赤松枯梢病生防真菌M75的筛选和鉴定1)
2022-05-16张铭谢宪梁军程元张星耀冯磊时良
张铭 谢宪 梁军 程元 张星耀 冯磊 时良
(中国林业科学研究院,北京,100091) ( 山东昆嵛山森林生态系统国家定位观测研究站)
松枯梢病,又名松梢枯病,是世界范围内在针叶树种上分布最广最常见的重要林木枝干病害之一。该病害的病原存在基因型的分化,在中国引起松枯梢病的病原菌是C型松球壳孢菌(Sphaeropsissapinea)[1]。松枯梢病的寄主有樟子松(Pinussylvestnis)[2]、马尾松(Pinusmassoniana)、火炬松(Pinustaeda)、欧洲赤松(Pinussylvestris)、北美短叶松(Pinusbanksiana)、辐射松(Pinusradiata)[3]、红松(Pinuskoraiensis)[4]等,可侵染8属约60多种针叶树[5]。松枯梢病导致的顶芽坏死、枯梢、枝干溃疡、流脂坏死等症状,对松林造成了严重危害[6]。除上述寄主外,王婧等[7]在油松上也分离得到松枯梢病原菌;谢宪等[8]在进行昆嵛山赤松内生真菌多样性调查时,通过高通量测序发现赤松上也存在松枯梢病原菌侵染,进一步证明了松枯梢病原菌的寄主范围正逐渐扩大。因此,现阶段亟待寻求有效防控松枯梢病的方法。
对于松枯梢病防治的研究,主要集中在营林技术措施及化学防治方面[9]。但目前营林措施还停留在理论阶段,无法为松枯梢病的防治提供实际指导。而长期过量施用化学农药增加了防控成本,长此以往会导致病原菌产生耐药性,甚至引起病害的再度流行[10],严重破坏生态系统的稳定性。近年来,生物防治被越来越多的研究者认为是防治植物病害的最佳策略之一[11]。张丽等[12]提出,微生物防治就是要利用有益微生物,通过微生物之间的竞争、抗生、寄生溶菌作用及诱导抗性等,抑制病原菌的存活和发展。生物防治控制时间长、易于开发,可大幅节约防控成本,其可通过利用拮抗微生物及其产生的次生代谢产物达到病害防控目的,是对环境、资源都更加友好的调控手段。目前,在许多已鉴定的生防微生物中,对生防真菌木霉属(Trichodermaspp.)的研究和应用较多[13]。木霉属真菌是一类普遍存在的土壤真菌,因其对植物病害的有效抑制作用,本身具有广泛的生物利用度及高度的安全性而逐渐被研究者关注。木霉菌作为生防真菌,在对松枯梢病的生物防治方面已有报道。李宝年等[14]在对樟子松枯梢病进行拮抗真菌筛选时,发现绿木霉(Trichodermaviride)对松枯梢病原菌的抑菌效果最好,病原菌被抑制率达70.9%。研究发现,木霉菌在抑制病原的同时,也能促进植物根系生长,从而促进养分吸收[15]。尹大川等[16]发现,引进绿色木霉菌株T43及其代谢产物对松枯梢病具有明显抑制作用。Regliński et al.[17]发现,绿色木霉菌株灌根处理辐射松幼苗,能有效促进其生长,并增强寄主植物对松枯梢病原菌的系统抗性。本研究以松枯梢病原菌为目标病原菌,通过对赤松林地土壤真菌的分离,筛选得到1株具有广谱抑菌活性的森吉木霉菌株M75,为松枯梢病及其他重要林木病原菌的生物防治提供了新的生防菌种资源。
1 材料与方法
1.1 试验材料
在山东省烟台市昆嵛山自然保护区内,随机采集距离赤松主干30 cm范围内的林地土壤,去除土壤表层杂物,采集深度约5~20 cm左右的土壤50 g,共36份,用于拮抗真菌的分离与筛选。分离得到的菌种由中国林业科学研究院森林生态环境与自然保护研究所森林病理学实验室保存。
试验用培养基为马铃薯葡萄糖琼脂培养基(PDA)、马铃薯葡萄糖水培养基(PDB)。
病原菌(Sphaeropsissapinea)由中国林业科学研究院森林生态环境与自然保护研究所森林病理学实验室提供;金黄壳囊孢(Cytosporachrysosperma)、葡萄座腔菌(Botryosphaeriadothidea)、栗疫菌(Cryphonectriaparasitica)、链格孢(Alternriasp.)、围小丛壳菌(Glomerellacingulata)、尖孢镰刀菌(Fusariumoxysporum)、犁头霉(Absidiasp.)、枯斑拟盘多毛菌(Pestalotiopsisfunerea)均由中国林业微生物保藏中心(CFCC)提供。
1.2 研究方法
采用梯度稀释平板涂布法[18]分离土壤微生物。将每份土壤样品取10 g置于装有90 mL灭菌水的250 mL三角瓶中制成土壤悬浮液母液,将母液置于28 ℃摇床中160 r·min-1震荡1 h,静置30 min,在无菌条件下对土壤母液进行10倍梯度稀释,稀释成体积分数为10-2、10-3、10-4的3个梯度,然后依次吸取0.1 mL涂布在PDA平板上,每个梯度做3次重复。将上述平板放入28 ℃恒温培养箱中培养2~3 d。根据形态各异的菌落进行纯化,编号并保存备用。
采用平板对峙法[19]进行拮抗真菌初筛,选取活化5 d的病原菌和拮抗真菌,用打孔器选取5 mm菌饼,分别接种在PDA平板左右两侧,菌饼圆心相距约5 cm,距培养皿边缘约2 cm,菌饼中心的连线经过平板中心。以仅接松球壳孢菌为对照。每株拮抗真菌平行重复3次。接种后的PDA平板倒置于恒温培养箱中28 ℃培养5 d后,测量病原菌菌落指向拮抗菌的生长半径(从菌饼圆心处开始测量)。
被测真菌抑菌率:抑菌率=(病原菌对照菌落半径-处理菌落半径)/病原菌对照菌落半径×100%。
采用菌丝生长速率法[20]进行拮抗真菌复筛。将初步筛选出在平板上具有一定抑菌效果的生防真菌,制备成无菌发酵滤液,进行拮抗真菌的发酵液复筛。将初筛选出的菌株在PDA培养基上活化培养5 d,用打孔器取5 mm的菌饼,接入装有100 mL PDB培养基的250 mL三角瓶中,在28 ℃、180 r·min-1的摇床上振荡培养96 h,得到生防菌的发酵液。收集30 mL发酵液在4 ℃、10 000 r·min-1的条件下离心10 min,收集上清液并使用0.45 μm微孔滤膜对所获上清液过滤除菌,将过滤后的上清液与加热冷却至45~50 ℃的PDA培养基以1∶10的比例混合均匀并倾倒平板,待平板冷却凝固后,在平板中央接种直径5 mm的松枯梢病菌菌饼。以加无菌水制备成的平板接种病原菌作为对照。所有处理进行3次重复。28 ℃恒温倒置培养,5 d后采用十字交叉法测量病原菌菌落的直径大小,计算抑菌率。
通过上述测定,获得具有强抑菌效果的菌株M75,参照《真菌鉴定手册》[21]对该菌进行初步形态学鉴定,并结合Tef1-a序列(EF1-688F(5’-CGG TCA CTT GAT CTA CAA GTG C-3’EF1-1251R(5’-CCT CGA ACT CAC CAG TAC CG-3’)构建系统发育树。DNA扩增和测序体系参考Zhang et al.[22]的方法。PCR产物送至北京华大基因科技股份有限公司经测序后,在NCBI(http://blast.ncbi.nlm.nih.gov/)中BLAST比对分析后,通过MEGA7.0对选取同源性较高的相关菌株序列再次进行比对分析,采用最大似然法构建系统发育树,重复取样1 000次进行自展分析。
采用菌丝生长速率法测定拮抗真菌M75发酵液对8种常见真菌:金黄壳囊孢、葡萄座腔菌、栗疫菌、链格孢、围小丛壳菌、尖孢镰刀菌、犁头霉、枯斑拟盘多毛菌的抑菌活性。
1.3 数据处理
采用Excel 2010(Microsoft,USA)、SPSS 24.0(IBM,USA)软件对试验数据进行统计和分析,并应用Duncan法对不同处理组间的差异进行单因素方差分析(P<0.05)。
2 结果与分析
2.1 拮抗真菌的初筛
将从土壤中分离得到的271株土壤真菌,与松枯梢病原菌进行对峙培养,选取抑菌率前10的菌株,见表1。菌株M75在与松枯梢病原菌对峙培养时,能有效竞争平板内的生长空间,限制病原菌菌丝的生长,使病原菌菌落无法向平板四周延伸(图1B)。在培养5 d后,对照组的病原菌菌落正常生长(图1A),菌落半径为4.51 cm,而在接种菌株M75的平板上,病原菌菌落指向拮抗菌的菌落半径仅为1.23 cm,拮抗真菌M75平板初筛抑菌率为72.65%。
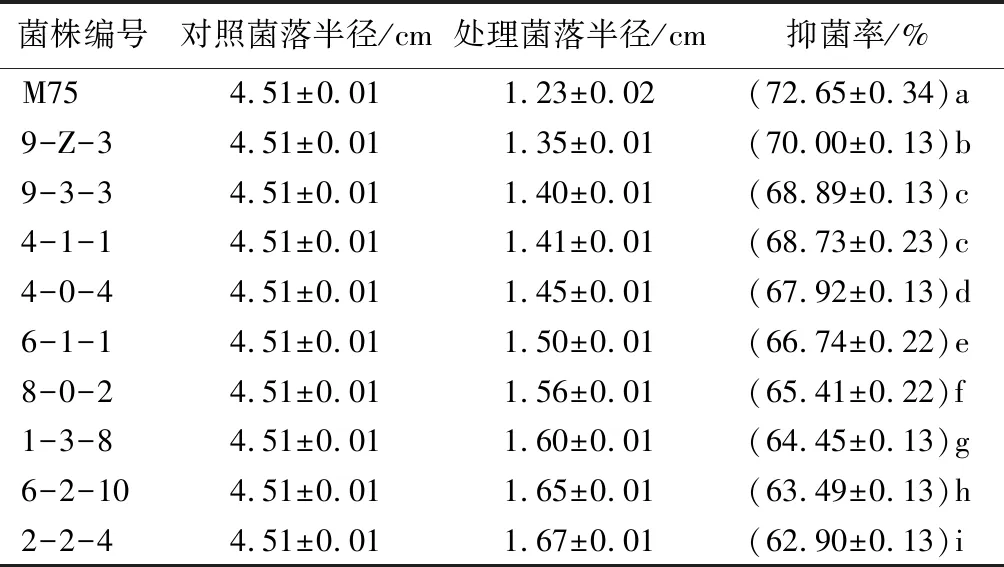
表1 拮抗真菌初筛抑菌率
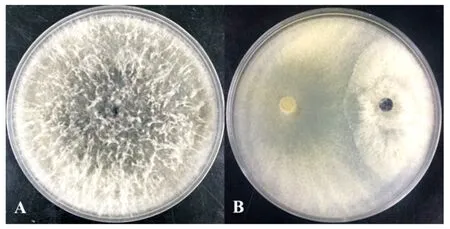
A为对照;B为处理(B中左侧为M75菌株,右侧为松枯梢病原菌)。
2.2 拮抗真菌的复筛
采用PDB培养基对菌株M75进行发酵处理,获得无菌发酵滤液。采用十字交叉法测量病原菌在添加了M75无菌发酵滤液后平板上的菌落直径(表2、图2)。病原菌在含有M75发酵滤液的培养基上菌落生长速度缓慢,菌落生长受到明显抑制。培养5 d后利用生长速率法计算发酵液复筛抑菌率,其复筛抑菌率为78.78%。
2.3 拮抗真菌M75形态、培养特征
在PDA培养基上,M75菌株菌落呈圆形,菌落初呈白色,轮廓清晰,气生菌丝浓密,初期为白色,绒毛絮状,致密,后逐渐变为浅黄色。28 ℃培养3 d菌落半径为45~55 mm,5~7 d长满平板,生长速率为6.7~7.5 mm·d-1。菌落不产生色素,但有椰子味气体产生。菌株M75的形态学特征与Park et al.[23]发现的森吉木霉新种SFC20130926-S001及秦文韬等[24]发现的森吉木霉中国新纪录种HMAS 248793基本一致。
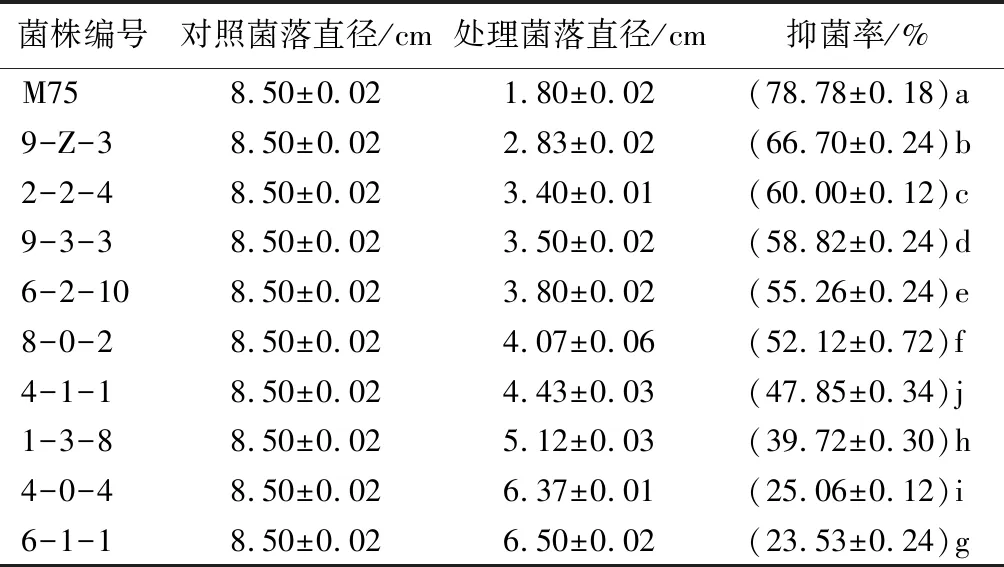
表2 拮抗真菌发酵液复筛抑菌率
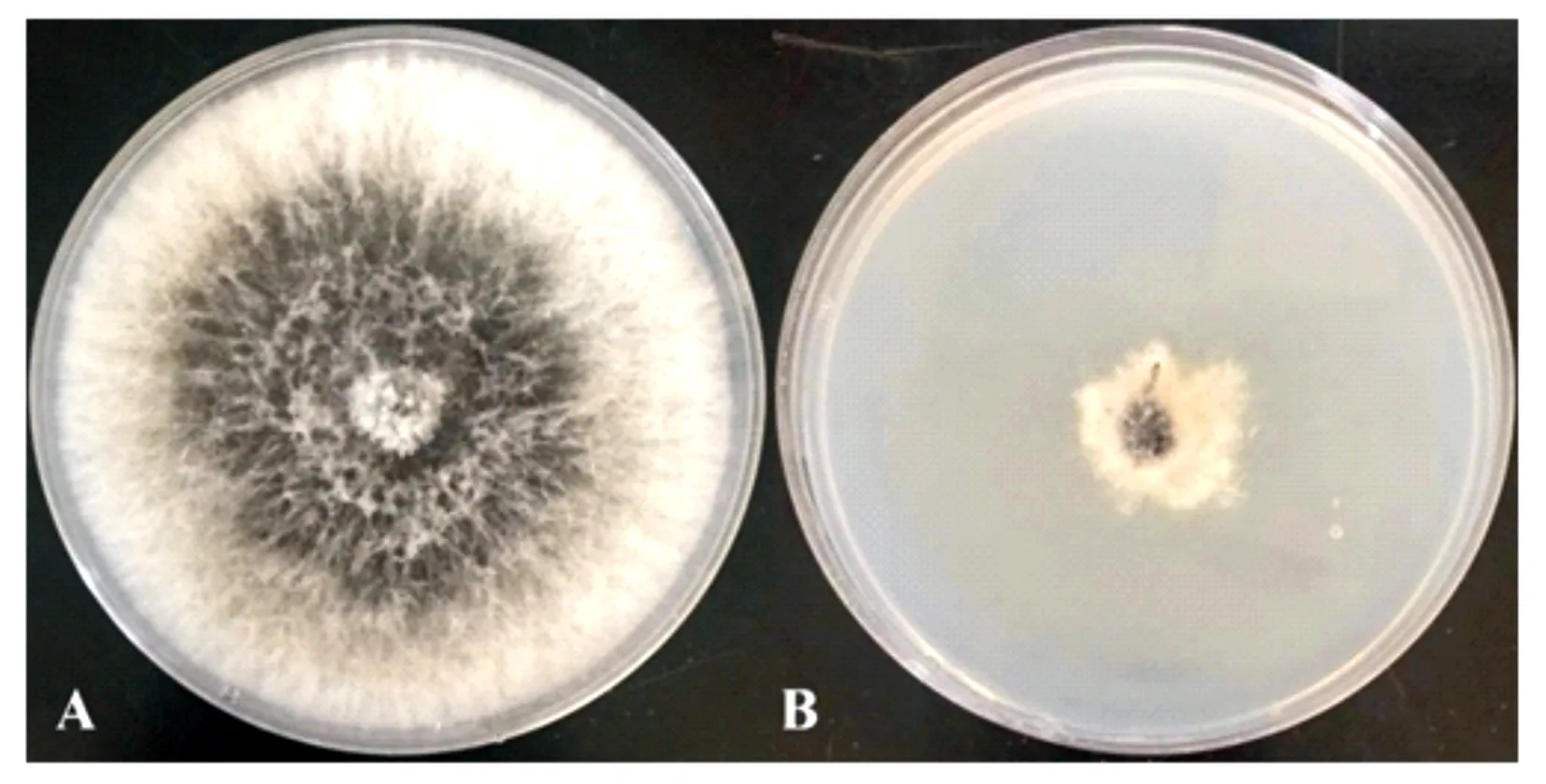
A为对照;B为处理。
图3 M75在PDA上生长14 d的菌落
2.4 拮抗真菌M75的Tef1-a序列分析
拮抗真菌M75的Tef1-a基因PCR产物经测序后,在NCBI(http://blast.ncbi.nlm.nih.gov/)中进行BLAST比对分析,发现其与森吉木霉(Trichodermasongyi)TC556最为接近,相似度达98%。选取同源性较高的序列与M75构建最大似然树,如图4所示,发现在系统发育树上菌株M75与TrichodermasongyiTC556聚在1个分支上,支持率为96%,因此将菌株M75鉴定为森吉木霉CFCC54490。

括号内容为菌株在Genbank中的登录号;Bootstrap次数设置为1 000,最大似然法构建系统发育树;分支处的数字为Bootstrap支持率;标尺为每个核苷酸位点上的替换值。
2.5 拮抗真菌M75发酵滤液抑菌谱测定
采用菌丝生长速率法测定生防菌株M75对金黄壳囊孢等8种常见真菌的抑菌活性见表3、图5。结果显示,M75对金黄壳囊孢的抑菌率最高,达94.05%;对葡萄座腔菌、栗疫菌的抑菌效果较好,抑菌率分别为82.30%、77.87%;对链格孢的抑菌效果一般,抑菌率为52.56%;对尖孢镰刀菌、围小丛壳菌、犁头霉、枯斑拟盘多毛菌也具有一定的抑菌性,但抑菌效果不高,抑菌率分别为36.35%、29.91%、28.72%、23.53%。
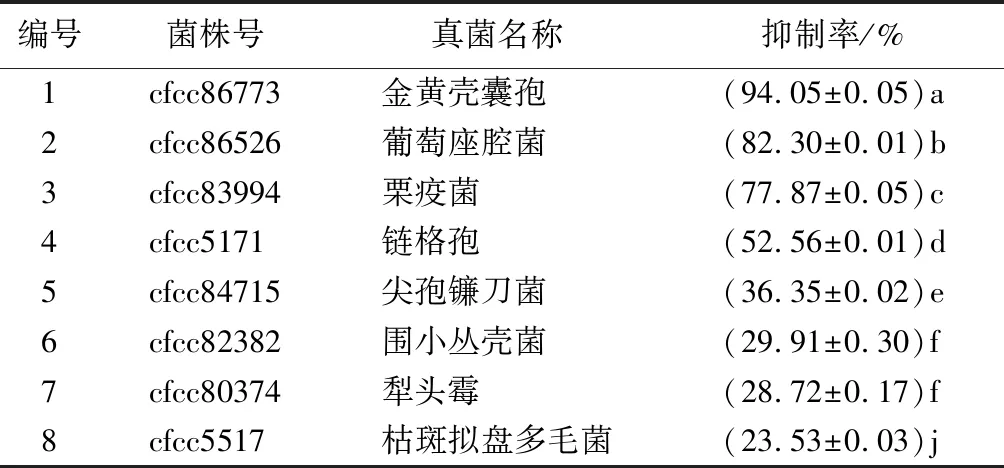
表3 菌株M75发酵液对8种常见真菌的抑制率
3 结论与讨论
松枯梢病是由松球壳孢菌引起的一种世界性的针叶树种病害,因其导致的枯梢、树干溃疡、流脂、根茎腐烂、木材蓝变等问题,对松树人工林造成了严重的危害和损失,因此亟需寻求一种有效的防控手段。近年来,随着生物防治理念的兴起,利用微生物菌种资源防控植物病害已成为病害防控发展的新趋势。木霉属真菌作为一类具有广泛应用前景的生防真菌,越来越受国内外学者的重点关注。木霉菌能通过与寄主竞争营养和空间、诱导植物抗性、产生抗生素等次级代谢产物方面抑制植物病原菌的发生与发展,同时能促进植物的生长发育及对营养的利用[25]。
本研究从分离自赤松林地土壤的271株真菌中,筛选得到1株对松枯梢病原菌具有显著拮抗作用的土壤真菌M75,结合形态学特征及Tef1-a分子序列分析将其鉴定为森吉木霉CFCC54490。该菌株对松枯梢病原菌具有较好的抑菌作用,其平板对峙抑菌率达到72.65%,发酵滤液原液的抑菌率则达到78.78%,原因是在液体培养状态下,促进了M75菌株产生更多的次级代谢产物,从而提高了抑菌效果。试验发现,菌株M75不只对松枯梢病原菌具有抑制作用,对其他多种重要的林木病原真菌均具有一定的抑菌效果。M75发酵液对引起杨树腐烂病的病原菌金黄壳囊孢抑菌率高达94.05%;对引起林木溃疡病的葡萄座腔菌抑菌率为82.30%;对引起板栗疫病的栗疫菌抑菌率为77.87%,抑菌谱较广,具有较大的生防潜力,可作为1株优质的生防菌株开展后续研究。同时,这也是木霉属除绿色木霉、哈茨木霉(Trichodermaharzianum)、棘孢木霉(Trichodermaasperellum)等常见的生防木霉以外[26],首次发现森吉木霉也可应用于生物防治领域,扩充了生防木霉菌微生物资源库,为森吉木霉菌株M75的进一步开发利用提供了参考。但微生物防治在实际应用过程中,会受到外界因素的影响,因此还需进一步探究不同培养条件下森吉木霉M75的抑菌效果,另外,对其抑菌机理还需进一步深入探究。
