随机分布);C> 1,属于聚集分布。实测与预期的偏离程度用t检验来确定,检测实测值与理论值之间的差距是否显著,计算公式为:

2)负二项参数(negative binomial parameter,K)

式中:K用于度量聚集程度,当K< 8时,为聚集分布(K越小聚集程度越高);当K≥ 8时,为泊松分布。
3)平均拥挤度(mean crowding degree,m*)

式中:m*为生物个体在一个样方中的平均邻居数,反映了样方内生物个体的拥挤程度,数值越大聚集程度越大,表示一个个体受其他个体的拥挤效应越大。
4)丛生指标(cluster index,I)

若I= 0,属于随机分布;I< 0,属于均匀分布;I>0属于聚集分布。
5)聚块性指标(clustering index,PI)

当PI< 1时,为均匀分布;PI= 1时,为随机分布;PI> 1时,为聚集分布。
6) Cassie指标(Cassie index,CA)

当CA< 0时,为均匀分布;CA= 0时,为随机分布;CA> 0时,为聚集分布。
7) Green指数(Green index,GI)

当GI< 0时,为均匀分布;GI= 0时,为随机分布;GI> 0时,为聚集分布。
对不同生境条件下的两栖蔊菜种群数量、高度、盖度等进行差异性统计分析。
利用Excel软件对数据分析和统计,利用SPSS 26.0软件进行单因素方差分析及差异显著性检验,采用Origin 9.0作图。
1.3.2群落分析
利用PAST 4.01对数据进行多元统计分析并作图,包括聚类分析和相关性分析。利用SPSS 26.0对数据进行皮尔逊相关性分析,并用Origin 9.0作图。
2 结果分析
2.1 两栖蔊菜所在不同生境的生态因子特征
调查的两栖蔊菜所在的草坪绿地大致处于3种不同生境类型,分别为无遮阴、半遮阴和全遮阴。无遮阴的生境为无建筑物、树木等遮阴,位于较开阔的大型绿地;半遮阴的生境为位于教学楼等建筑物的东、西侧面,有一定程度遮阴;全遮阴的生境为位于教学楼等建筑物的北面,且绿地边缘有较密集的高大乔木,几乎无直射光。3种生境的照度、湿度、温度等生态因子有较大差别(表1)。无遮阴条件下的照度和空气温度最高且极显著高于全遮阴生境(P< 0.01);而全遮阴条件下的空气湿度和土壤含水量最高且极显著高于其他两种生境下的(P<0.01);半遮阴生境的生态因子均处于中间水平。
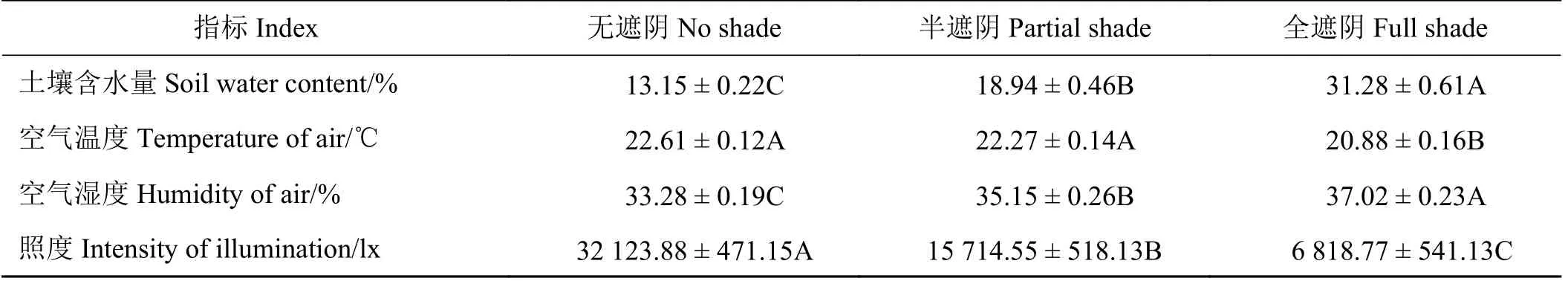
表1 调查各种群生境环境指标Table 1 Environmental indicators of population habitats
2.2 不同生境中两栖蔊菜的空间分布格局
用种群分布格局指标C、K、m*、I、PI、CA、GI判定不同生境类型中两栖蔊菜种群的空间分布格局,上述各种分析方法均判断出在不同生境类型中两栖蔊菜种群为聚集分布(表2)。C和PI均大于1,I、CA和GI均大于0,K为负二项参数,K值越大,表示种群的聚集程度越低,3种生境类型样地中两栖蔊菜种群的K值均远小于8,表明两栖蔊菜种群聚集强度相对较高。针对扩散系数(C)进行t检验的结果达到极显著水平,说明两栖蔊菜种群的分布格局与预期假设呈极显著差异(P< 0.01)。
针对传统的骨盆修复仪所存在的缺陷,本文中提出的骨盆修复仪结构简单、使用方便,且左右壳体调节方便,便于调节使用。可以根据使用者的臀部尺寸自动调节大小,通过连杆的移动带动齿槽的运动,使得齿槽脱离固定齿,实现左壳体与右壳体的移动,且成本较低。同时通过对触摸屏的两侧同时涂抹胶粘剂,能够大大地缩短生产过程所需时间,增加全贴合触摸屏产率,且成本较低。
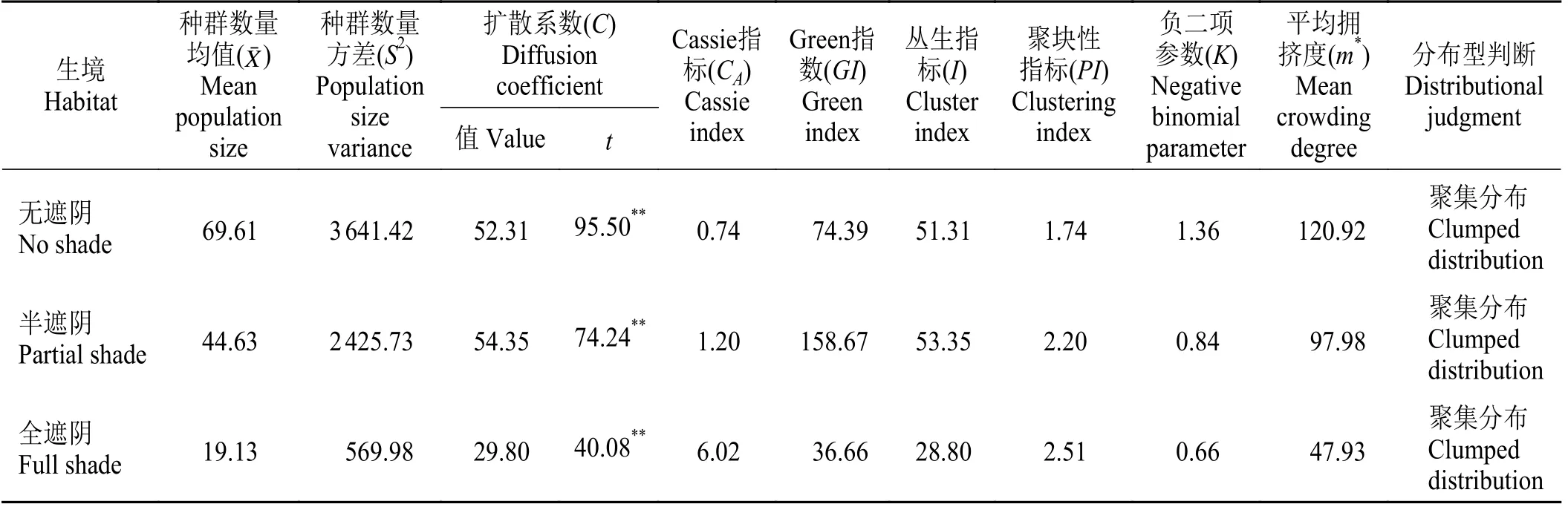
表2 两栖蔊菜的空间分布格局指标计算结果Table 2 Calculation results of spatial distribution pattern index of Rorippa amphibia
2.3 不同生境下两栖蔊菜的种群特征
两栖蔊菜在无遮阴、半遮阴和全遮阴条件下的密度分别为17.41、16.23和5.94株·m-2;高度分别为5.87、8.88 和3.74 cm;盖度分别为25.34%、26.86%和5.60%。这说明在全遮阴条件下两栖蔊菜种群的密度、高度、盖度均最低。在无遮阴和半遮阴条件下,两栖蔊菜的种群密度分别为全遮阴条件下的2.93倍和2.73倍,盖度分别为4.52倍和4.79倍,高度分别为1.57倍和2.37倍。种群密度全遮阴与无遮阴生境条件间差异显著(P< 0.05),但在无遮阴与半遮阴生境之间、半遮阴与全遮阴生境之间无显著差异(P> 0.05) (图1)。不同生境条件下两栖蔊菜的平均高度出现显著差异。全遮阴条件下的种群盖度与无遮阴、半遮阴条件下差异显著,而无遮阴和半遮阴条件下的种群盖度差异不显著。

图1 不同生境类型中两栖蔊菜的种群特征Figure 1 Population characteristics of Rorippa amphibia in different habitat types
2.4 两栖蔊菜所在群落的聚类分析与排序
2.4.1群落的聚类分析
当生物入侵物种侵入到新环境时,入侵种与本地种间的竞争能力也是影响植物入侵的一个重要因素,因此划分入侵植物所在群落的类型,对于揭示该入侵植物在群落中所处的地位、认识入侵种与群落中其他植物的生态关系具有重要意义。
聚类分析在欧氏距离7 500处将所有样方分为3类(图2),3个群落类型及其群落物种组成特征如下:

图2 两栖蔊菜所在群落的聚类分析Figure 2 Clustering analysis of communities of Rorippa amphibia
1)两栖蔊菜 + 草地早熟禾(Poa pratensis)群丛。该群丛所处生境为无遮阴生境类型。样方总盖度平均为55.79%。该类型以两栖蔊菜和草地早熟禾为优势种,其中两栖蔊菜的平均高度5.87 cm,平均盖度25.34%;草地早熟禾的平均高度8.82 cm,平均盖度9.09%。主要伴生种有车前(Plantago asiatica),平均高度1.2 cm,平均盖度5.09%;长裂苦苣菜(Sonchus brachyotus)平均高度3.25 cm,平均盖度5.61%。其他植物有紫花地丁(Viola philippica)、地锦大戟(Euphorbia humifusa)、蒲公英(Taraxacum mongolicum)、狗尾草(Setaria viridis)、牛筋草(Eleusine indica)、抱茎苦荬菜(Xeridium sonchifolium)、铁 苋 菜(Acalypha australis)、东北蒲公英(Taraxacum ohwianum)、柳叶旋覆花(Inulasalicina)、东北油柑(Phyllanthus ussuriensis)等。
2)两栖蔊菜 + 水田碎米荠(Cardamine lyrate)群丛。该群丛为半遮阴条件下的群落类型。群落总盖度平均为58.88%。以两栖蔊菜和水田碎米荠为优势种,其中两栖蔊菜的平均高度9.78 cm,平均盖度26.86%;水田碎米荠平均高度4.22 cm,平均盖度8.47%,该植物多生在湿度比较大的地方,反映出了该群落类型所处的环境比较阴湿;其他主要伴生种有附地菜(Trigonotis peduncularis),平均高度2.87 cm,平均盖度约7.9%;紫花地丁平均高度3.36 cm,平均盖度3.8%;原本为人工建植的优势植物草地早熟禾已经退化,其平均盖度仅为3.48%,但在群落中其平均高度最高,为12.14 cm;地面苔藓植物较发达,盖度32.78%;其他植物种类有白车轴草(Trifolium repens)、狗尾草、马唐(Digitaria sanguinalis)、朝天委陵菜(Potentilla supina)、牛膝菊、车前、东北蒲公英、小飞蓬(Conyza canadensis)、马 齿 苋(Portulaca oleracea)、小藜(Chenopodium serotinum)等。
3)阴地堇菜(Viola yezoensis) + 草地早熟禾群丛。该群丛类型分布于全遮阴条件下。该群落类型以阴地堇菜和草地早熟禾为优势种。群落总盖度约89.08%,其中阴地堇菜的平均高度5.01 cm,平均盖度7.58%;草地早熟禾的平均高度8.32 cm,平均盖度7.49%。与上述两种植物相比,两栖蔊菜的优势度已经不明显,其平均高度3.97 cm,平均盖度5.6%;水田碎米荠平均高度0.74 cm,平均盖度1.53%。其他植物有紫花地丁、车前、铁苋菜、狗尾草、东北蒲公英、附地菜、牛膝菊、马唐、白车轴草、鸭跖草(Commelina communis)、狼尾珍珠菜(Lysimachia barystachys)、繁缕(Stellaria media)等。值得注意的是,在地表覆盖的苔藓植物非常发达,盖度达66.12%,充分体现了环境条件的阴湿特征。
2.4.2群落的PCA排序
群落的排序分析能客观地揭示植被类型和分布与环境因子之间的生态关系,已经成为认识植被分布格局的重要手段。
PCA分析结果(图3)显示,所有调查样方沿着第1主分量和第2主分量分成了3组,反映了不同群落类型所处的环境梯度。主分量分析表明这两个主成分占了总信息量的89.43%,其中第1主分量贡献率为75.91%,该主分量上起主要作用的是土壤含水量,其次是照度;第2主分量贡献率为13.52%,在该分量上起主要作用的是大气温度和湿度(表3)。第1主分量贡献率为第2主分量的5.61倍,因此在第1主分量上起主要作用的环境因素,即土壤含水量和照度对于群落的排序起最主要作用。经过对环境因子指标进行相关性分析,发现虽然各生态因子间都有较显著的关联(图4和表4),如照度与大气温度极显著正相关(P< 0.01),土壤含水量与大气湿度极显著正相关(P< 0.01)等,但土壤含水量和照度这两个生态因子间的相关性最大(-0.903),也说明了二者的综合作用对于两栖蔊菜入侵的群落类型和分布起到了最重要的作用。

表4 环境指标间的皮尔逊相关性分析Table 4 Pearson correlation analysis among environmental indicators
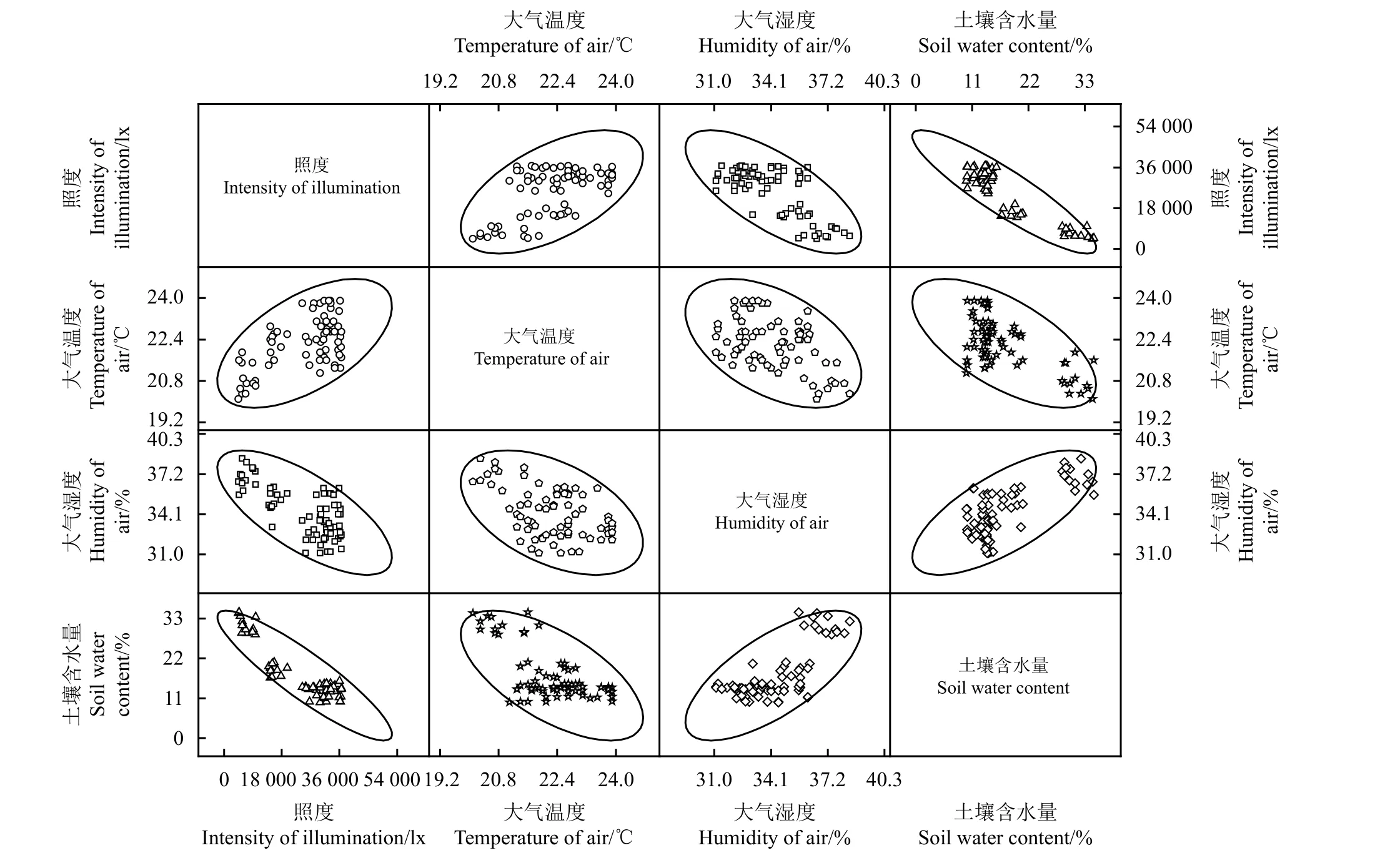
图4 环境指标间的相关性分析Figure 4 Correlation analysis among environmental indicators

表3 主分量的负荷量表Table 3 Load scale of main components

图3 两栖蔊菜所在群落的PCA排序Figure 3 PCA sequencing f communities of Rorippa amphibia
3 讨论与结论
3.1 两栖蔊菜的种群空间分布格局
本研究中两栖蔊菜在3种生境条件下均呈现聚集分布格局,这与甘肃敦煌西湖荒漠湿地生态系统优势植物种群分布格局的研究结果[10]相近。两栖蔊菜聚集分布的原因在于它的繁殖方式,当两栖蔊菜以种子萌发、定植入侵后,可以通过两种方式扩展种群数量,一是通过根状茎进行营养繁殖,二是其种子在成熟之后可能较多地散落在母株的周围,最终导致了两栖蔊菜聚集分布的空间分布格局。另外本研究还发现无遮阴及半遮阴生境中两栖蔊菜的聚集程度较高,全遮阴生境中的聚集程度较低,可能是因为生境条件的不同影响了其营养繁殖和有性繁殖能力。
3.2 环境因素对两栖蔊菜种群特征的影响
有研究表明,阳生外来入侵植物对光的响应更强,光辐射强度增加能提高光合速率、光合资源利用效率,促进其入侵[39],如入侵植物紫茎泽兰(Ageratina adenophora)在适宜的高光环境下能更好地利用环境资源,加快生长[40]。在本研究调查的3类生境中,两栖蔊菜不是在无遮阴环境中而是在半遮阴的环境中生长最好,表现为盖度和高度最大,密度虽然比无遮阴条件下略低但统计学上无显著差别,这可能与半遮阴环境中的空气湿度和土壤含水量较为适宜有关,因为两栖蔊菜具有喜湿的特性[26]。而在全遮阴的环境中,虽然环境湿度大,但由于光照强度过低,不利于其进行光合作用,进而导致其在有机物积累和种群的无性和有性繁殖方面受到影响。因此种群密度、盖度和植株高度较低,说明光照、湿度等环境因子对两栖蔊菜的种群有综合且重要的影响。
3.3 环境因素对两栖蔊菜所入侵群落的影响
本研究运用聚类分析的方法将所调查的两栖蔊菜入侵的草坪植物群落分为3个群丛类型,与所调查时简单区分的生境分类相同,充分反映出环境因素的变化对植物群落类型的影响,这与对老师傅海湿地的研究[41]和程莅登等[42]的研究结果相似。
对大青山小井沟自然植物群落的主成分分析研究[43]发现海拔和坡向这两个非生物因素是影响植物群落分布的主要因素。本研究运用PCA方法,对土壤含水量、湿度、温度及照度等环境因子对两栖蔊菜所处群落进行排序,结果表明照度和空气湿度在植物群落分布中起到关键的作用。在半遮阴和无遮阴条件下,两栖蔊菜均能入侵并成为草坪绿地中的优势种。尽管不同生境中两栖蔊菜在群落中的地位会出现一定的变化,但其能入侵不同环境中的草坪群落,并对群落结构产生重要影响。
本研究发现,虽然两栖蔊菜在全遮阴条件下的盖度、高度、密度均显著下降,但其在差别甚大的不同生境中都有出现,且在无遮阴、半遮阴环境条件下均成为群落的优势种,表明它对于水分、温度、光照等环境因子变化的适应能力强,这进而表明其具有较强的适应和入侵能力。将聚类分析结果与PCA排序对比分析可以看出,聚类分析所得各群丛类型在二维排序图上各自有比较明确的分布范围,基本上反映了群丛随环境梯度分布的变化趋势。
余海滨等[44]对乌斯吐自然保护区草本群落进行研究发现优势物种对群落分布起主导作用,本研究划分出的3个群丛类型中有2个两栖蔊菜都已成为了优势物种,说明了两栖蔊菜对入侵群落的影响十分严重。另外,两栖蔊菜在各种条件下的草地早熟禾草坪绿地上入侵现象较为普遍,说明其生物入侵机制与草坪植物引入和种植之间存在一定的正关联,即两栖蔊菜入侵可能是随草坪草引入的[18,23],这可归结为生物因素对该入侵植物的影响。综上,应进一步加强对两栖蔊菜的相关研究,并针对草坪的引入、管理方面加强对两栖蔊菜的防控,遏制其对各地不同生态系统的入侵。
