七种木本植物水分胁迫的适应性研究
2022-05-18刘小冬孙叶熔汤粤婷王敏肖冰
刘小冬 孙叶熔 汤粤婷 王敏 肖冰
(1.仲恺农业工程学院园艺园林学院园林系 广东广州 510230;2.澳门城市大学创新设计学院 澳门 999078)
湿地是城市生态系统的重要组成部分,具有游憩、防止水土流失、水质净化、多样性保护等功能[1-2]。植物是湿地的重要景观要素,其群落构建直接影响景观质量与生态功能发挥[3-4]。当前湿地植物群落构建以草本植物为主,但存在更换维护成本高、易冬枯产生二次污染等问题[5-6],而引入耐水湿木本植物,构建复层植物群落有助于缓解此类问题。虽然,已有研究表明,红花玉蕊、串钱柳等植物在高水饱和度土壤具有较好的适应性[7-9]。然而,城市河流、湖泊与湿地存在季节性洪水或突发性暴雨造成岸际水位涨落,导致沿岸植物生境水分条件发生变化,面临水淹、湿润、干旱等生境。而不同程度的水分胁迫影响着植物生长状况,并与植物光合作用密切相关[10-11]。众多学者通过测量净光合速率、胞间CO2浓度、气孔导度等光合指标分析植物在不同土壤湿度下生理特征[12-16];或通过茎干增粗、不定根形成等树种的形态结构和解剖特征[17],并评估其水分适应性。以植物光合指标、形态特征研究红花玉蕊、串钱柳等7种优选木本植物在不同水分条件下的适应性,以期为华南地区可持续湿地植物景观的构建提供参考。
1 材料与方法
1.1 材料
试材为红花玉蕊(Barringtonia racemosa)、串钱柳(Callistemon viminalis)、对叶榕(Ficus hispida)、蒲桃(Syzygium jambos)、铁冬青(Ilex rotunda)、木芙蓉(Hibiscus mutabilis)、木槿(Hibiscus syriacus)7种木本湿生植物1年生健康小苗木(详见表1)。

表1 试验植物种类
1.2 方法
1.2.1 试验设计试验地点位于广东省从化市试验大棚内,于 63个规格相同的白色塑料箱(81 cm×57 cm×59 cm)中进行;试验时间为2020年10月,当月气温23~32℃。试验土壤为塘泥。植株高度均为60~70 cm且株型基本一致。分为3种不同土壤湿度处理∶处理1(K1),保持土壤处于长期湿润状态(土壤含水率80%以上);处理2(K2),对植物进行淹水,淹过基部 3 cm,划定水位线刻度,当低于原始水位线时及时补水;处理3(K3),保持土壤干旱状态,30 d不浇水,并保证外界环境条件相同。每种土壤湿度处理各设3个重复,每个重复3株。试验时长为30 d。
1.2.2 项目测定使用 Li-6800 XT便携式光合测定仪对每株植物的光合作用进行测定,采用的外接红蓝光源设置光合有效辐射强度(PAR)为1 500、1 200、1 000、800、600、400、200、150、100、50、25、0 μmol/(m2·s),外接 CO2小钢瓶设置 CO2流速设置为 400 μmol/(m2·s),温度控制为25~28℃。每株植物选择健康成熟的叶片进行测定,连续测定3 d,时间为每天上午9:00—12:00。测试前先将叶片置于饱和光强下进行诱导,每个光合有效辐射强度下平衡200 s后仪器自动测定,测定指标包括:净光合速率(Pn)变化、胞间CO2浓度(Ci)变化以及气孔导度(Gs)变化;同时,观察不同土壤湿度条件处理下种植 10 d和种植30 d后各个植株的生长状况和叶片颜色。
1.2.3 数据分析使用 Excel分析数据;利用SPSS 25.0进行各项光合数据之间差异显著性的分析。
2 结果与分析
2.1 植物生长状态变化分析
图1、2分别为湿润(K1)、水淹(K2)、干旱(K3)处理下,木槿和蒲桃的生长状况。蒲桃和木槿仅在湿润(K1)处理下,整体长势良好,起始10 d长出新叶,试验30 d植株长势良好,叶片颜色正常,植株试验前后均无明显变化。在水淹(K2)和干旱(K3)处理下,蒲桃和木槿均能存活但整体长势一般,水淹(K2)处理下起始10 d有长出新叶,但试验30 d后木槿少数叶片偏小,未长出不定根,而蒲桃长势较差,植株节间较长,叶色偏浅;在干旱(K3)处理下起始10 d亦长出新叶,但蒲桃和木槿大部分叶片出现黄斑、偏小且叶缘卷曲现象,轻度缺水萎蔫。

图1 木槿生长状况
图3~7分别为湿润(K1)、水淹(K2)、干旱(K3)处理下,红花玉蕊、串钱柳、对叶榕、铁冬青和木芙蓉的生长状况。5种木本植物在湿润(K1)和水淹(K2)2种处理下,整体长势较好;在干旱(K3)处理下,整体长势一般甚至不良。在湿润(K1)处理下,起始10 d均有新叶长出且叶色正常,试验30 d后叶片颜色依旧正常,植株试验前后均无明显变化。在水淹(K2)处理下,起始10 d均有长出新叶且叶色正常,试验30 d后串钱柳、对叶榕、铁冬青、木芙蓉叶片形态与湿润(K1)处理下的相似,叶色正常,极少部分稍有变浅,植株茎基部稍有膨大且60%的植株长出稀疏的不定根,其中,木芙蓉大部分会开花;而红花玉蕊有少部分叶片颜色变淡,未长出不定根。在干旱(K3)处理下,起始10 d均有长出新叶,但试验30 d后不同植物表现出不同的胁迫状态,串钱柳植株生长较慢,几乎无新叶长出,植株出现明显缺水现象;红花玉蕊大部分植株轻度缺水萎蔫且叶片枯黄,少部分则出现严重生长不良、矮小萎蔫的现象;对叶榕、铁冬青和木芙蓉叶片均出现黄斑褐化、偏小且叶缘卷曲现象,轻度缺水萎蔫,其中对叶榕仅小部分叶片出现,铁冬青有半数叶片出现,木芙蓉有大部分叶片出现且不开花。

图3 红花玉蕊生长状况

图2 蒲桃生长状况

图4 串钱柳生长状况

图5 对叶榕生长状况

图6 铁冬青生长状况

图7 木芙蓉生长状况
2.2 水分胁迫对7种植物光合指标的影响
2.2.1 净光合速率(Pn)变化分析由表2、3可知,在3种土壤湿度下,串钱柳的Pn由大到小依次为:水淹(K2)>湿润(K1)>干旱(K3),而其余6种木本植物的Pn由大到小依次为:湿润(K1)>水淹(K2)>干旱(K3)。串钱柳在水淹(K2)处理下的Pn最高,其余6种木本植物在湿润(K1)处理下的Pn最高;而7种植物均在干旱(K3)条件下的Pn最低,其Pn显著低于湿润(K1)处理与水淹(K2)处理。
通过单因素方差分析(表2),不同水分条件处理之间,除串钱柳外,其他6种植物的Pn均存在显著差异;根据多重比较结果(表3),红花玉蕊、对叶榕、铁冬青、木芙蓉的湿润(K1)与水淹(K2)之间Pn均差异不显著,而蒲桃、木槿则在湿润(K1)下Pn均显著高于水淹(K2);除串钱柳外,其余6种木本植物在湿润(K1)下Pn均显著高于干旱(K3);对叶榕、蒲桃、铁冬青、木芙蓉、木槿的水淹(K2)与干旱(K3)之间均无显著差异,而红花玉蕊、串钱柳在水淹(K2)下均显著高于干旱(K3)。

表2 七种木本植物在不同土壤湿度下Pn的单因素方差分析

表3 水分胁迫对7种木本植物Pn影响的多重比较分析
2.2.2 气孔导度(Gs)变化分析从表4、5可看出,在3种土壤湿度下,串钱柳的Gs由大到小的顺序为:水淹(K2)>湿润(K1)>干旱(K3),而其余6种木本植物的的Gs由大到小的顺序为:湿润(K1)>水淹(K2)>干旱(K3)。串钱柳在水淹(K2)处理下的Gs最高;其余的6种木本植物都在湿润(K1)处理下的 Gs最高,在干旱(K3)处理下最低。但不同土壤湿度对各组植物之间的Gs的影响不大。
单因素方差分析结果(表4)表明,不同水分条件处理之间,除铁冬青和木芙蓉外,红花玉蕊、串钱柳、对叶榕、蒲桃、木槿的Gs存在极显著差异(p<0.01)。根据多重比较结果(表5),除蒲桃和木槿在湿润(K1)下均显著高于水淹(K2)外,其余3种木本植物在湿润(K1)与水淹(K2)之间均差异不显著;5种木本植物在湿润(K1)下均显著高于干旱(K3);蒲桃和木槿的水淹(K2)与干旱(K3)之间均无显著差异,而红花玉蕊、串钱柳、对叶榕在水淹(K2)下均显著高于干旱(K3)。
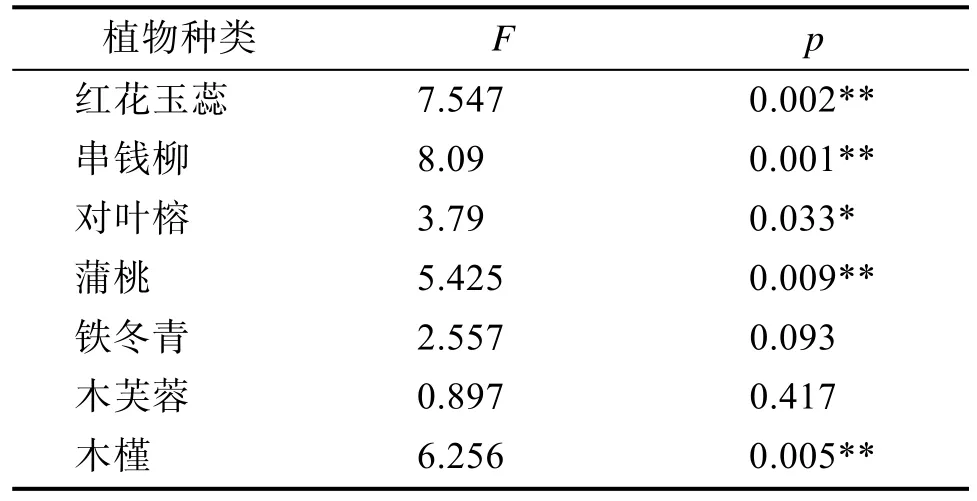
表4 七种木本植物在不同土壤湿度下Gs的单因素方差分析

表5 水分胁迫对7种木本植物Gs影响的多重比较分析
2.2.3 胞间CO2浓度(Ci)变化分析由表6、7得出,在3种土壤湿度下,串钱柳的Ci由大到小依次为:干旱(K3)>湿润(K1)>水淹(K2),而其余6种木本植物的Ci由大到小依次为:干旱(K3)>水淹(K2)>湿润(K1)。串钱柳在水淹(K2)处理下的Ci最低;其余的 6种木本植物都在湿润(K1)处理下的Ci最低,干旱(K3)处理下的最高。但不同土壤湿度对各组植物之间的Ci影响不大。
单因素方差分析结果(表6)表明,不同水分条件处理之间,除红花玉蕊外,其他6种植物的Ci均存在显著差异(p<0.05),且不同土壤湿度对木槿与对叶榕的Ci存在极显著的差异(p<0.01)。根据多重比较结果(表7),串钱柳、对叶榕、铁冬青、木芙蓉的湿润(K1)与水淹(K2)之间均差异不显著,而蒲桃和木槿则在湿润(K1)下均显著低于水淹(K2);除串钱柳外,其余 5种植物在湿润(K1)下均显著低于干旱(K3);蒲桃、铁冬青、木槿、和木芙蓉的水淹(K2)与干旱(K3)之间均无显著差异,而串钱柳和对叶榕在水淹(K2)下均显著低于干旱(K3)。
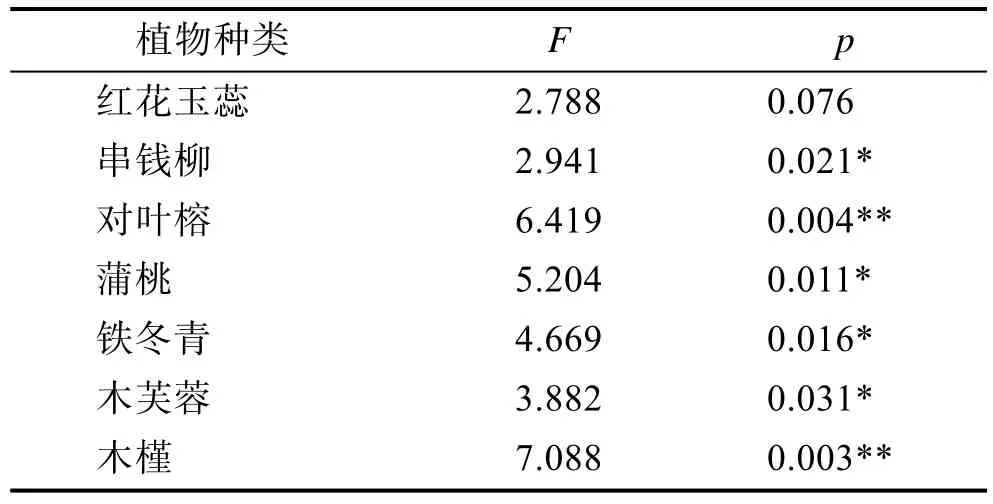
表6 七种木本植物在不同土壤湿度下胞间CO2浓度的单因素方差分析

表7 水分胁迫对7种木本植物胞间CO2浓度影响的多重比较分析
3 讨论与结论
3.1 讨论
水分胁迫会对植物生长状况造成一定程度的影响,能使植物的形态发生改变。不定根是植物对水分胁迫的积极适应,具改善植物与土壤间气体交换的作用[18]。本试验中,蒲桃和木槿仅在湿润(K1)处理下,整体长势良好,在水淹(K2)和干旱(K3)处理下,均能存活但长势一般,部分叶片出现黄斑、偏小且叶缘卷曲现象,说明蒲桃和木槿的耐湿能力较好,但耐淹耐旱能力不足。而红花玉蕊、串钱柳、对叶榕、铁冬青和木芙蓉在湿润(K1)处理和水淹(K2)处理下,整体长势良好,均有新叶长出,叶片无萎蔫且叶色正常,水淹(K2)处理下除红花玉蕊外的植株大部分长出稀疏不定根,且木芙蓉大部分会开花,而在干旱(K3)处理下均能存活但长势一般甚至不良,出现叶片枯黄、褐化、轻度缺水萎蔫的现象,说明以上5种植物具有较好的耐湿耐淹能力,但耐旱性不足且相对于蒲桃和木槿的较弱。
此外,水分胁迫也对不同植物的生理指标具有不同程度的影响,能使植物的光合指标发生适应性变化。其中,净光合速率(Pn)是反映植物适应性的最重要生理指标,而且其强弱直接影响植物的生长发育状况[19],净光合速率显著降低,反映了植物生长发育受到较大抑制[20]。本研究发现,在3种土壤湿度下,除串钱柳在水淹(K2)处理下的Pn最高外,其余6种木本植物的Pn在湿润(K1)处理下最高,而7种植物的Pn均在干旱(K3)处理下最低。蒲桃和木槿在水淹(K2)和干旱(K3)处理下的Pn降幅较大,而在湿润(K1)处理下的Pn降幅较小;其余5种植物则仅在干旱(K3)处理下的Pn降幅较大,而在湿润(K1)和水淹(K2)处理下的Pn降幅较小。因此,水淹(K2)和干旱(K3)对蒲桃和木槿的Pn的抑制较大,而干旱(K3)对红花玉蕊、串钱柳、对叶榕、铁冬青和木芙蓉的Pn的抑制较大,而湿润(K1)对7种植物均无明显抑制。由此推测,根系通气组织的调节作用和根际土壤细菌群落的变化,是使植物的 Pn在湿润(K1)和水淹(K2)处理下减弱程度小于干旱胁迫的原因[21-22];而基因组不同所表达的水淹胁迫响应机制不同,是使耐湿植物的 Pn在水淹胁迫下减弱程度不一的原因[23]。
气孔是植物叶片与外界进行气体交换的主要通道[24],轻度胁迫下,气孔关闭会阻碍CO2进入叶片,胞间CO2浓度(Ci)降低,从而间接影响植物的生长发育状况[25],重度胁迫下,非气孔限制因素则起到了主要作用[26-27]。研究表明,不同水分胁迫下,7种木本植物的气孔导度(Gs)与上述 Pn的变化规律一致,Ci的变化规律则与上述Pn的变化规律相反,说明干旱处理下对7种植物的Pn、Gs、Ci限制较为显著,植物生长发育受到阻碍,这与以往的研究结果一致[28-29]。但试验中,Gs和Ci受到水分胁迫的影响相对于Pn较小,推测非气孔限制因素是植物净光合速率的降低主要原因。
在前人研究的基础上,本试验的光合指标与形态特征的分析结果相当,进一步确定了这7种木本植物的水适应性程度及其适合生存的环境。但也应注意到,仅以小规格苗木为研究对象的短期试验,可能与大规格苗木存在差异性[30],后续应作进一步验证;同时,本试验仅选用了3个光合指标进行测度分析,往后应多挖掘影响木本植物水适应性可能存在的因素,尽可能结合多个指标如光合色素含量[31]、根系活力[32]等进行试验,以期为华南地区可持续湿地景观的构建提供更全面的参考依据。
3.2 结论
综合分析得出,木槿和蒲桃的 Pn、Gs和 Ci在湿润(K1)和干旱(K3)处理下差异不显著,Pn、Gs在湿润(K1)处理下显著高于淹水(K2)和干旱(K3)处理下的,Ci显著低于淹水(K2)和干旱(K3)处理下的,且仅在湿润(K1)处理下长势良好,适合种植在水岸交界处或者水生-湿生交替变化的环境中,如河岸涨落带以及湿地滨水岸边林带;铁冬青、串钱柳、对叶榕、红花玉蕊和木芙蓉的Pn、Gs和Ci在湿润(K1)和淹水(K2)处理下差异不显著,而 Pn、Gs显著高于干旱(K3)处理下的,Ci显著低于干旱(K3)处理下的,且仅在湿润(K1)和淹水(K2)处理下整体生长状况良好,更适合种植在水饱和环境中,如湿地岸际短期浅水淹地带。
