降植烷通过内质网应激诱导大鼠巨噬细胞自噬
2022-05-21朱文华孟列素
朱文华,韩 燕,宁 启 兰,张 富 军,孟列素,吕 社 民
(1. 西安交通大学基础医学院生物化学与分子生物学系,陕西西安 710061;2. 西安交通大学分子转化医学研究所,陕西西安 710061;3. 环境与疾病相关基因教育部重点实验室,陕西西安 710061)
佐剂是疫苗的重要组成部分,发挥免疫调节剂和抗原传递载体的作用。它可以提高抗原的免疫原性,增强免疫反应,对于疫苗、药物的研发具有重要意义[1]。已发现部分油脂如不完全弗氏佐剂、角鲨烯、矿物油及其组分具有佐剂性质[2-3],但由于其相对较高的细胞毒性、降解效率低等缺点,目前主要应用于动物体内。近年来,MF59 和AS03 这两种含角鲨烯的油性佐剂成功应用于人用流感病毒疫苗,尤其在H1N1 流感暴发时发挥了重要作用,使油性佐剂重新受到了免疫学家的关注[4-5]。大多数的油性佐剂都可在动物体内诱发自身免疫病[3],如不完全弗氏佐剂、角鲨烯、矿物油、降植烷(pristane)等在大鼠中具有致关节炎的能力,但原因不明[6-9]。虽然目前尚缺少致关节炎油剂与类风湿关节炎(rheumatoid arthritis,RA)病因关系的流行病学调查,但这些油性佐剂的广泛存在及其作为佐剂、关节炎诱导剂和免疫调节剂的有效作用,都提示其可能与RA 的发病密切相关。因此,揭示油性佐剂免疫增强作用的机制,不仅有助于了解其佐剂作用,而且有助于阐明关节炎等自身免疫病的发病机制。
自噬是细胞在外源、内源刺激下维持内环境稳态的重要方式,也具有免疫调控作用。本课题组在降植烷诱导的大鼠关节炎模型中发现降植烷能够诱导巨噬细胞自噬,通过自噬-信号转导与转录激活子1(signal transducer and activator of transcription 1,STAT1)-Toll 样受体3(toll-like receptor, TLR3)调控轴介导关节炎症的发生发展[10-11],然而,自噬诱导的具体机制仍不清楚。已有研究发现能量代谢途径、内质网应激等因素参与了自噬的诱导[12]。其中能量代谢的传感分子腺苷酸活化蛋白激酶(AMPK)和哺乳动物雷帕霉素靶蛋白(mTOR)可激活或抑制细胞自噬[13],而内质网应激相关的真核起始因子2α(eIF2α)、肌醇依赖酶1α(IRE1α)等通路也同自噬的发生密切相关[14]。但仍然未知油性佐剂是否能影响这些通路从而调控自噬的发生。因此,本研究在降植烷刺激的大鼠巨噬细胞系中,探讨能量代谢感应器AMPK、mTOR及内质网应激通路的调控情况及其在自噬诱导中的作用,以明确油性佐剂诱导自噬增强的确切分子机制。
1 材料与方法
1.1 细胞培养和降植烷刺激的模型构建
大鼠巨噬细胞系NR8383 购自中科院上海细胞库,用含150 mL/L 胎牛血清(HyClone)的F-12K 培养基(Sigma-Aldrich)培养。
降植烷刺激的巨噬细胞模型构建:将细胞以2.5×105/mL 接种于6 孔板中,过夜使细胞贴壁,将降植烷(ACROS)与完全培养基以1∶1 的比例混合,并通过注射器反复抽吸以制备乳浊液,然后用培养基稀释至工作浓度(1 mmol/L),用新制的油乳剂代替培养基刺激细胞构建模型。根据实验需求于0、0.5、1、3、6 h各时间点收集细胞并抽提蛋白质进行相关检测。
1.2 Western blotting 检测降植烷刺激模型中能量代谢感应器及内质网应激通路蛋白表达
按照1.1 项构建降植烷刺激的模型,分别于刺激后0、0.5、1、3 h 收集细胞,用含蛋白酶和磷酸酶抑制剂(Roche)的RIPA 溶液(Beyotime)抽提总蛋白,采用BCA 试剂盒(Thermo Scientific)测定每个样品的蛋白浓度。以每个样本20 μg 总蛋白,按照Bio-Rad电泳系统标准程序进行SDS-PAGE 凝胶电泳和转膜。一抗分别包括兔抗p-AMPK 抗体(1∶1 000,CST,#2535),兔抗AMPK抗体(1∶1 000,CST,#2532),兔 抗p-mTOR 抗 体(1∶1 000,CST,#2971),兔 抗mTOR 抗体(1∶1 000,CST,#2972),兔抗p-IRE1α 抗体(1 μg/mL,Novus),兔抗-IRE1α 抗体(1 μg/mL,Abcam),兔抗p-eIF2α 抗体(1∶1 000,CST,#3398),兔抗eIF2α 抗体(1∶1 000,CST,#5324)和兔抗-β-肌动蛋白(β-actin)抗体(1∶1 000,CST,#4970),二抗为HRP 耦联的羊抗兔IgG(0.4 μg/mL, Abcam)。
经免疫印迹显色试剂盒Supersignal®West Pico Kit(Thermo Scientific)曝光显色后,采用化学发光成像仪(Syngene)照相并进行条带灰度扫描。采用凝胶成像分析软件Syngene 获得Western blotting 条带的灰度值,将第一泳道(对照组)的灰度值设为1,其他泳道的相对表达值通过灰度值倍数计算获得。
1.3 Western blotting 检测内质网应激细胞干预时自噬蛋白的表达
在降植烷乳液刺激细胞模型中,分别给予内质网应激的诱导剂二硫苏糖醇(DTT,Sigma)(1、5 mmol/L)或抑制剂4-苯基丁酸(4-PBA,Santa Cruz Biotech)(1、10、20 mmol/L)干预,作用6 h,收集细胞并按照1.2 项中Western blotting 方法检测细胞mTOR 和自噬分子微管相关蛋白1-轻链3(microtubule associated protein 1 light chain 3, LC3)的表达。一抗分别包括兔抗mTOR 抗体(1∶1 000,CST,#2972)、兔抗LC3 抗体(1∶1 000,CST,#4108)和兔抗-β-actin 抗体(1∶1 000,CST,#4970)。
1.4 转录因子TFEB 表达与活化检测
降植烷刺激的巨噬细胞模型经降植烷乳液作用6 h,收集细胞并用Beyotime 核质蛋白提取试剂盒分离胞质蛋白和核蛋白,按照1.2 项中Western blotting方法检测TFEB 在胞质和细胞核中的表达,以反映TFEB 的入核情况;以Western blotting 检测管家蛋白β-actin和层粘连蛋白A(Lamin A)的表达,以鉴定胞质蛋白和核蛋白的纯度。一抗分别包括兔抗TFEB 抗体(1∶1 000,CST,#8478)、兔抗-β-actin 抗体(1∶1 000,CST,#4970)、兔抗-Lamin A 抗体(5 μg/mL,BIOSS)。
1.5 统计学分析
细胞实验重复3 次,实验结果以均数±标准误表示。采用统计软件GraphPad Prism 进行差异分析。两组间均数比较采用t检验;多组间均数比较使用One way-ANOVA 方差分析,组间两两比较采用Tukey 检验。以P<0.05 为差异有统计学意义。
2 结 果
2.1 降植烷刺激细胞模型中能量代谢感应器及内质网应激通路蛋白的表达情况
Western blotting 检测降植烷刺激细胞的早期阶段(0、0.5、1、3 h)时,能量代谢感应器AMPK、mTOR及内质网应激通路eIF2α 和IRE1α 的表达变化,结果显示,降植烷刺激时,mTOR 表达显示出下调趋势,尤其在刺激后1、3 h 显著下调(P<0.05),这提示mTOR 是通过总量下调而非改变磷酸化比率发挥作用(图1A、图1B)。与之不同的是,AMPK 无论总量还是磷酸化水平都没有明显改变,提示其可能不受降植烷刺激的影响(图1A);降植烷刺激细胞0.5 h 时,内质网应激通路eIF2α 的磷酸化水平显著增强(P<0.05),在之后的1、3 h 迅速回落,IRE1α 的总量和磷酸化水平则无明显改变(图1A、图1C)。这提示eIF2α 介导的内质网应激通路在降植烷刺激后瞬时激活,且这一过程早于mTOR 通路的改变,其可能通过抑制细胞mTOR 的水平而实现自噬增强。
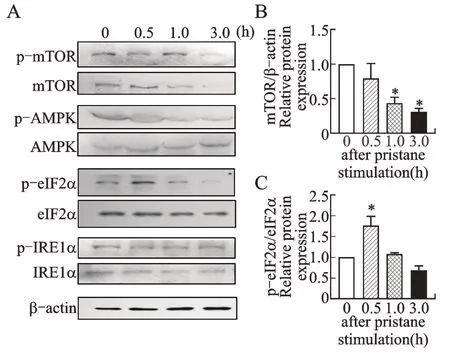
图1 降植烷刺激NR8383 细胞系不同时间点的能量代谢感应器及内质网应激通路蛋白检测Fig. 1 Detection of energy metabolism sensors and ER stress pathways in NR8383 cell line stimulated by pristane at different time points
2.2 干预内质网应激通路对降植烷诱导的巨噬细胞自噬的影响
NR8383 细胞单纯给予1 mmol/L 降植烷刺激(模型细胞)时,mTOR 总量减少(P<0.01),同时LC3-Ⅱ表达增高(即自噬增强,P<0.001);当联合降植烷和内质网应激通路抑制剂4-PBA 刺激时,细胞mTOR 表达回升而LC3-Ⅱ表达减弱,尤其在20 mmol/L 4-PBA 干预组,mTOR 表达显著升高(P<0.05)而LC3-Ⅱ表达显著降低(P<0.01,图2)。由此可见,抑制巨噬细胞内质网应激会抑制mTOR的减少,从而阻碍细胞的自噬增强现象。
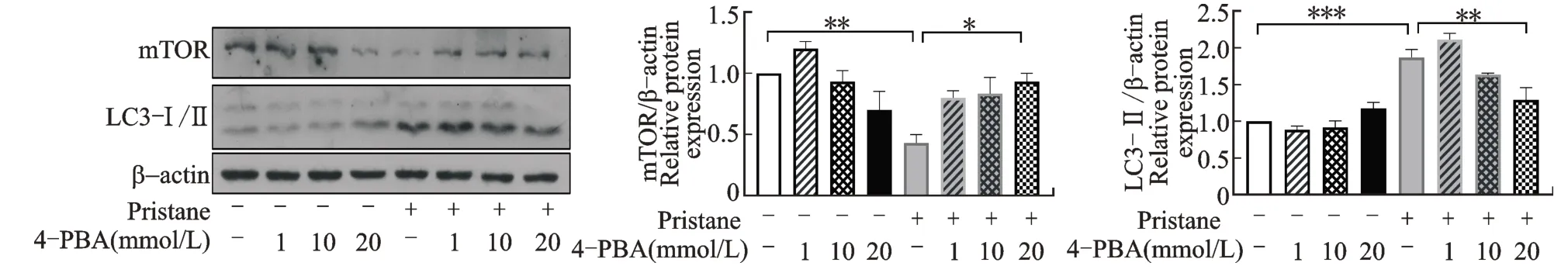
与空白对照组(pristane、4-PBA 未刺激组)比较,*P<0.05,**P<0.01,***P<0.001。
NR8383 细胞单纯给予1 mmol/L 降植烷刺激时,mTOR 总量减少(P<0.01)同时LC3-Ⅱ表达增高(P<0.05);当联合降植烷和内质网应激诱导剂DTT 刺激细胞时,LC3-Ⅱ表达进一步增强,尤其在5 mmol/L DTT 干预组中LC3-Ⅱ表达较单纯降植烷组显著升高(P<0.001,图3)。在DTT 干预组并未出现mTOR 表达进一步降低,这可能是因为降植烷刺激下mTOR 本身已维持较低表达。以上提示促进巨噬细胞内质网应激会进一步增强细胞的自噬水平,降植烷通过诱导巨噬细胞内质网应激降低细胞mTOR 水平,从而实现自噬增强。
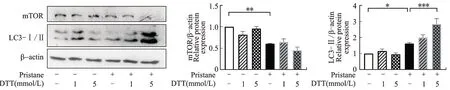
图3 内质网应激激动剂DTT 对降植烷刺激的NR8383 细胞系中mTOR 和自噬水平的影响Fig.3 Effect of ER stress inducer DTT on mTOR and autophagy in pristane stimulated NR8383 cell line
2.3 降植烷刺激的巨噬细胞中TFEB 的表达与活化情况
Western blotting 检测降植烷刺激NR8383 细胞的胞质蛋白和核蛋白中的TFEB 的表达情况,结果显示,胞质蛋白表达β-actin 而不表达Lamin A,而核蛋白表达Lamin A 且未检测到β-actin,提示核、质蛋白分离较好、纯度较高;在降植烷刺激下,TFEB 在胞质蛋白和核蛋白中表达都明显增强,尤其在核蛋白中;降植烷刺激组的TFEB 蛋白表达较对照组显著升高(P<0.01,图4),提示降植烷刺激时TFEB 的表达升高且活化水平增强,可能参与了自噬相关基因的表达调控,这一发现是对降植烷自噬增强机制的重要补充。
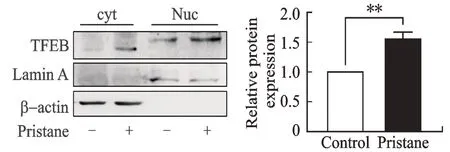
图4 Western blotting 检测降植烷刺激NR8383 细胞系中转录因子TFEB 的表达与活化Fig. 4 Translocation of transcriptional factor TFEB in pristane stimulated NR8383 cell line detcted by Western blotting
3 讨 论
自噬通过清除、降解胞质组分维持细胞内环境稳态,是真核细胞处理内部成分的过程。近年来已发现,自噬广泛参与机体免疫应答调控,其对天然免疫和获得性免疫都有重要调控作用,具有效应者和调控者双重身份。目前已有众多研究证实自噬参与RA的发病和发生发展过程。本课题组团队率先在RA患者滑膜组织中发现增强的自噬现象,自噬抵抗了滑膜细胞的凋亡[15];之后自噬在滑膜组织中的双重作用被进一步全面揭示,研究发现,自噬可促进滑膜细胞中内质网应激诱导的凋亡,并抑制蛋白酶体介导的细胞凋亡[16]。除滑膜细胞以外,本课题组亦通过关节炎的动物模型研究了自噬在免疫细胞中的作用,发现降植烷诱导的关节炎大鼠脾脏巨噬细胞的自噬水平升高,增强的自噬通过STAT1-IRF1 通路介导了TLR3 的表达升高,在体干预时自噬或TLR3 都能显著抑制疾病的严重程度[11],这确证了自噬-TLR3 调控轴在关节炎发病机制中的重要作用,也通过自噬的诱导回答了油性佐剂的免疫增强机制,为理解油性佐剂的佐剂性和致关节炎性提供了理论和实验基础。然而,降植烷诱导巨噬细胞自噬增强的确切机制仍不清楚。
研究发现,降植烷可引起大鼠淋巴细胞膜流动性的改变[17],推测其可能影响包括代谢转运在内的细胞转运系统。而能量代谢的传感分子AMPK 和mTOR可与UNC-51样激酶1(UNC-51-like kinase 1,ULK1)竞争性结合,激活或抑制细胞的自噬水平[13]。因此,本研究在降植烷刺激大鼠巨噬细胞的早期检测了AMPK 和mTOR 通路的表达情况,结果显示,mTOR而非AMPK 受到了降植烷的调控,即mTOR 总量的降低实现了巨噬细胞自噬的增强。另外,也有报道内质网应激可诱导自噬发生[14]。内质网富含多种酶类,是外源物质在细胞内被催化的主要场所。降植烷在细胞内将经历末端羟基化或末端氧化,然后通过经典的β-氧化过程被彻底代谢。因此,推测巨噬细胞过度摄取降植烷可能导致内质网功能障碍,引发内质网应 激。eIF2α 和IRE1α 通 路 是 内 质 网 应 激 的 重 要 通路,已有研究报道它们可参与自噬调节[18-20]。本研究在降植烷刺激的细胞模型中也检测了eIF2α 和IRE1α通路蛋白的表达情况,结果显示eIF2α 在降植烷刺激后瞬时激活,且这个过程早于mTOR 的表达下调,提示降植烷诱导了巨噬细胞的内质网应激从而降低了mTOR 的表达,最终诱导自噬。这一猜测也进一步通过了内质网应激抑制剂和激动剂干预实验得到了证实,但内质网应激调控mTOR 表达的机制仍有待深入解析。
此外,研究发现mTOR 除竞争性结合ULK1 干预自噬外,还可作用于转录因子TFEB,mTOR 失活使其活化并转运至细胞核内,从而控制自噬体和溶酶体相关基因的转录调控[21]。本课题组之前的研究也发现,降植烷刺激的大鼠脾脏组织及巨噬细胞中自噬基因(如Becn1、LC3 等)的表达受到调控[11]。因此,本研究检测了降植烷刺激的巨噬细胞模型中TFEB 的易位入核情况,发现其在刺激时表达增强,且活化增强。结果提示mTOR 有可能通过影响TFEB 的表达与活化调控了自噬的增强,但这仍需要进一步实验验证。
综上,本研究证实了降植烷为代表的油性佐剂通过诱导内质网应激、抑制mTOR 的水平、活化转录因子TFEB,实现了巨噬细胞自噬增强。本研究明晰了油性佐剂的自噬增强机制,并对其致关节炎性和佐剂性的作用机制有了更全面的认识。
