纳米银胁迫对汤氏纺锤水蚤的生殖及基因毒性效应研究
2022-05-20刘星辰吕泓玥金扬湖
刘星辰,吕泓玥,金扬湖,周 超
(浙江海洋大学海洋科学与技术学院,国家海洋设施养殖工程技术研究中心,浙江舟山 316022)
1980 年代中期人造纳米技术发展迅速,纳米材料被广泛应用于电气,医学和材料领域[1]。近年来纳米工艺愈加完善,纳米消费品使用量日益增加,天然及人工合成的纳米材料不可避免地被排入海洋、湖泊等环境,成为亟待解决的环境污染问题之一。纳米银颗粒作为已被广泛应用的纳米颗粒之一,于微电子领域及医用消毒领域占据重要地位,每年天然开采以及人工合成的数量都在上升[2-4]。早有研究表明,纳米银颗粒于天然水中释放银离子,且观察到不同大小和形状的纳米银胁迫导致天然水域生物受到毒性作用[5]。已证明纳米银颗粒对大多数生物组织和生物体有毒性作用,例如细菌、人类、哺乳动物及藻类等[6-8],所以研究纳米银对海洋生态系统毒性作用机制已迫在眉睫。
桡足类作为海洋种群数量最大的类群之一,凭借对各类污染物高度敏感性,常作为优异海洋污染物监测的模式生物[9-10]。水蚤作为最早人工养殖的滤食性桡足类,广布于温带及亚热带地区,与轮虫、卤虫同为经济崽鱼的开口饵料,占据海洋生物食物链的特殊地位,对海洋生态系统物质循环及污染监控与防范具有重大意义[11]。本研究样品为汤氏纺锤水蚤Acartia tonsa,个体尺寸小,成幼体形态结构差异大,对外界刺激敏感,实验室内可大规模培养,故可作海洋生态毒理学模式生物,其毒理学相关研究对海洋海域生态指标评定有重要指示作用[12]。
本文通过研究不同环境浓度暴露下纳米银对汤氏纺锤水蚤生殖力、酶活力水平及抗氧化相关基因表达水平的影响,探究纳米银暴露时间增加是否会导致其颗粒于水蚤体内富集,探究汤氏纺锤水蚤对纳米银刺激相关应激效应机制,为海洋纳米银污染监控及治理奠定基础,为桡足类生态毒理学评估提供科学依据。
1 材料与方法
1.1 微藻饵料培养
培养汤氏纺锤水蚤所用微藻饵料为球等鞭金藻Isochrysis galbana Parke(Iso)、波海红胞藻Rhodmonas baltica Karsten(Rhodo)及Rhinomonas reticulate Lucas(Rhino)3 种海水微藻。微藻藻种皆由意大利环境保护所提供,于实验室恒温培养箱内世代培养。恒温培养箱培养条件:温度(20±0.5)℃,海水盐度30,光照条件(12 h光照:12 h 黑暗),光照强度为2 000 lx。每日监测微藻密度,微藻生长至指数生长期时及时补充海水F/2 培养基[13]。微藻藻种每半个月保种1 次,将微藻无菌接种至含F/2 培养基的50 mL 细胞培养瓶,并置于摇床上培养。
1.2 水蚤样品
研究所用汤氏纺锤水蚤初代水蚤样品取自亚德里亚海泻湖区,于浙江海洋大学国家海洋设施养殖工程技术研究中心浮游生物实验室连续世代培养。水蚤样品置于3 L 培养缸中培养,成体投喂Iso-Rhino 及Rhodo,幼体投喂Iso,其余培养条件同上。培养期间气泵不间断补氧,定期调整气泵大小,保证产生气泡不影响水蚤正常运动。培养过程中定期换水(海水取自舟山市长峙岛揽月湖,筛网粗滤后,0.45 μm 滤膜进行抽滤,高温灭菌后备用)、清理培养缸底藻渣及水蚤粪便,并且定期进行汤氏纺锤水蚤成幼体分离培养[14]。
1.3 仪器
RXZ-500D 智能人工气候箱(宁波江南仪器厂);高压灭菌锅(上海博迅医疗生物仪器股份有限公司);隔膜真空泵(天津津腾实验设备有限公司);SZ650BP 双目体式显微镜(重庆奥特光学仪器有限公司);BM-37XB 倒置生物显微镜(上海彼爱姆光学仪器制造有限公司);TRIzon RNA 提取液(康为世纪生物科技有限公司);荧光定量PCR 试剂盒(日本Takara 公司);逆转录试剂盒(日本Takara 公司)。
1.4 急性、半慢性试验
解剖镜下用筛网分离培养体系中汤氏纺锤水蚤成及幼体,挑取500 只个体健康、肢体完整汤氏纺锤水蚤成体,于5 L 培养缸中暂养3 d,投喂4×105cell·mL-1微藻Iso,每日利用血细胞计算表计算剩余微藻浓度并补充饵料。其间保证曝气,确保水蚤无大规模死亡,移除暂养过程中产生的卵,试验开始取新鲜卵。根据文献设立汤氏纺锤水蚤卵纳米银急性毒理试验纳米银梯度浓度:0、0.1、0.5、1、2.5 和5 mg·L-1[15],每个浓度组设定12 个平行,每孔放置10 枚健康纺锤水蚤卵,纳米银持续暴露48 h,光照(12 h 光照:12 h 黑暗),温度20 ℃,根据试验结果计算得到急性48 h-EC50=0.67 mg·L-1。依据纳米银对汤氏纺锤水蚤卵急性48 h-EC50设定半慢性毒理试验浓度组:0、0.01、0.1、0.5 和1 mg·L-1,处理同上,持续刺激7 d,其他条件同上,计算得到卵半慢性7 d-EC50=0.2 mg·L-1。
1.5 慢性毒理试验
根据纳米银对汤氏纺锤水蚤卵的半慢性7 d-EC50,设立汤氏纺锤水蚤纳米银慢性毒理试验浓度为0,0.2 及0.4 mg·L-1,分别挑取1 对健康及肢体完整的雌雄汤氏纺锤水蚤置于100 mL 结晶皿,每组设立16个平行试验,培养条件同上,水蚤于纳米银下暴露4 d,隔天转移个体至新结晶皿,毒性刺激体系保持一致。观察记录成体死亡情况,若雄性个体死亡则补充雄体,雌性个体死亡则视为试验失败。其间分别记录汤氏纺锤水蚤纳米胁迫下的产卵量,卵孵化率以及排便量。
1.6 超氧化物歧化酶SOD 蛋白酶活性测定
挑取健康且肢体完整汤氏纺锤水蚤成体,暂养3 d。根据半慢性7 d-EC50设定0、1/8 EC50、1/4 EC50和1/2 EC50浓度组,每组50 只取形态大小差异小、运动能力强的成体置于100 mL 结晶皿中,培养体系为50 mL,其间正常投喂饵料,其他条件同上,进行慢性毒理试验。设立取样时间点:0、24、36、48、72 和96 h,将各个组各时间点的汤氏纺锤水蚤置于1.5 mL 离心管,加生理盐水至1 mL,4 ℃暂存。冰浴条件下,研磨枪破碎水蚤样品2 min,于4 ℃离心3 000 r·min-110 min,取上清液0.1 mL,按样品比生理盐水比例为1:9,稀释成1%匀浆,使用南京建成总蛋白含量测定试剂盒,分别测定各个样品OD 值,根据总蛋白标准曲线计算得汤氏纺锤水蚤总蛋白含量(mg prot·mL-1)。稀释样用南京建成超氧化物歧化酶(SOD)试剂盒测定OD 值,结合总蛋白含量计算得纳米银胁迫后汤氏纺锤水蚤SOD 平均活力水平(U·mg-1prot)。
1.7 Ferritin 及Hsp70 基因表达水平测定
挑取健康且肢体完整汤氏纺锤水蚤成体50 只,暂养3 d。根据半慢性7 d-EC50设定0、1/8 EC50、1/4 EC50和1/2 EC50浓度组,处理条件同上,进行慢性毒理试验。设立取样时间点0、24、48、72 和96 h,将30 只水蚤成体置于1.5 mL 离心管。设计汤氏纺锤水蚤Hsp70 及Ferritin 基因的荧光定量PCR 引物及1 对βactin 基因(内参基因)引物,通过Trizol 试剂提取法提取水蚤RNA 后,通过TAKARA 反转录试剂盒反转录成cDNA 模板,对上述反转录产物进行RT-qPCR 试验,目的基因与内参各设置10 个技术重复,排除重复组别中差异较大的数据,其余数据采用2-△△CT法进行基因表达分析。引物见表1。

表1 汤氏纺锤水蚤Hsp70 及Ferritin 基因荧光定量PCR 引物Tab.1 RT-PCR primers for Hsp70 and Ferritin genes in A.tonsa
1.8 结果计算及数据处理
汤氏纺锤水蚤孵化率计算:统计放置的新卵、孵化后的空卵及无节幼体数量进行综合计数,摄食力通过统计粪便数计算。研究相关数据拟用SPSS25.0 软件进行显著性及方差分析,最终计算结果以均值±标准差(x±s)表示,P<0.05 表示有显著性差异。
2 结果
2.1 纳米银胁迫对汤氏纺锤水蚤产卵量及卵孵化率的影响
研究结果显示,纳米银慢性毒理试验毒性积累具备时间效应。汤氏纺锤水蚤受纳米银胁迫时间增加,导致其产卵量逐日下降(图1)。暴露于0.2 mg·L-1纳米银下,汤氏纺锤水蚤前两日产卵量未受到影响,24 h 均产(11±1.5)枚卵,48 h 均产卵(11.5±1.7)枚。水蚤72 h 时产卵量开始出现下降趋势,均产(10±0.9)枚卵,且于96 h 时有最低均产卵量为(6.1±1.8)枚(P<0.05),产卵量相比对照组下降了58.8 %。随着纳米银浓度增加,水蚤产卵量显著下降。汤氏纺锤水蚤暴露于0.4 mg·L-1纳米银下,24 h 均产卵(8.1±1.6)枚,48 h 均产卵(6.3±1.2)枚,72 h时均产卵(5.3±1.3)枚,96 h 时有最低均产卵量为(2.9±0.6)枚(P<0.05),各个时间段产卵量相较于对照组及0.2 mg·L-1纳米银组都有显著下调。

图1 纳米银暴露对汤氏纺锤水蚤产卵量的影响Fig.1 Effect of Ag NPs exposure on egg production of A.tonsa spawning quantity
研究发现卵孵化率与产卵量变化趋势相似,纳米银暴露也会导致汤氏纺锤水蚤的卵孵化率显著性下降,且浓度越高,平均卵孵化率与对照组差异越大(图2)。0.2 mg·L-1纳米银暴露下,汤氏纺锤水蚤新卵孵化率低于对照组卵孵化率(P<0.05),随着暴露时间增加,卵孵化率分别为24 h(65.5±3.1)%,48 h(58.84±4.2)%,72 h(56.09±5.3)%,96 h 孵化率下降至最低值(44.89±4.1)%。0.4 mg·L-1纳米银作为高浓度组,组内卵孵化率相较于对照组和0.2 mg·L-1纳米银组有显著差异,24 h及48 h 孵化率无明显下降,72 h 孵化率为(31.54±2.2)%,96 h有最低孵化率为(15.71±4.1)%,2 个时间段卵孵化率显著性降低(P<0.05)。

图2 纳米银暴露对汤氏纺锤水蚤卵孵化率的影响Fig.2 Effect of Ag NPs exposure on hatching rate of A.tonsa eggs
2.2 纳米银胁迫对汤氏纺锤水蚤排便量的影响
试验结果发现,纳米银暴露使水蚤摄食力大幅下降(图3)。汤氏纺锤水蚤饥饿驯化24 h 后,相比于对照组水蚤的排便量,试验组排便量显著性减少。0.2 mg·L-1纳米银组于24 及48 h 下降幅度较小,从72 h 观察到(79.8±11.9)个粪便(P<0.05),96 h 时有最低排便量(56.8±9.2)个粪便(P<0.05)。0.4 mg·L-1纳米银暴露下,汤氏纺锤水蚤排便量远低于对照组,呈现持续下降的趋势,24 h 平均粪便量(70.7±7.8)个,48 h 平均粪便量(56.5±5.1)个,72 h 平均粪便量(49.8±9.1)个,96 h 观察到最低平均粪便量(27.1±12.2)个。纳米银慢性胁迫下,汤氏纺锤水蚤排便量受纳米银浓度及暴露时间调控。

图3 纳米银暴露对汤氏纺锤水蚤排便量的影响Fig.3 Effect of silver nanometer exposure on fecal pellet of A.tonsa
2.3 纳米银胁迫对汤氏纺锤水蚤SOD 蛋白酶活性的影响
纳米银胁迫对汤氏纺锤水蚤SOD 蛋白酶活力影响见图4,可知纳米银胁迫下12 h 时SOD 酶活皆受到抑制,1/2 EC50纳米银暴露下平均SOD 酶活为(27.5±4.8)U·mg-1prot 远低于对照组(48.6±4.5)U·mg-1prot。1/8 EC50纳米银于24 h 时有最高SOD 酶活(67.6±8.5)U·mg-1prot(P<0.05),该浓度组SOD 活力水平随后回复正常水平。48 h 时2 个高浓度组SOD 活力水平都到达峰值,1/4 EC50SOD 平均酶活为(92.2±3.9)U·mg-1prot(P<0.01),1/2 EC50纳米银胁迫下SOD 平均酶活为(70.3±3.1)U·mg-1prot(P<0.05)。72 h 之后1/4 EC50及1/8 EC50组SOD 酶活恢复正常水平,而1/2 EC50纳米银胁迫抑制SOD合成及酶活力,活性水平远低于对照组水平(P<0.05)。

图4 纳米银暴露对汤氏纺锤水蚤SOD 酶活水平的影响Fig.4 Effect of Ag NPs exposure on SOD enzyme activity of A.tonsa
2.4 纳米银胁迫对汤氏纺锤水蚤Ferritin 及Hsp70 基因表达水平的影响
通过RT-PCR 对汤氏纺锤水蚤Ferritin 及Hsp70 基因,以汤氏纺锤水蚤β-action 为内参基因,选取对照组汤氏纺锤水蚤表达水平作为参照,每个处理进行3 次重复,试验结果表明,纳米银胁迫汤氏纺锤水蚤,其Ferritin 及Hsp70 基因于不同时间段及不同浓度组下呈现出差异性表达(图5,图6)。
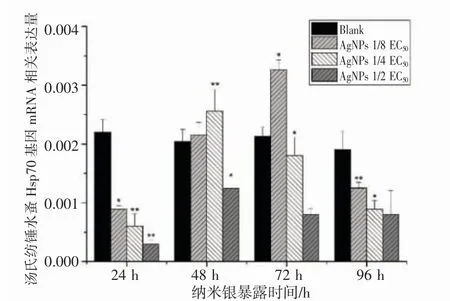
图5 纳米银对汤氏纺锤水蚤Ferritin 基因表达水平的影响Fig.5 Experssion level of Ferritin in A.tonsa with Ag NPs exposure

图6 纳米银对汤氏纺锤水蚤Hsp70 基因表达水平的影响Fig.6 Experssion level of Hsp70 in A.tonsa with Ag NPs exposure
分析结果如下:不同浓度纳米银胁迫下汤氏纺锤水蚤Ferritin 基因mRNA 表达24 h 时皆有显著性低量表达,1/2 EC50纳米银暴露下有最低表达量仅对照组表达水平0.13 倍;48 h 时1/2 EC50有最大表达量,1/4 EC50表达水平到达峰值(P<0.05);72 h 除1/8 EC50组表达量到达峰值后(P<0.05),其余组别皆呈现下降趋势;水蚤受纳米银胁迫96 h 后,各组Ferritin 基因表达水平相对回落。Hsp70 基因mRNA 表达量于纳米银胁迫下24 h 同为低量表达;48 h 时1/2EC50表达量到达峰值(P<0.05),其余组别依旧为低表达;1/8 EC50及1/4 EC50纳米银组于72 h 时有最高表达量(P<0.05)。高浓度纳米银胁迫对Ferritin 及Hsp70 表达量的抑制远高于低浓度纳米银组,说明纳米银胁迫对2 种基因表达水平存在时间效应及浓度效应。
3 讨论
汤氏纺锤水蚤作为典型海洋滤食性浮游生物,因自身缺乏作为排毒器官的肝胰腺,消化道仅由一层上皮细胞组成,并负责水蚤消化道的消化作用。ZHOU,et al[16]通过电镜发现汤氏纺锤水蚤成体会摄入附着纳米镍颗粒的微藻,并在消化道上富集,导致机体摄食力降低。研究发现50 nm 左右的纳米材料不但可以渗透到水蚤的消化器官,甚至于生殖器官和胚胎都可以发现纳米颗粒存在[17]。结果显示纳米颗粒暴露导致汤氏纺锤水蚤产卵量显著下降,说明具备小尺寸效应的纳米银颗粒能够进入水蚤体内,从消化道转移至生殖器官,对水蚤产生生殖毒性[18]。随暴露时间及纳米银浓度增加,毒性积累越多,对水蚤生殖能力抑制越强。LEE,et al[19]研究发现纳米颗粒可以通过胚胎绒毛膜孔,直接胁迫胚胎发育。纳米银可导致斑马鱼Danio rerio 绒毛膜上氧气交换效率降低或停滞,造成胚胎孵化延迟、心率降低、心包水肿和胚胎死亡。汤氏纺锤水蚤卵孵化率同样随纳米银浓度及时间增加而降低,这可能是水蚤成体在纳米银暴露时摄入纳米银颗粒,进入生殖系统,穿刺水蚤卵细胞,干预卵细胞生长发育;另一种可能是卵直接暴露在纳米银环境中,小颗粒或游离的银离子通过细胞膜进入水蚤卵,抑制水蚤卵细胞发育,并产生致死作用。
水环境中纳米颗粒团聚、沉降行为是评估其水环境暴露风险的重要凭证[20]。目前研究表明,纳米材料团聚及沉降对水环境底栖生物更有威胁。纳米材料由于自身尺寸较小,相对比表面积大、表化学能高,环境中的纳米颗粒会自发团聚,或结合其他有机物、有害物质形成复合体[21]。AAL,et al[22]发现纳米二氧化钛团聚,会增加其在水环境中的停滞时间,增加其被水体食物链富集可能性,从而对高等生物产生毒性迫害。纳米银颗粒凭借范德华力、静电力等作用力,纳米银颗粒自发团聚,于水蚤体运动器官上富集,导致水蚤运动能力大幅下降,干扰生命活动,其中摄食力大幅度下降,排便量也相应地减少。纳米银颗粒也能通过摄食途径被汤氏纺锤水蚤成体摄入,于其体内富集,并积累在消化道内,造成水蚤消化能力减弱,观察到的粪便量也因此减少。研究发现纳米氧化锌颗粒会导致大型蚤肢体受损,体内脂类过氧化,能量储备能力下降。而通过摄食的纳米颗粒会导致水蚤体内与氧运输和抗氧化酶有关的指数下降[23]。因此纳米银颗粒进入汤氏纺锤水蚤正常健康细胞中,可能引起各种细胞功能的自我调节,如发生氧化应激、细胞本身或细胞器膜的损伤。汤氏纺锤水蚤在纳米银胁迫之下,游离银离子对其器官细胞产生强烈的氧化反应,并在暴露初期不彻底破坏其抗氧化系统情况下,短期内促进机体合成超氧化物歧化酶对抗这种氧化反应,但随着暴露时间及纳米银浓度增加,对水蚤抗氧化系统会造成不可逆的损伤,SOD 酶合成水平逐渐下降,进而导致机体生殖能力、运动能力下降,并机体逐渐失去活力或者死亡。
热激蛋白作为应激类功能蛋白家族,在细胞受到刺激就会大量合成。Hsp70 是热激蛋白家族中研究比较透彻的一种,能对抗外界不利因素。Ferritin 铁蛋白存在于各种有机体体内,参与储存及释放铁离子,同样可以反映机体对外界环境产生应激反应。研究表明,日本虎斑猛水蚤Tigriopus japonicus 暴露在纳米塑料颗粒中,水蚤体内活性氧水平上升,抗氧化剂相关的基因表达受到调节,抗氧化酶的活性显著变化[18]。研究表明暴露于纳米银中的奥利亚罗非鱼,其所有组织中均观察到高水平的Hsp70 表达,加之肌肉、肾脏和肝脏等免疫组织化学染色反映了罗非鱼Oreochromis mossambicus 对纳米颗粒强烈的应激反应[24]。本研究中纳米银胁迫汤氏纺锤水蚤Hsp70 及Ferritin 2 个基因,二者的表达水平存在相对时间效应及相对浓度效应。纳米银初胁迫对水蚤机体造成损伤,24 h 有显著低量表达。试验期间会产生表达峰值,到达峰值后,反而会呈现显著下降的趋势,说明汤氏纺锤水蚤成体长期暴露下会产生基因毒性,且这个毒性会随时间积累。2 种基因变化趋势说明汤氏纺锤水蚤机体会受纳米银胁迫,受到氧化损伤,机体产生应激反应,Hsp70和Ferritin 短期内会大量表达,随着毒性积累超过水蚤能够承受的阈值,可能会影响汤氏纺锤水蚤的生殖能力及摄食能力,同样可能造成抗氧化相关基因不可逆的损伤,甚至因体内ROS 过量导致死亡。
综上所述,纳米银暴露对汤氏纺锤水蚤生殖力,酶活力以及抗氧化基因表达水平皆存在较强抑制作用。水蚤体内抗氧化机制存在纳米银低浓度初期诱导,长期及高浓度暴露有显著抑制趋势,因此汤氏纺锤水蚤对纳米银胁迫的响应存在着明显的时间效应以及浓度效应。本文通过研究纳米银暴露对汤氏纺锤水蚤生理指标、酶活力指标及基因表达水平响应机制,从生态毒理学层面初步分析纳米银胁迫对汤氏纺锤水蚤的毒性作用机制,为海洋桡足类毒理学研究提供基础数据,为海洋纳米银污染监控及治理提供监测手段。
