面向探地雷达应用的活立木木材介电常数受主要应用条件的影响
2022-05-18李怡娜徐国祺王立海阚相成谢军明
李怡娜,徐国祺,王立海,阚相成,谢军明
(东北林业大学 工程技术学院,黑龙江 哈尔滨 150040)
受各种因素的影响,活立木生长过程中会出现树干内部腐朽、中空、开裂等情况,给树木生长和应用带来严重的损害。为了能及时对树干内部腐朽进行检测,达到早发现早治理的目的,学者们开发出了很多木材无损检测技术,常见的木材无损检测技术有:应力波法、电阻率法、超声波、探地雷达等[1-2]。
在众多树木无损检测技术中,探地雷达技术是近些年来新发展起来的无损检测技术,极具发展前途[3]。该技术具有无损性、检测快、操作简单等优点,能够对树木内部的健康状况进行检测,并且可以直观地观察到腐朽的分布。随着探地雷达技术的不断发展,越来越多的国内外学者应用该技术来检测树木内部腐朽。如Butnor 等[4]发现探地雷达在松柏类、针叶类树木的检测中,能够很好地对树皮表面的孔洞以及内部空洞进行检测,同时对被子植物的检测也具有不错的潜力。田凌鸿等[5]利用树木雷达对古侧柏的空腐规律进行研究,发现其腐朽与树干高度、立地条件、年龄等有关。文剑等[6]针对活立木结构复杂导致雷达波形难以解释的问题,开发出了基于雷达波的活立木内部分析软件。
将探地雷达应用于树木检测主要是依据木材内部的腐朽部分与健康部分存在的电性差异,因此应用探地雷达技术检测的理论基础是介电常数,对木材的介电常数进行研究是十分必要的[3,7]。介电常数是介电材料在交流电场中衡量介质极化的程度[8]。木材的介电常数是指:木材在交流电场介质极化状况和储存电能能力的一个量[9]。木材是一种天然生长的有机材料,主要由纤维素、半纤维素、木素和木材抽提物组成,它的极化主要是指纤维素、半纤维素、木素等有机大分子中的极性基团以及湿材中的水分子等偶极子在电场作用下的取向运动。目前,对木材介电特性的研究主要集中于频率、含水率、纹理方向等方面。白同仁等[10]对含水率处于纤维饱和点以下的马尾松的介电性质进行研究,指出了含水率、纹理方向、频率等对马尾松介电性质的影响,具有重要参考意义。吕静霞等[11]通过对不同含水率下木材的介电常数进行修正,提高了树木雷达的检测精度。但涉及温度方面的研究较少,且所涉及到的频率、含水率等区间较窄,缺少与探地雷达无损检测相结合的木材介电常数的研究。由于探地雷达应用过程中不同应用条件会对介电常数产生不同影响,从而导致检测结果的不准确。因此为了探究不同因素对介电常数的影响规律,同时为提高探地雷达检测的准确性提供理论基础。本研究以东北常见树种樟子松、红松、大青杨和水曲柳为研究对象,以探地雷达常见的应用条件为基础,对不同含水率(10%~100%),不同外电场频率(40 Hz~110 MHz),不同环境温度条件(-15~20℃)下的试件进行介电常数的测量。基于所得的数据,应用SPSS 软件进行相关性分析以及多元回归分析,探究外电场频率、含水率以及探地雷达常见工作温度对介电常数的影响以及影响程度,丰富了不同条件下木材介电常数的变化规律,对后续进一步研究木材介电特性具有重要意义。同时,与探地雷达的应用相结合为修正活立木的介电常数,提高探地雷达的检测精度建立基础。
1 材料与方法
1.1 材料
本试验所用样品采自黑龙江省方正县林业局星火林场(45°43′5.73″N,129°13′34.37″E)。实验选取了樟子松Pinus sylvestris、水曲柳Fraxinus mandshurica、大青杨Populus ussuriensis和 红松Pinus koraiensis4 个树种,分别加工成规格为25 mm×25 mm×3 mm(弦向×横向×径向)的方形无瑕疵小木片,弦切面(阴影面)为检测面,见图1。基本密度分别为0.228、0.255、0.204、0.207 g/cm3,将试件作抛光处理并在试件中心镀半径为5 mm 的圆形电极。
图1 实验材料制备Fig.1 The preparation of experimental materials
1.2 主要仪器与设备
实验中所用到的仪器与设备见表1。

表1 实验仪器与设备Table 1 Experimental instruments and equipment
1.3 实验方法
基于探地雷达常见的应用条件,本实验对参数进行等差梯度设计,设置含水率范围为10%~100%,外电场频率范围为40 Hz~110 MHz,环境温度范围为-15~20℃。
将试件放入恒温恒湿箱中,通过调节恒温恒湿箱的温度(0、5、15、20℃)、湿度(10%、20%、30%、40%),使试件达到所设定的温度和含水率,并进行测量。对于含水率要求为60%、80%、100%的试件,采用恒温恒湿箱与人工浸泡相结合的方法,先将试件在恒温恒湿箱中调节含水率至纤维饱和点以上,随后采用人工浸泡的方法调节含水率至60%、80%、100%。对于温度为-5、-15℃的试件,采用先恒温恒湿箱调湿至设定含水率后立即用塑料膜包裹并将其密封,随后将冰柜温度分别设定为-15、-5℃。将密封好的试件放置在冰柜内部12 h,以使试件内外温度相同,再进行后续测量。为了防止测试过程中试件的温度发生明显变化,测试每一试件的时间控制在30 s之内。测量前,采用红外测温仪检测试件的温度,确保木材的温度与设定的温度值相差在±0.5℃ 范围内。最后将所有试件放入烘干箱内,在105℃下烘干24 h 至恒质量,并记录试件的绝干质量m0。
将试件泡于水中,每隔一段时间对试件进行称重,当试件达到所需含水率时,则立即用塑料膜将其密封并记为湿材质量m。试件的含水率计算公式(1)如下:

式(1)中,M为试件的含水率;m为试件测量时的质量;m0为试件的绝干质量。
1.4 介电常数的计算
木材的介电常数一般用ε来表示,它满足如下关系式(2):

式(2)中:ε为介电常数;ε0为真空介电常数,ε0=8.85×10-12F/m;S为电极面积;d为电极直径,c为电容。
1.5 数据处理与分析
使用介质频谱测试仪自带的程序Excel VBA软件进行数据的采集与保存。将数据进行插值处理,并采用Origin 8.0 软件绘制介电常数变化趋势图。采用SPSS 20.0 软件进行相关性分析以及多元回归分析。
2 结果与分析
2.1 外电场频率对木材介电常数的影响
图2所示为室温条件下,不同含水率木材的介电常数随频率变化的趋势图。由图2中可看出,当环境温度和含水率一定时,随着外电场频率的增大,4 种木材的介电常数不断下降并趋于平缓。当外电场频率小于31 MHz 时,介电常数的下降速率较快,而在外电场频率大于31 MHz 后,随着频率的增大,介电常数下降的速率逐渐减慢并趋于稳定。以含水率为100%的樟子松为例,外电场频率为1~31 MHz 时,其介电常数下降速率为-2.6 MHz-1,当外电场频率为31~110 MHz 时,其下降速率为-0.08 MHz-1。含水率也会影响下降的速率,含水率越高,木材介电常数随外电场频率下降的速率越快。以樟子松为例,含水率为100%的介电常数的下降速率为-0.82 MHz-1,含水率为20%的介电常数下降速率为。从数据中可以明显看出,含水率为100%的木材介电常数的下降速率快于含水率为20%的木材。

图2 室温环境下不同含水率木材介电常数随频率变化的趋势(a-红松,b-樟子松,c-杨树,d-水曲柳)Fig.2 Variation trend of wood dielectric constant with different frequency conditions and moisture contents(a-Pinus koraiensis,b-Pinus sylvestris,c-Fraxinus mandshurica,d-Populus ussuriensis)
Tsutsumi 等[12]研究发现:在含水率和温度条件一定时,介电常数随外电场频率的增大而减小,且含水率不同,曲线形状不同。出现上述趋势的原因是介质的极化通常可分为4 类,分别是电子极化,原子极化,偶极极化(取向极化)和界面极化[13]。其中,前两种极化所需时间极短,约10-12~10-15秒,而后两种极化所需时间更久一些,偶极极化所需时间在10-9秒以上,界面极化则需要几秒至几十秒的时间。介电常数会随着外电场频率的增大而减小是因为当介质处在低频率的外电场中时,4 种极化均可建立,此时木材介电常数值最大,木材的极化强度也最大。而随着频率的不断增大,木材内部的极性分子被快速极化,但是越来越跟不上电场的变化,极化出现滞后,偶极子的运动方向与频率方向相反,极化强度降低,因此介电常数呈减小趋势[14]。而当外电场频率增大至31 MHz 后时,4 种木材的介电常数随着频率的增加下降趋势缓慢。这是由于在射频范围内,影响木材介电常数的主要是偶极极化,而当电场的频率达到红外光频和可见光频范围内时,电子极化和原子极化才会来不及建立。故当电场频率超过一定值时,介电常数会趋于稳定[15]。同时,木材的介电常数也受到水分子的影响。含水率越高,木材内部的极性分子越多,当频率变化时,就会有越多的极性分子剧烈运动,使其介电常数随频率的增大降低得越快。故含水率越大,外电场频率对介电常数的影响越明显。
2.2 含水率对木材介电常数的影响
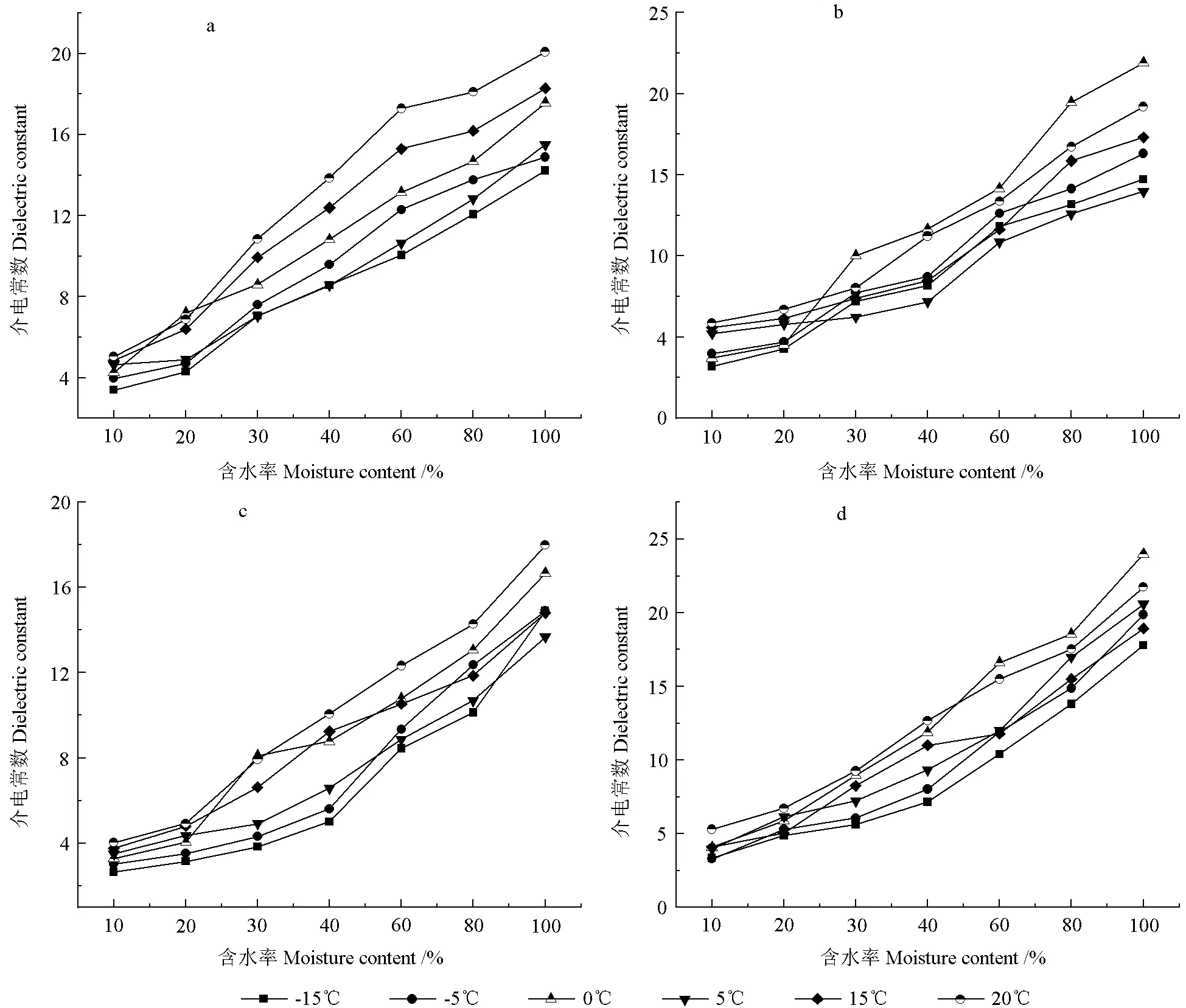
图3 相同频率条件下不同温度的木材介电常数随含水率变化的趋势图(a-红松,b-樟子松,c-杨树,d-水曲柳)Fig.3 Variation trend of wood dielectric constant with different moisture contents and temperatures (a-Pinus koraiensis,b-Pinus sylvestris,c-Fraxinus mandshurica,d-Populus ussuriensis)
图3分别为红松、水曲柳、樟子松和大青杨在相同的外电场频率(15 MHz)下,木材介电常数随含水率变化趋势图。从图3中可以看出:当频率相同时,含水率变化对4 种木材的介电常数的影响规律具有相似性。即随着含水率的增加,木材的介电常数呈增大趋势,且木材含水率高于纤维饱和点后的介电常数增大速率整体大于含水率处于纤维饱和点以下时的增长速率。当含水率处于纤维饱和点以下时,介电常数的增长相对较为缓慢,而随着含水率的增加,当木材含水率达到纤维饱和点附近时,介电常数明显增加。以樟子松为例,20℃下含水率从10%增加至20%时,其介电常数增加了1.8,而含水率从20%增加至30%时,介电常数增加了3.5。当含水率高于纤维饱和点时,介电常数与含水率呈线性增长关系。
根据木材中的水分和木材的结合形式,可将木材中的水分分为3 类,分别是:化合水、吸附水和自由水[16-17]。这3 类水中,化合水由于含量较少且在高温条件下才会对木材的介电常数产生影响,因此在本研究中,只考虑吸附水和自由水的影响,而吸附水与自由水又由纤维饱和点加以区分。纤维饱和点以下木材中不含有自由水,而在纤维饱和点以上,木材中存在自由水和吸附水[18]。
如图3中所示,当含水率处于纤维饱和点以下时,介电常数增大缓慢,这是因为当含水率较低时,木材中只存在吸附水且水分子主要存在于细胞壁上,此时属于单分子层度的吸附,水分子与木材细胞壁物质结合紧密,不能自由移动。因此,在外电场的作用下,水分子只能同相结合的各极性基团作取向运动,水分子此时对木材的极化效应影响较小。所以在纤维饱和点以下,含水率即使增加,木材的介电常数的增加也是比较缓慢的。
当含水率增加至纤维饱和点左右时,木材中的水分子增多,此时木材内部已经由单分子层度的吸附逐渐转变成多分子层度的吸附,吸附层厚度增加,吸附力减弱。此时在外电场的作用下,外层水分子和中层水分子能够分别作独立的取向运动以及半独立的取向运动,此时水分子对木材的极化效应影响较大,故此时的介电常数有了明显的增大。
当含水率高于纤维饱和点时,木材中的水分包含有细胞壁中饱和的吸附水,以及细胞腔中的自由水,并且由于木材吸水膨胀,空隙度变小,吸附水大部分变成了自由水,不再受木材的束缚,此时的介电常数主要取决于自由水在木材中所占的比例[19]。水的介电常数比较高,可达80 左右,对木材的极化起到了明显作用。因此,在纤维饱和点以上,随着含水率的增加,介电常数呈增长趋势,与纤维饱和点以下的变化趋势相比,整体速率较快。同时,温度也会对增长速率产生一定的影响。与我国学者曹绿菊、白同仁、李先泽等[8,10,19]均针对含水率对木材介电常数的影响研究规律类似。
2.3 温度对木材介电常数的影响
图4分别为红松、水曲柳、樟子松和大青杨在相同频率(15 MHz)下,木材介电常数随温度变化趋势图。观察图4可以看出,当温度处于-15~20℃区间内,随着温度的逐渐升高,木材的介电常数整体呈现出增大的趋势。当木材含水率处于纤维饱和点以下时,随着温度的升高,介电常数平稳地增加,在0℃附近未出现明显的突变。而当木材的含水率高于纤维饱和点时,随着温度升高至0℃左右时,介电常数会产生突变,随后出现下降趋势,随着温度继续升高至5℃附近时,介电常数又再次增大。与樟子松、大青杨、水曲柳相比,红松在含水率为20%时就有相似的规律。
温度则是主要通过改变水的状态来影响木材的介电常数。当温度较低,且木材含水率在纤维饱和点以上时,木材中的自由水主要以固态的形式存在于木材中,木材作为含有极性分子或极性基团的类晶体固体介质,其中的偶极基团被束缚在固体结构中,晶格的刚性阻碍了偶极基团的取向运动,偶极基团无法作转向运动,因此极化程度较低,相应的介电常数也低。随着温度升高,偶极基团获得了能量,这使它们能够重新做取向运动,因此随着温度升高,介电常数呈增加趋势[20]。
随着温度升高,0℃左右介电常数发生突变的原因是当木材含水率高于纤维饱和点,即木材细胞中含有自由水时,随着温度的降低,木材内部的水分会发生相变,即由液态转变为固态,变成了冻结水[16]。而液态水的介电常数远远大于冰,因此,当温度升高,木材中的水分发生相变,即由固态转变为液相时,木材的介电常数会有明显的增加。而在0~5℃范围内,随着温度的继续升高,分子的无规则布朗运动加剧,这使得偶极子的排列变得无序,介电常数出现下降;当温度继续升高,偶极子由开始的跟不上电场变化逐渐能够跟得上电场变化,介电常数又会呈现出增大的趋势。
当木材含水率低于纤维饱和点时,由于木材中没有自由水的存在,温度的降低并不会引起木材内部水分的相变,因此当木材含水率低于纤维饱和点时,温度对木材介电常数的影响不大,木材的介电常数会随着温度的升高缓慢增大。在零度以下,并不是木材中所有的水分都会转化为冰,而是一部分水分以冰的形式存在,而另一部分仍处于不结冰的状态,冻结木材中的不冻结水分含量取决于含水率和负温度。木材中冰与不冻结水分的比例是决定潮湿木材在负温度下介电特性变化的原因。吴晔[15]、James[21]也发现了类似的规律。
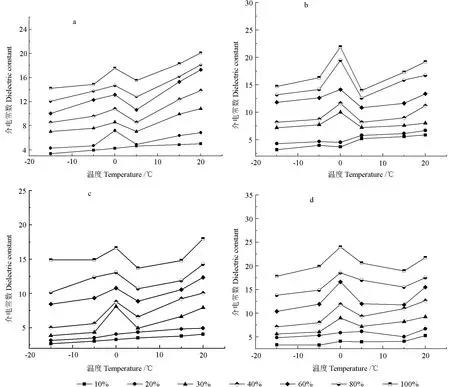
图4 相同频率条件下不同含水率木材介电常数随温度变化的趋势图(a-红松,b-樟子松,c-杨树,d-水曲柳)Fig.4 Variation trend of wood dielectric constant with different temperatures and moisture contents (a-Pinus koraiensis,b-Pinus sylvestris,c-Fraxinus mandshurica,d-Populus ussuriensis)
2.4 木材的介电常数与各因素之间的关系研究
由于4 种木材的介电常数变化规律相似,因此本研究以红松为例,探究介电常数与温度、含水率和频率的相关关系,分析结果见表2。相关性分析表明介电常数与含水率和频率具有较为显著的相关关系(P<0.01)。介电常数与频率呈较为显著的负相关关系,与含水率呈较为显著的正相关关系,而与温度无显著相关性。介电常数与含水率的相关程度最高,频率次之,相关程度最低的因素是温度。出现这种情况的原因可能是由于实验所设置的温度区间(-15~20℃)较窄,在此温度区间内仅取6 个温度点,且在此温度区间内介电常数随着温度变化的波动较大,在零摄氏度附近出现突变,故介电常数和温度没有呈现出显著的线性关系。同时,相关系数不是很高的原因在于温度、含水率只是影响木材介电常数众多因素中较为重要的因素,木材的介电常数还受其他因素的影响。
本研究应用SPSS 20.0 软件,基于所测的数据来进行多元回归分析。设置自变量为环境温度、外电场频率、含水率,因变量为介电常数,得到4种木材的介电常数与含水率、环境温度和外电场频率的多元回归方程表,见表3。
表3公式中,y、x1、x2、x3分别表示木材的介电常数、含水率、外电场频率和环境温度。根据标准误差检验,该回归方程通过了P=0.01 的显著性检验,表明回归的整体效果显著。从回归方程可以看出,介电常数随着含水率、温度的增大而增大,随着外电场频率的增大而减小。回归分析结果表明,在此温度区间内,含水率和频率对木材介电常数的解释度更高,是影响介电常数的主要因素,可以应用含水率、外电场频率和环境温度来预测木材介电常数。在应用探地雷达对东北地区活立木进行健康检测时,可先提取活立木含水率、环境温度,以及选定外电场频率等信息,通过回归模型修正该活立木的介电常数,从而获得更加精准的介电常数,提高检测的准确性。

表2 红松的介电常数与频率、含水率和温度的相关系数†Table 2 Correlation coefficients of the dielectric constant of Pinus koraiensis with frequency,moisture content and temperature

表3 4 种木材的多元线性回归模型†Table 3 Multiple linear regression models of the four wood types
在应用探地雷达对活立木内部缺陷进行检测时,由于其内部可近似看为层状结构,因此垂直分辨率是要关注的重点。垂直分辨率[22]可表示为式(3):

式(3)中可以看出,频率f与垂直分辨率R和介电常数ε成反比。即树木雷达天线频率选得越大,垂直分辨率越小,探测的目标越大,探测的深度越浅,该频率下的介电常数也越小。
不同季节、不同地区、不同树种,甚至是同一天,不同时间段活立木的含水率都不尽相同。东北大部分地区冬季会长达半年左右,树木很长一段时间内处于冰点温度以下。王玉婷等[23]通过对东北林业大学知园林内的杨树进行24 h 含水率检测发现,其含水率最高可达98%,最低可达50%,最高温度可达5℃,最低温度可达-8℃。将温度、频率(选用110 MHz)和含水率数据代入回归方程中,得到其介电常数为14、6,与系统默认的介电常数值10 相比,存在差距。故应用探地雷达进行活立木腐朽检测时,活立木的介电常数不能一概而论,应合理地设置介电常数。
3 结论与讨论
3.1 结 论
本研究以红松、樟子松、杨树和水曲柳为研究对象,通过测量不同条件下4种木材的介电常数,分析了含水率、环境温度和外电场频率对木材介电常数的影响。研究发现:
1)频率、温度和含水率均会对木材的介电常数产生不同的影响。随着外电场频率的增大,介电常数呈减小趋势,且下降趋势逐渐趋于平缓。同时,含水率越大,介电常数减小的速率越快。
2)随着含水率的增加,木材的介电常数呈增大趋势,且木材含水率高于纤维饱和点后的介电常数增大速率整体大于含水率处于纤维饱和点以下时的增长速率。
3)在-15~20℃温度区间内,介电常数随着温度的升高呈现出整体增大的趋势。含水率高于纤维饱和点时的增长速率大于含水率低于纤维饱和点时的增长速率,且当木材含水率高于纤维饱和点时,会在0℃左右出现突变。
4)在环境温度为-15~20℃区间内,介电常数与频率和含水率具有显著相关性,与介电常数相关程度由高到低依次是含水率、频率、温度。通过回归分析,获得了介电常数随3 因素变化的回归模型,可用来预测不同条件下活立木的介电常数。
3.2 讨 论
当前对木材介电常数的研究主要集中于单因素分析,缺少将介电常数与探地雷达应用相结合的分析。本研究将介电常数与探地雷达的应用条件相结合,检测了不同应用条件下木材的介电常数,并利用相关系数法来分析各影响因子与介电常数的关系,建立了多元回归方程。
应用探地雷达对东北地区活立木进行健康检测时,可先提取活立木含水率、环境温度,以及外电场频率等信息,通过回归模型修正该活立木的介电常数,从而获得更加精准的活立木健康状况。研究结果可用于行道树树木、古树名木,以及珍贵树种等的健康检测,为保护人员的生命与财产安全以及活立木的保护及复壮提供理论基础。本研究局限性在于选取的树木种类较少,选取的影响因子范围具有一定的局限性。因此,在今后的研究中,应研究更多树种,更广的应用条件范围,结合更多影响因素如密度、纹理方向等,来构建更加完善的介电常数预测模型。
