海拔对入侵植物粗毛牛膝菊种群繁殖力与种子扩散力的影响
2022-05-14陈晓艳张文刚刘芮伶刘刚
陈晓艳, 张文刚, 刘芮伶, 刘刚
海拔对入侵植物粗毛牛膝菊种群繁殖力与种子扩散力的影响
陈晓艳, 张文刚, 刘芮伶, 刘刚*
陕西师范大学生命科学学院, 西安 710119
外来入侵植物在高大山体中沿着海拔扩散的过程中, 由于环境选择压力的改变, 其繁殖力和种子扩散力可能会发生改变。为探究入侵植物粗毛牛膝菊()在秦巴山区扩散过程中的繁殖力和种子传播力的变化趋势, 通过野外调查对比分析了其位于不同海拔梯度(505—1947 m)上的种群密度、盖度、单株种子产量和种子羽流载荷(MAR)等的差异, 并对比分析了不同生境类型中的粗毛牛膝菊种群繁殖特性上的差异。研究结果显示: (1)粗毛牛膝菊的种群盖度和密度随海拔升高均未表现出显著的变化趋势; (2)单个花序成熟种子数随海拔升高而增加, 但单株花序数、单株种子产量(数量和重量)均随海拔升高而降低; (3)种子MAR随海拔升高而显著升高, 即种子传播力呈显著下降趋势; (4)相较于荒地生境中的种群, 农田中的粗毛牛膝菊种群盖度和单株种子重量更高, 且MAR更小。研究结果说明随着海拔升高, 粗毛牛膝菊繁殖力和种子传播力均呈显著下降趋势, 农田中的粗毛牛膝菊种群繁殖力和种子传播力均显著高于荒地中的种群, 这暗示了秦巴山地在降低该物种的入侵扩散速度方面具有重要意义, 但农业生产活动带来的人为干扰可能会促进该种的入侵与扩散进程。
入侵植物; 粗毛牛膝菊; 秦巴山区; 种子扩散力; 海拔梯度
0 前言
繁殖体压力假说认为, 外来种在引入地的个体数量越多, 繁殖体压力越大, 其在该地的入侵潜力就越大[1, 2]。通常认为, 当一个地区受干扰程度越高时, 那么外来种入侵该地所需要的繁殖体压力就越小[3]。在山地生态系统中, 人类活动对低海拔地区的干扰较大, 且外来种繁殖体被引入的机率更高、繁殖体压力更大, 因而低海拔地区容易成为外来植物入侵的第一站[4-7]。随着海拔升高, 温度、湿度、光照等诸多环境因子都在发生较大的变化, 这导致了山地生态系统具有环境异质性高的特点, 且这种环境异质性会极大地影响植物的生长发育、甚至导致部分器官组织的结构与功能发生变化[8]。此外, 随着海拔升高, 入侵种繁殖体压力可能会降低, 能够成功入侵到高海拔地区的物种需要更加良好的自身条件[9]。表型可塑性理论认为, 入侵种之所以能够迅速占据不同生境, 是因为其具有较高的表型可塑性, 能够使其对扩散过程中遇到的不同环境条件快速地做出适应性的调整[10]。但当前已有的研究尚未明确, 在外来入侵植物随着海拔向上扩散的过程中, 影响其扩散潜力的种子形态结构、传播力和繁殖体压力是否会发生改变, 以及会如何改变。越来越多的研究认为, 随着人类干扰和全球气候变化的加剧, 外来种向高海拔地区扩散的趋势可能会逐步增强[11, 12]。因此, 随着越来越多的外来入侵种持续向高海拔地区扩散, 可能对高海拔地区的生态系统造成巨大的负面影响。因此, 针对入侵种向高海拔山地扩散的研究亟待得到足够重视。
菊科植物不仅是被子植物中最大的科而且也是我国种子植物中最大的科[13, 14]。但我国现有的菊科植物很多都是外来种, 菊科的入侵种占我国外来入侵植物的22%左右[15, 16]。较高的种子传播能力是很多菊科植物之所以能够迅速扩散并占领新生境的关键所在[15]。植物传播种子的方式多种多样, 其中, 种子风媒传播是入侵植物种群扩散传播最为常见的方式[17, 18]。菊科植物的花萼常变态为冠毛, 冠毛的存在使得很多菊科植物的种子能够依靠风力进行远距离传播扩散。除了适合远距离传播的种子结构, 大多数菊科植物还有很高的种子产量[19], 由此而造成的高繁殖体压力正是外来种成功入侵的首要条件。粗毛牛膝菊()为菊科牛膝菊属一年生植物。其原产地位于中、南美洲[20], 现已广泛入侵至世界上大部分地区。近几年来该种在我国呈爆发式生长[21, 22]。粗毛牛膝菊主要出现在诸如农田、路边等受到过人为活动干扰的生境中, 是一种重要的农业入侵杂草[23, 24]。目前该种已经从东南逐步扩展进入秦岭和巴山地区[22]。秦巴山区(102°24'—112°40'E和30°43'—35°29'N)西接青藏高原东缘, 东临华北平原, 汉江以北为秦岭, 汉江以南为巴山, 中部分布有汉中盆地和安康盆地[23]。秦巴山区山体高大, 涵盖面积广, 由南向北随着纬度升高和由低到高随着海拔升高, 气候类型逐步由北亚热带气候向温带气候过渡[24, 25]。北侧的秦岭是我国生物、地理和气候的南北分界线, 该地区生物多样性极其丰富, 在我国生物多样性保护工作中处于无比重要的地位[21]。随着粗毛牛膝菊在秦巴山区的持续扩散, 尤其是向高海拔地区的持续进行, 其有可能能够成功翻越秦岭进入北部的关中平原, 并威胁到当地的农业生态系统。本研究旨在通过比较分析秦巴山区不同海拔梯度上的粗毛牛膝菊种群在繁殖和种子扩散能力方面是否出现分化, 以及会如何变化, 以评估其向秦岭和巴山高海拔地区扩散的潜力, 为揭示该外来种的入侵机制和制定防控措施提供理论基础。
1 研究方法
1.1 野外调查
于2018年8月在秦巴山区沿宁陕县—石泉县—汉阴县—安康县—岚皋县—平利县一线开展粗毛牛膝菊种群野外调查(图1)。随着海拔变化设置了18个调查样带(表1)。在每个调查样带内随机选取粗毛牛膝菊入侵的群落, 设置至少3个1 m×1 m的调查样方, 统计样方内粗毛牛膝菊种群盖度(%)和种群密度(株·m-2)等。在每个调查样带随机选取20株成熟的粗毛牛膝菊植株, 统计株高(cm)、单株花序数, 并随机选取20个成熟的花序, 统计每个花序内成熟种子数量, 并收集相应的种子置于封口袋内带回实验室备用。记录样方的海拔(m)、经纬度和土地利用类型等信息。
随机选取每个样带内采集到的粗毛牛膝菊种子至少10组, 在实验室内测定种子的百粒重(g), 并根据每个花序种子数量和种子百粒重的平均值计算出每个样带内的单株种子数量(粒)和单株种子重量(g)。利用种子分析系统WinSEEDLE Pro(WinSEEDLE™, Régent Instruments Inc., Québec, QC, Canada)测量种子的长度、冠毛幅度和冠毛长度(PapL)等。
采用羽流载荷(mass-area ratio, 以下简称MAR)这一常见的形态学特征来评估种子的扩散能力。羽流载荷是菊科植物风媒种子扩散能力的可靠指标, 它是由种子的质量面积比来量化的。MAR的计算方法如下[26]:

其中是种子重量,是种子冠毛半径。越低, 种子的扩散潜力越大。
1.2 数据分析
采用线性回归分析路线调查中粗毛牛膝菊种群盖度、种群密度、株高、花序数量和单个花序种子数量以及百粒重与海拔间的关系, 以及各形态指标之间的权衡关系。
采用非配对的t检验对比分析农田和荒地生境中的粗毛牛膝菊种群盖度、种子羽流载荷、单株种子数量和单株种子重量的差异。所有数据分析在R 3.5.3中完成。采用DIVA-GIS软件绘制样点分布图。
2 结果与分析
2.1 种群密度和盖度
线性回归分析结果显示, 粗毛牛膝菊种群盖度(图2a;= -0.11,= 0.42)、种群密度(图2b;= 0.14,= 0.31)随海拔均未发生显著变化。但粗毛牛膝菊植株的株高随海拔升高显著降低(图3;= -0.22,<0.001)。这表明, 随着海拔的升高, 同样大小的样地中分布的粗毛牛膝菊数量相差不大但植株个体变小。

图1 野外定位监测研究和路线调查研究涵盖区域所在地理位置
Figure 1 The location of the field experiment and sampling survey

表1 野外调查的18个种群的地理信息

图2 粗毛牛膝菊种群盖度(a)和密度(b)随着海拔的变化
Figure 2 The variation of coverage(a)and density(b)ofpopulation along elevation
2.2 种子产量
线性回归分析结果显示, 粗毛牛膝菊单株花序数与海拔显著负相关(图4a;= -0.16,= 0.0026),单个花序成熟种子数与海拔显著正相关(图4b;= 0.23,= 0.00013), 而单株种子数量和重量均与海拔显著负相关(图5; 数量:= -0.11,= 0.039; 重量:= -0.23,<0.001)。即, 虽然随着海拔的升高粗毛牛膝菊单个花序中的成熟种子数增加, 但由于单株花序数的降低, 粗毛牛膝菊单株的种子数量和重量都是减少的。

图3 粗毛牛膝菊植株高度随着海拔的变化
Figure 3 The variation of height ofalong elevation
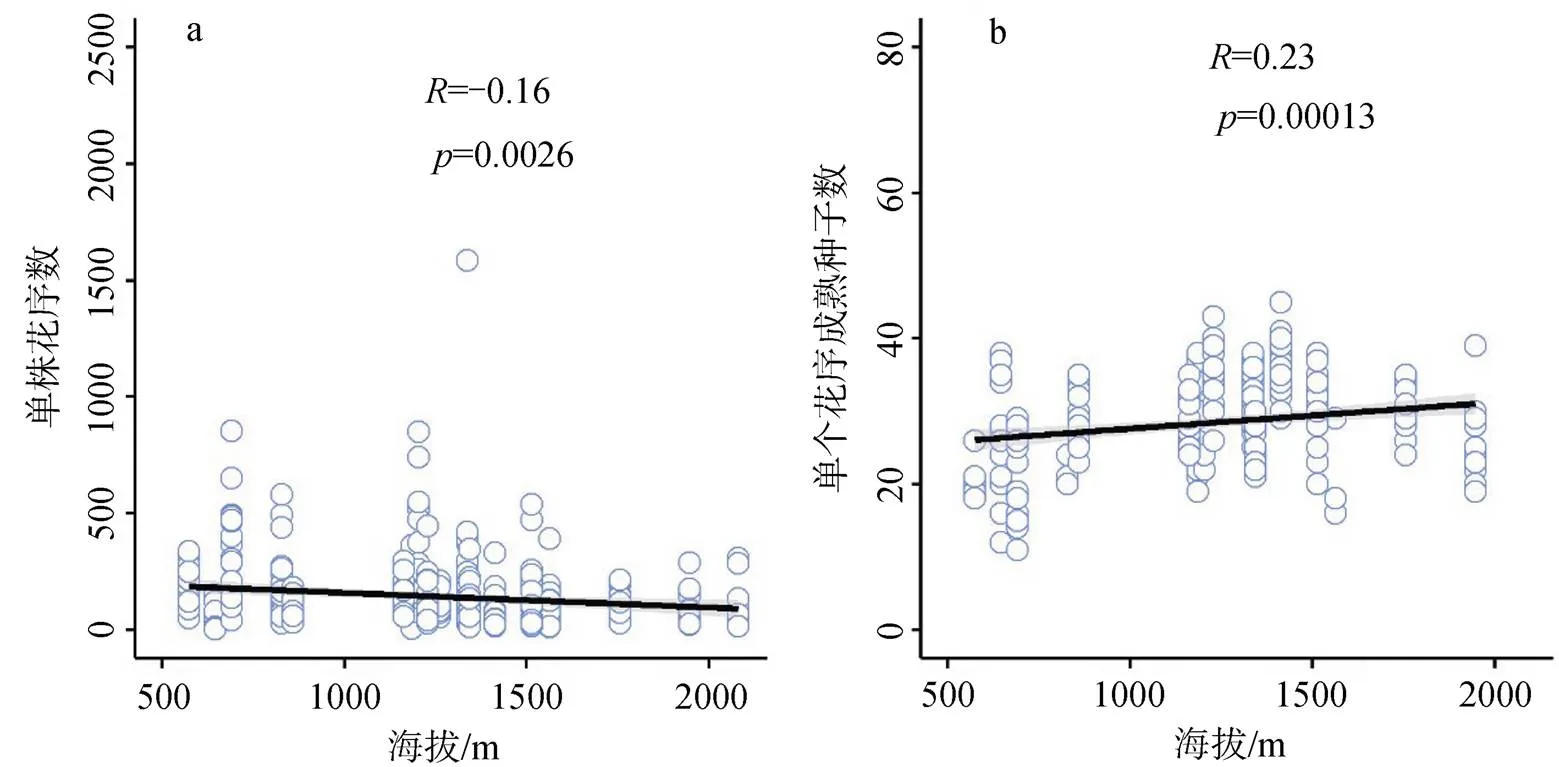
图4 粗毛牛膝菊单株花序数(a)和单个花序成熟种子数(b)随着海拔的变化
Figure 4 The variation of No. of inflorescence per plant (a) and No. of seeds per inflorescence (b) ofalong elevation

图5 粗毛牛膝菊单株种子数量(a)和重量(b)随着海拔的变化
Figure 5 The variation of seed quantity per plant (a) and seed mass per plant (b) ofindividual along elevation
2.3 种子传播力
线性回归分析结果显示, 粗毛牛膝菊种子百粒重与海拔显著正相关(图6c;= 0.65,<0.001), 种子冠毛长度与海拔显著负相关(图6b;= -0.3,<0.001)。即, 随着海拔升高, 粗毛牛膝菊单粒种子重量在增加, 但冠毛长度在减小。种子羽流载荷(MAR)是由种子的质量面积比来量化的, 所以粗毛牛膝菊种子羽流载荷随着海拔升高显著升高(图6a;=0.43,<0.001)。因为种子的羽流载荷越高, 其扩散潜力就越小。所以, 粗毛牛膝菊种子的传播力随海拔升高而降低。
2.4 不同土地利用类型的对比
对比不同生境中的粗毛牛膝菊种群的生长与繁殖特性, 结果显示, 农田中的粗毛牛膝菊种群盖度更高, 种子羽流载荷更小, 单株种子数量无明显差异, 但单株种子重量更高(图7)。

图6 粗毛牛膝菊种子百粒重、冠毛长度、MAR随着海拔的变化
Figure 6 The variation of hundred seeds weigh, PapL, mass-area ratio(MAR) ofalong elevation
3 讨论
3.1 粗毛牛膝菊繁殖体压力随着海拔升高的变化趋势及可能的原因
从外来种自身的生物学特性来看, 物种的生殖方式和繁殖能力的大小对繁殖体压力会产生重要影响[3]。一般认为繁殖体压力的两个重要组成部分是引入繁殖体的数量和大小, 但繁殖体质量的差异也不容忽略[3, 27]。而且繁殖体压力与环境因子、生物和非生物抗性等共同作用[1-3]。粗毛牛膝菊种子的特征符合典型菊科植物种子特征, 因此其种子具有远距离扩散的特性。本研究中, 随着海拔升高, 粗毛牛膝菊种子的羽流载荷升高、单株种子的数量与重量降低, 所以其种群繁殖体压力随海拔升高呈下降趋势。此外, 生境干扰强度越大时, 入侵该生境所需的繁殖体压力就越小[3]。农田生态系统的干扰强度比荒地的要大, 所以入侵农田所需的繁殖体压力较小。对比这两种不同生境中的粗毛牛膝菊的繁殖特性, 我们发现农田中的粗毛牛膝菊种群繁殖体压力更大, 这暗示农田中的粗毛牛膝菊种群可能更容易定植和扩散, 可能会对农业系统造成较大破坏。
植物的繁殖力与营养生长之间也存在一种权衡关系, 有研究表明随海拔梯度的升高植物的繁殖分配也逐渐增加[28]。在本研究中随海拔梯度的增加, 粗毛牛膝菊种群密度和盖度随着海拔的升高没有产生显著变化, 但植株高度显著降低, 即种群中的植株数量没有显著变化但植株个体变小。这表明粗毛牛膝菊种群在繁殖力与营养生长之间也存在权衡, 随着海拔升高其繁殖分配增加。主要原因可能是随着海拔高度的升高, 环境条件越来越恶劣, 输送给植物的可利用能量可能随之减少, 植物的生存和繁殖都受到威胁, 那么尽可能分配较少的能量给支持结构(营养器官)而提高繁殖分配, 才能保证亲代本身的存活和后代的延续[29]。

图7 粗毛牛膝菊在不同土地利用类型中种群盖度、种子羽流载荷、单株种子数量和单株种子重量对比
Figure 7 Comparisons on the traits (coverage, MAR, seed production per plant and seed mass per plant) ofamong different land use types
胁迫梯度假说(stress-gradient hypothesis)认为, 对于一年生入侵植物来说, 低纬度地区更有利的环境条件可能会导致共生物种之间更激烈的竞争, 有可能推动对高营养生物量投资的选择, 而高纬度地区恶劣的气候条件和相关的繁殖不确定性可能会减少入侵植物对营养生物量的投资, 而增加对高生殖投资的选择[30, 31]。这些驱动因素结合在一起, 可能导致随着纬度的增加入侵植物生殖分配的增加或恒定[30, 31]。本研究的结果暗示随着海拔的升高入侵植物在营养生物量和生殖投资之间可能也存在类似的规律。
3.2 粗毛牛膝菊种子扩散力随着海拔升高的变化趋势及可能的原因
本研究结果表明, 粗毛牛膝菊种子产量和扩散力均随其种群所处的海拔梯度的升高而呈显著降低趋势。种子扩散力的这种改变是适应海拔梯度上不同的环境因子的结果。已有的研究表明, 种子扩散力往往会随着环境梯度发生改变。例如, 纬度梯度能够引起降水、温度、热量等环境因子的变化进而导致入侵种的分布范围、群落结构、入侵效应等特征发生显著变化[32, 33]。入侵种的扩散方式和扩散速度是影响外来植物建立种群后扩散传播的重要因素[10]。有研究表明, 在扩散方式方面, 纬度越高, 入侵种种子越倾向于风力传播[34]。随着纬度梯度的变化, 植物的扩散能力也可能会发生变化。入侵杂草在美国草原从低纬度向高纬度区域的扩散过程中, 首先作为能够抑制本土植物的优势种快速生长建群, 随着环境压力的增大, 入侵杂草作为先锋物种改善生境进而提高本土植物多样性[35]。即, 随着纬度的升高入侵种的扩散能力是下降的。
海拔梯度上温度、降水等环境因子的变化与纬度梯度有很多相似之处, 但与纬度梯度不同的是, 季节和日长在海拔梯度上变化不大[36, 37]。在全球范围内, 外来种沿海拔梯度的入侵受到繁殖体有效性、环境因子、进化和生物相互作用的影响[11]。在本研究中, 粗毛牛膝菊种子数量与海拔负相关, 种子重量与海拔正相关。所以, 随着海拔的升高, 粗毛牛膝菊趋向于产生数量更少, 但单粒重量更大的种子。而较大的种子一般可以产生具有较大竞争优势的幼苗[38], 所以粗毛牛膝菊种子特性的这种变化暗示, 随着海拔的升高, 粗毛牛膝菊的繁殖体压力和传播力都降低。这可能是粗毛牛膝菊为了适应不同的生境, 利用其自身的表型可塑性对形态特征和生理特性方面进行的一种调节[10]。表型可塑性是植物适应异质生境的主要对策之一, 它被认为是植物在可变环境中最大适合度的功能反应[39, 40]。很多入侵植物都具有较强的表型可塑性, 如表型可塑性赋予喜旱莲子草对不同水陆生境的广泛适应性和对污染环境很强的耐受性[39]。在本研究中, 粗毛牛膝菊的这种表型可塑性使得它可以适应不同海拔地区的生境。
从根本上来说, 入侵植物的这种表型可塑性其实是植物功能性状之间的一种权衡关系的外在表现[40, 41]。概括地讲, 权衡泛指性状间的一切反比关系, 植物存在很多类似的权衡关系, 主要包括叶片性状间、叶片与枝条及树干性状间、繁殖性状与数量间、繁殖性状与幼苗叶片间、叶片与根性状间的权衡[40, 41]。随着海拔高度的升高, 环境条件越来越恶劣, 输送给植物的可利用能量可能随之减少, 植物在两对具有权衡关系的性状之间必须进行取舍。有研究表明随着海拔升高, 植物可能分配较少能量给支持结构(营养器官), 而较多能量用于生殖来完成生活史[29]。在本研究中, 粗毛牛膝菊为了适应高海拔地区恶劣的生存环境, 在种子存活率和种子扩散力的权衡中选择了更为稳妥的策略, 分配更多的能量用来提高种子的存活率。从表型上来看, 就是种子数量减少、单粒种子重量增加、种子的传播力下降。此外, 在不同生境中的粗毛牛膝菊种群的生长与繁殖特性的对比中, 我们发现农田中的粗毛牛膝菊种群盖度更高, 种群羽流载荷更小, 单株种子数量无明显差异, 但单株种子重量更高。即农田中的粗毛牛膝菊分布更多, 产生的繁殖体压力更大, 而且种子传播力更强。农田生境因受到人为干扰较大, 其区别于非农田生境的地方主要表现在营养物质可利用性水平更高和竞争者种类更少等方面。已有的研究暗示了人为干扰导致的土壤营养升高可能会减少入侵植物在营养分配上的权衡, 进一步增强入侵植物的竞争力和扩散力, 如氮的升高会增强粗毛牛膝菊在种间竞争中的优势[22]。
3.3 高大山体对入侵植物扩散的阻挡效应
本研究结果说明了随着海拔升高, 粗毛牛膝菊繁殖力和种子传播力均显著下降, 且农田中的粗毛牛膝菊种群繁殖力和种子传播力均显著高于荒地中的种群, 这暗示了秦岭和巴山在降低该种的入侵扩散速度方面具有重要意义。山地对于外来植物而言可能是一种有效的入侵屏障[42]。如前文所述, 随着海拔升高, 温度、湿度、光照等诸多环境因子都会发生较大的变化[8, 36, 37], 在外来入侵植物随着海拔梯度向上扩散的过程中, 影响其繁殖和扩散的种子形态结构、传播力和繁殖体压力等关键性质也因响应这种环境因子的变化而会发生改变[11]。为了在高海拔地区成功定植, 外来植物必须克服多重阻碍, 即生态过滤[43]。种子可用性是初始过滤, 即扩散限制, 随后是包括非生物条件和生物相互作用在内的其他生态过滤, 生态过滤决定外来植物个体是否能够成功建立种群[43]。许多假设认为这些非生物条件的生态过滤使高山生态系统具有抵抗外来植物入侵的能力[42]。已有研究表明, 海拔高度是许多外来生物入侵的重要限制因子, 随着海拔的上升, 外来入侵植物种类和数量都相对减少, 危害程度也减小[44-47]。山体的阻挡对外来物种扩散的限制具有重要意义。
但越来越多的研究认为, 随着人类干扰和全球气候变化的加剧, 外来种向高海拔地区扩散的趋势可能会逐步增强[11, 12], 生物入侵对高海拔地区生态环境安全的威胁不容忽视。本研究也显示, 农田中的粗毛牛膝菊种群繁殖力与种子传播力均显著高于荒地, 这可能是因为人为的农业生产活动对该种的入侵与扩散具有一定的促进作用。秦岭和巴山所处的地理位置特殊, 西接青藏高原东缘, 东临华北平原, 是中国大陆青藏高原以东最高山体。秦巴山区是我国生物、地理和气候的南北分界线, 该地区生物多样性极其丰富, 在我国生物多样性保护工作中处于无比重要的地位, 尤其对阻挡来自东南的外来入侵种具有重要意义[21-25]。秦岭和巴山地区所面临的诸多生态问题需要相关研究人员更多的关注, 对于可能发生的生物入侵应该及时采取合适的防治措施。
[1] HOVICK S M, WHITNEY K D. Propagule pressure and genetic diversity enhance colonization by a ruderal species: a multi-generation field experiment[J]. Ecological Monographs 2019, 89(3): 1–22.
[2] MOYANO J, CHIUFFO M C, POLICELLI N, et al. The interplay between propagule pressure, seed predation and ectomycorrhizal fungi in plant invasion[J]. NeoBiota, 2019, 42: 45–58.
[3] 陈中义, 江红英. 繁殖体压力—一种解释生物入侵的机制[J]. 长江大学学报(自科版), 2008, 5(4): 79–81.
[4] DIETZ H, EDWARDS P J. Recognition that causal processes change during plant invasion helps explain conflicts in evidence[J]. Ecology, 2006, 87(6): 1359–1367.
[5] KHUROO A A, RESHI Z, RASHID I, et al. Plant invasions in montane ecosystems[J]. Frontiers in Ecology and the Environment, 2009, 7(8): 408–408.
[6] NOGUES-BRAVO D, ARAUJO M B, ROMDAL T, et al. Scale effects and human impact on the elevational species richness gradients[J]. Nature, 2008, 453(7192): 216–220.
[7] WILSON J R U, DORMONTT E E, PRENTIS P J, et al. Something in the way you move: dispersal pathways affect invasion success[J]. Trends in Ecology and Evolution, 2009, 24(3): 136–144.
[8] 尹鹏, 黄娇, 廖金花, 等. 峨眉山不同海拔迎阳报春花的生理特性[J]. 暨南大学学报(自然科学与医学版), 2019, 40(6): 550–555.
[9] ALEXANDER J M, KUEFFER C, DAEHLER C C, et al. Assembly of nonnative floras along elevational gradients explained by directional ecological filtering[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(2): 656–661.
[10] 宫伟娜, 万方浩, 谢丙炎, 等. 表型可塑性与外来入侵植物的适应性[J]. 植物保护, 2009, 35(4): 1–7.
[11] ALEXANDER J M, LEMBRECHTS J J, CAVIERES L A, et al. Plant invasions into mountains and alpine ecosystems: current status and future challenges[J]. Alpine Botany, 2016, 126(2): 89–103.
[12] YINGBO Y, GANG L, XIONG S, et al. Where will Invasive plants colonize in response to climate change: predicting the invasion ofin China[J]. International Journal of Environmental Research, 2018, 12(6): 929–938.
[13] 朱世新, 覃海宁, 陈艺林. 中国菊科植物外来种概述[J]. 广西植物, 2005, 25(1): 69–76.
[14] 徐跃良, 鲁益飞. 中国菊科植物拾零[J]. 浙江大学学报(理学版), 2019, 46(2): 73–78.
[15] 诸葛晓龙, 朱敏, 季璐, 等. 入侵杂草小飞蓬和钻形紫菀种子风传扩散生物学特性研究[J]. 农业环境科学学报, 2011, 30(10): 1978–1984.
[16] WEBER E, SUN Shiguo, LI Bo, et al. Invasive alien plants in China: diversity and ecological insights[J]. Biological Invasions, 2008, 10(8): 1411–1429.
[17] HOLLE B V, SIMBERLOFF D. Ecological resistance to biological invasion overwhelmed by propagule pressure[J]. Ecology, 2005, 86(12): 3212–3218.
[18] BUORO M, CARLSON S M. Life-history syndromes: integrating dispersal through space and time[J]. Ecology Letters, 2014, 17(6): 756–767.
[19] 周俗, 唐川江, 张新跃. 四川省紫茎泽兰危害状况与治理对策[J]. 草业科学, 2004, 21(1): 24–26.
[20] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 1973.
[21] 刘刚, 张璐璐, 孔彬彬, 等. 外来种粗毛牛膝菊在秦巴山区的种群发展动态[J]. 生态学报, 2016, 36(11): 3350– 3361.
[22] LIU Gang, YANG Yingbo, ZHU Zhihong. Elevated nitrogen allows the weak invasive plantto become more vigorous with respect to inter-specific competition[J]. Scientific Reports, 2018, 8(1): 31–36.
[23] 陈超男, 朱连奇, 田莉, 等. 秦巴山区植被覆盖变化及气候因子驱动分析[J]. 生态学报, 2019, 39(9): 3257-3266.
[24] 陆福志, 鹿化煜. 秦岭—大巴山高分辨率气温和降水格点数据集的建立及其对区域气候的指示[J]. 地理学报, 2019, 74(5): 875–888.
[25] 林霆. 秦岭是一座怎样的山?[J]. 西部大开发, 2019, 255(5): 144–149.
[26] MATLACK G R. Diaspore size, shape, and fall behavior in wind-dispersed plant species[J]. American Journal of Botany, 1987, 74(8): 1150–1160.
[27] KOONTZ M J, OLDFATHER M F, MELBOURNE B A, et al. Parsing propagule pressure: number, not size, of introductions drives colonization success in a novel environment[J]. Ecology and Evolution, 2018, 8(16): 8043–8054.
[28] ZHAO Fang, YANG Yongping. Reproductive allocation in a dioecious perennial() along altitudinal gradients[J]. Journal of Systematics and Evolution, 2008, 46(6): 830–835.
[29] 张林静, 石云霞, 潘晓玲. 草本植物繁殖分配与海拔高度的相关分析[J]. 西北大学学报(自然科学版), 2007, 37(1): 77–80.
[30] MALKINSON D, TIELBöRGER K. What does the stress-gradient hypothesis predict? Resolving the discrepancies[J]. Oikos, 2010, 119(10): 1546–1552.
[31] HELSEN K, ACHARYA K P, GRAAE B J, et al. Earlier onset of flowering and increased reproductive allocation of an annual invasive plant in the north of its novel range[J]. Annals of Botany, 2020, 126(6): 1005–1016.
[32] MARON J L. The relative importance of latitude matching and propagule pressure in the colonization success of an invasive forb[J]. Ecography, 2006, 29(6): 819–826.
[33] 冯建孟, 董晓东, 徐成东. 中国外来入侵植物区系成分的聚类分析和排序[J]. 大理学院学报, 2009, 8(4): 58–63.
[34] BUTLER D W, GREEN R J, LAMB D, et al. Biogeography of seed-dispersal syndromes, life-forms and seed sizes among woody rain-forest plants in Australia's subtropics[J]. Journal of Biogeography, 2007, 34(10): 1736–1750.
[35] RICHARDSON P J, MACDOUGALL A S, STANLEY A G, et al. Inversion of plant dominance-diversity relationships along a latitudinal stress gradient[J]. Ecology, 2012, 93(6): 1431–1438.
[36] BERTUZZO E, CARRARA F, MARI L, et al. Geomorphic controls on elevational gradients of species richness[J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(7): 1737–1742.
[37] MERCKX V S F T, HENDRIKS K P, BEENTJES K K, et al. Evolution of endemismon a young tropical mountain[J]. Nature, 2015, 524(7565): 347–362.
[38] 于顺利, 陈宏伟, 李晖. 种子重量的生态学研究进展[J]. 植物生态学报, 2007, 31(6): 989–997.
[39] 高乐旋. 入侵种喜旱莲子草表型可塑性变异和快速适应的分子基础[D]. 上海: 复旦大学, 2012: 1–203.
[40] 朱志红, 王刚. 燕麦表型可塑性与繁殖分配的研究[J]. 兰州大学学报, 2002, 38(1): 76–83.
[41] 刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学:生命科学, 2015, 45(4): 325–339.
[42] POTTER T S, BOWMAN W D. Testing invasion filters for the alpine: the roles of temperature, nitrogen deposition and soil[J]. Biological Invasions, 2020, 22(6): 1889–1901.
[43] THEOHARIDES K A, DUKES J S. Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion[J]. New Phytologist, 2007, 176(2): 256–273.
[44] 李明阳, 巨云为, KUMAR S, 等. 美国大陆外来入侵物种斑马纹贻贝()潜在生境预测模型[J]. 生态学报, 2008, 28(9): 4253–4258.
[45] 梁玉. 外来种大花金鸡菊入侵的影响因子及其遗传多样性研究[D]. 济南: 山东大学, 2007: 1–147.
[46] 赵见明. 瑞丽主要外来入侵植物[J]. 西南林学院学报, 2007, 27(1): 20–24.
[47] 申时才, WILLSON A, MELICK D. 滇西北高山牧场入侵物种土大黄生态学调查[J]. 西南林学院学报, 2006, 26(3): 11–14.
Effects of elevational gradients on reproductivity and seed dispersal ability ofin mountain ranges
CHEN Xiaoyan, ZHANG Wengang, LIU Ruiling, LIU Gang*
College of Life Sciences, Shaanxi Normal University, Xi'an 710119, China
Due to changes on natural selection pressure, the reproductive capacity and seed dispersal ability of invasive plants would change along elevational gradients during their expansion in mountain ranges. To test the changing pattern of these traits of invasive plant, we conducted a field survey on the elevational populations of(505-1947m) in Qinling-Bashan Mountains. We compared the differences on population coverage, density, seed production per plant, and the mass-area ratio (MAR) of seeds among these elevational populations. In addition, the growth and reproduction traits ofin different habitats (farmland and uncultivated land) were also compared. The results showed that: (1) the population coverage and density ofdid not change significantly with increasing altitude. (2) The number of seeds per inflorescence increased while the number of inflorescences per plant and the seed production (quantity and mass) per plant decreased with increasing altitude. (3) The MAR ofseed increased significantly with increasing altitude, which suggested a decreased seed dispersal ability ofalong elevation. (4) Compared to the populations in uncultivated land, populations ofin farmland had higher coverage,higher seed biomass per plant and smaller mass-area ratio. All in all, our results suggested that the reproductive capacity and the seed dispersal potential ofdecreased significantly with increasing altitude in Qinling-Bashan Mountains. Moreover,in farmland had higher reproductive capacity and the seed dispersal ability than those in uncultivated land. It suggests that the Qinling–Bashan Mountains play an important role in reducing the dispersal rate of, however, agricultural activities can promote its invasion and expansion process.
invasive plants;; Qinling-Bashan Mountain; seed diffusivity; elevational gradients
陈晓艳, 张文刚, 刘芮伶, 等. 海拔对入侵植物粗毛牛膝菊种群繁殖力与种子扩散力的影响[J]. 生态科学, 2022, 41(3):44–53.
CHEN Xiaoyan, ZHANG Wengang, LIU Ruiling, et al. Effects of elevational gradients on reproductivity and seed dispersal ability ofin mountain ranges[J]. Ecological Science, 2022, 41(3): 44–53.
10.14108/j.cnki.1008-8873.2022.03.005
Q948.1
A
1008-8873(2022)03-044-10
2020-08-12;
2020-08-27
国家自然科学基金(31600445); 陕西省自然科学基础研究计划(2020JM-286); 中央高校项目(GK201703036, 2019CSLY019和2020CSLY014)
陈晓艳(1997—), 女, 硕士, 主要从事全球气候变暖与入侵生态学研究, E-mail: 1537044725@qq.com
刘刚, 男, 博士, 副教授, 主要从事全球气候变暖与入侵生态学研究, E-mail: gangliu@snnu.edu.cn
