S-腺苷甲硫氨酸脱羧酶基因对银腺杨84K抗旱性的影响*
2022-05-11姚俊广刘依静黄李超卢孟柱
姚俊广 耿 娅 刘依静 安 轶 黄李超 曾 为 卢孟柱
(浙江农林大学林业与生物技术学院 省部共建亚热带森林培育国家重点实验室 杭州 311300)
我国森林覆盖率低,木材蓄积量低。作为木制品消费大国,木材供应严重不足。随着经济的发展,木材需求量日益增多。20世纪末至21世纪初,我国营造了超过1 000万hm2的速生林基地,成为世界上人工林面积最大的国家(主楚杰等, 2015)。杨树(Populus)作为重要的速生人工林树种,已成为解决木材供需矛盾的重要途径。我国大约有50%以上的地区属于干旱或半干旱,其中半干旱地区的人工造林成活率约为 30%,而干旱区仅为4%(黄荣辉等, 2012)。干旱已经成为严重阻碍各地区林业发展以及改善生态环境的重要因素之一,了解树种对干旱胁迫响应以及如何提高其抗逆性具有至关重要的意义。杨树人工林在干旱、半干旱地区的发展,依赖于抗逆品种。
胁迫条件下,植物能够做出相应的防御机制,包括形态调节以及生理生化反应(Pandeyetal., 2015)。Todorova等(2015)研究发现内源多胺含量升高是植物响应胁迫反应之一,对植物的抗逆性具有至关重要的作用。多胺作为一类带正电荷的脂肪族类化合物,普遍存在于动植物体内(Taboretal., 1984),可参与调节细胞分裂(Evansetal., 1989)、果实成熟(Guoetal., 2018)等植物发育过程,也可响应低温、干旱等胁迫途径(Liuetal., 2015)。高等植物的多胺主要分为腐胺(Put)、亚精胺(Spd)、精胺(Spm)、以及热精胺(T-spm)4类(Takanoetal., 2012)。腐胺和亚精胺是最常见的多胺,精胺主要存在于种子植物、多数动物和一些细菌中(Peggetal., 2010),而热精胺存在于整个植物界中(Eugenioetal., 2008)。多胺的合成途径受多种相关酶的催化,其合成过程主要有2种: 以鸟氨酸(ornithine, Orn)为底物,在鸟氨酸脱羧酶(ornithine decarboxylase, ODC)催化下,将鸟氨酸直接转化为腐胺的直接途径; 以精氨酸(arginine,Arg)为底物,在精氨酸脱羧酶(arginine decarboxylase, ADC)催化以及经N-氨甲酰腐胺脱去一分子氨,生成腐胺的间接途径。而腐胺能够作为亚精胺、精胺生物合成途径的主要前体。在亚精胺合成酶(spermidine synthases, SPDS)的催化下生成亚精胺,亚精胺在精胺合成酶(spermine synthases, SPMS)以及热精胺合成酶(thermospermine synthases, TSPMS)催化下分别生成精胺和热精胺(Walters, 2003; Tiburcioetal., 2014)。在合成亚精胺和精胺途径上,S-腺苷甲硫氨酸(S-adenosylmethionine, SAM)在S-腺苷甲硫氨酸脱羧酶(S-adenosylmethionine decarboxylase,SAMDC)催化下,生成脱羧后的S-腺苷甲硫氨酸(decarboxylated S-adenosylmethionine, dcSAM),提供亚精胺和精胺合成所需要的氨丙基,成为多胺合成途径上重要的限速步骤(Huetal., 2005)。
施加外源多胺,可以保护植物免受胁迫损伤。干旱胁迫下,对植株叶片喷施腐胺能够有效改善植株水分状况,提高植物的生物量,增强干旱胁迫耐受性 (Guptaetal., 2012; Hussainetal., 2013; Zhuetal., 2019)。内源提高多胺相关酶基因的表达水平,可进一步证明多胺对于非生物胁迫的适应性(Pathaketal., 2014; Shietal., 2014; Liuetal., 2015)。在盐胁迫下,过表达SAMDC可使水稻(Oryzasativa)生物量增加,内源Spd、Spm的积累提高了抗胁迫能力(Royetal., 2002)。甜菜(Betavulgaris)M14-SAMDC基因异源转化拟南芥(Arabidopsisthaliana),拟南芥Spd和Spm含量增加,植株表现出强耐盐性(Jietal., 2019); 干旱胁迫下,辣椒(Capsicumannuum)CaSAMDC异源转化拟南芥,其过表达CaSAMDC植株亚精胺、精胺含量显著升高,植株耐旱性显著增强(Sooetal., 2014)。因此,通过改变植物中精胺含量,可以调控植物的抗逆性。
杨树作为多年生木本植物,其生长发育长期受到各种胁迫的制约,研究其抗逆机制具有重要意义。本研究以PagSAMDC4a转基因银腺杨84K(Populusalba×P.glandulosa‘84K’)作为试验材料,研究其在干旱条件下生长发育的变化,进一步分析胁迫条件下植株内源多胺水平差异,系统地揭示杨树内源性多胺在干旱胁迫反应中的重要作用。
1 材料与方法
1.1 试验材料
银腺杨84K为笔者试验室保存,干旱胁迫处理所用过表达PagSAMDC4a株系为试验室前期研究所获得,经定量PCR检测后选取过表达PagSAMDC4a转基因银腺杨3个株系(PagSAMDC4a-OE#3、PagSAMDC4a-OE#15、PagSAMDC4a-OE#17)进行研究。组培苗生长1个月后进行土培,在装有混合基质(泥炭︰蛭石︰珍珠岩体积比为3︰9︰1)的育苗盆培养3个月,培养条件为: 温度23~25 ℃,相对湿度60%,光周期16 h光照/ 8 h黑暗,光照强度50 μmol · m-2s-1。
1.2 干旱胁迫处理
以生长3个月的未转基因银腺杨84K及其过表达PagSAMDC4a转基因株系为材料,设置对照组和干旱胁迫组。对照组保持充分水量,确保育苗盆中混合基质的水分含量保持在约70%; 干旱胁迫组模拟自然干旱的模式,处理时间为6天,土壤含水量降至约25%,未转基因84K杨叶片出现严重枯萎现象。观测植株的生长状况并取材进行相关指标的测定。
1.3 干旱胁迫下相关指标的测定
在干旱处理第6天,分别取未转基因银腺杨84K和过表达PagSAMDC4a植株叶片,测定相关指标。
1.3.1 内源多胺含量 取未转基因植株、过表达PagSAMDC4a植株由上至下第4和第5片叶片测定多胺含量(林绍艳等, 2016)。具体步骤: 配制腐胺、亚精胺、精胺标准品; 样品加入5%高氯酸浸提以及后续样品、标准品的衍生化处理; 使用超高效液相色谱仪(UPLC)进行多胺含量测定,选用色谱柱Poroshell 120 EC-C18(150 mm×3.0 mm×2.7 μm),设置参数: 流动相乙腈和水(体积比40︰60),流速为0.5 mL·min-1,检测波长为230 nm,进样体积为5 μL; 同时测定标准品腐胺(Put)、亚精胺(Spd)、精胺(Spm)建立标准曲线(表1)。

表1 多胺测定的回归方程和相关系数①
1.3.2 过氧化氢(H2O2)含量 取植株由上至下第1~3片叶片,参考H2O2检测方法(Leietal., 2013),选用H2O2检测试剂盒(A064-1-1)、蛋白定量(TP)测定试剂盒(A045-2)(南京建成生物工程研究所),测定样品蛋白浓度(考马斯亮蓝测定法)以及H2O2与钼酸作用形成的络合物在波长405 nm处吸光值,计算H2O2含量: H2O2含量(mmol·g-1prot)=(测定OD值-空白OD值)/(标准OD值-空白OD值)×标准品浓度(163 mmol·L-1)÷待测样品蛋白浓度(g prot·L-1)。
1.3.3 叶片相对含水量 取正常情况以及干旱胁迫处理的未转基因银腺杨84K、过表达PagSAMDC4a植株的由上至下第7片叶片测定含水量。首先取下新鲜的叶片后立即称量叶片的鲜质量(FW),而后把叶片浸没于水中室温放置24 h后称量叶片浸水后的质量(TW)。再将新鲜叶片放置在80 ℃恒温干燥箱里干燥72 h后,称量叶片的干质量(DW)。计算叶片相对含水量(Heetal., 2018): (FW-DW)/(TW-DW)×100%。
1.3.4 叶片失水率 取正常情况下未转基因银腺杨84K、过表达PagSAMDC4a植株由上至下第8片叶片,分别称取叶片的鲜质量(FW),每隔0.5 h称量1次(desiccated weight),总时间为4.5 h。最后将叶片放置80 ℃烘箱里烘干过夜,称其干质量(DW)。计算叶片失水率(Boetal., 2017): 叶片失水率(%)=(FW-desiccated weight)/(FW-DW)×100%。
1.3.5 电解质渗透率 取正常情况以及干旱胁迫下的未转基因银腺杨84K、过表达PagSAMDC4a植株由上至下第6片叶片测定电解质渗透率。首先用去离子水清洗叶片,经滤纸擦干叶片表面后,使用打孔器打出10片小圆片放入装有2 mL ddH2O的离心管中,完全浸泡并真空抽滤10 min后,向离心管中加入6 mL ddH2O,室温放置200 r·min-1摇床中振荡1 h,测其电导率为S1; 将离心管放置90 ℃水浴20 min,待溶液冷却至室温后充分摇匀,测定其电导率为S2。计算电解质渗透率(%)=(S1/S2)×100%。
1.4 数据分析
所有试验均进行10次生物学重复,样本数据采用SPSS 17.0数据处理软件进行独立样本t检验,在0.01水平上检验差异显著性,并用Excel或GraphPad prism8进行数据分析、图表绘制。
2 结果与分析
2.1 PagSAMDC4a转基因银腺杨84K内源多胺含量
经定量PCR分析,银腺杨84K转基因株系PagSAMDC4a-OE#3、PagSAMDC4a-OE#15、PagSAMDC4a-OE#17的基因表达量均显著高于未转基因植株,且基因表达量依次由低到高(图1A)。测定转基因株系和未转基因植株的内源多胺含量(图1B),采用UPLC测定多胺标准品和样品的色谱图(图1C、D),根据不同的标准品浓度、色谱峰面积计算标准曲线(表1)。在水分正常情况下,3个转基因株系的内源亚精胺、精胺含量都显著高于未转基因对照,其中PagSAMDC4a-OE#17株系的内源腐胺、亚精胺、精胺含量分别是未转基因对照的1.95、3.43、1.32倍(表2)。
2.2 干旱胁迫下PagSAMDC4a转基因银腺杨84K叶片相关生理指标
干旱胁迫影响植株的生长发育,且在叶片表型方面尤其明显。正常水分条件下,与未转基因对照相比,过表达PagSAMDC4a植株的叶片表型(图2A)、叶片相对含水量(图2B)、电解质渗透率(图2C)并无显著差异,但叶片失水较慢,失水率由高到低为未转基因对照、PagSAMDC4a-OE#3、PagSAMDC4a-OE#15、PagSAMDC4a-OE#17(图2 D)。在干旱胁迫下,未转基因植株表型出现明显变化,叶片严重萎蔫下垂,表现对干旱胁迫的敏感性; 而过表达PagSAMDC4a植株叶片出现不同程度的响应,PagSAMDC4a-OE#3植株叶片轻度萎蔫,PagSAMDC4a-OE#15、PagSAMDC4a-OE#17植株叶片生长状况正常,表现出了耐旱性(图2A)。同时测定叶片相对含水量、电解质渗透率等相关指标后发现,在干旱胁迫下,未转基因对照的叶片相对含水量比正常供水条件下降低26.44%(图2B),电解质渗透率升高27.68%(图2C); 而PagSAMDC4a-OE#3、PagSAMDC4a-OE#15、PagSAMDC4a-OE#17植株叶片相对含水量比正常供水条件下分别降低10.9%、3.66%、1.33%(图2B),电解质渗透率与正常供水条件下无显著差异(图2C)。过表达PagSAMDC4a植株电解质渗透率较低、变化幅度较小,叶片的相对含水量较高,并与未转基因对照差异显著,表明其抗旱性明显加强。
2.3 干旱胁迫下PagSAMDC4a转基因银腺杨84K叶片内源H2O2含量
为验证干旱胁迫下84K杨植株氧化损伤情况,测定正常供水及干旱胁迫下植株叶片的H2O2含量(图3)。在正常供水条件下,相比未转基因对照,转基因植株的H2O2含量显著降低,H2O2含量由高到低依次为未转基因对照 >PagSAMDC4a-OE#3 >PagSAMDC4a-OE#15 >PagSAMDC4a-OE#17。干旱胁迫6天后,未转基因对照的H2O2含量显著升高,而过表达PagSAMDC4a植株H2O2含量增加幅度显著低于对照,含量由高到低为未转基因对照 >PagSAMDC4a-OE#3 >PagSAMDC4a-OE#15 >PagSAMDC4a-OE#17。以上结果表明过表达PagSAMDC4a明显增强了转基因84K杨的抗旱性。

图1 PagSAMDC4a转基因84K杨的基因表达及多胺含量

表2 PagSAMDC4a转基因84K杨叶片中3种多胺含量①
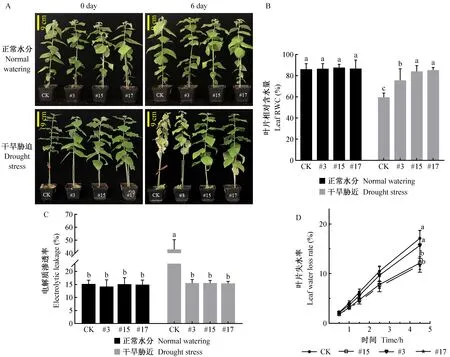
图2 干旱胁迫下过表达PagSAMDC4a植株相关生理指标的变化
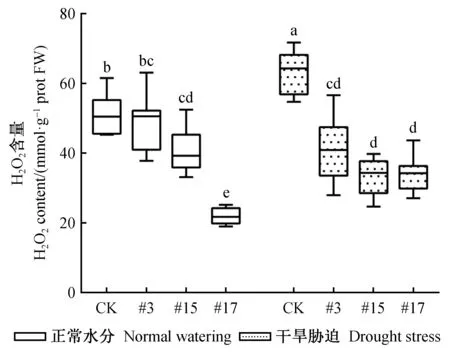
图3 干旱胁迫下PagSAMDC4a转基因84K杨的H2O2含量
3 讨论
干旱严重影响植物生长发育以及生物量积累(Yinetal., 2014),阻碍干旱、半干旱地区的植物生长。植物内源性多胺影响植物的生长发育以及胁迫响应,在干旱、低温、高温以及机械损伤等多种胁迫下,植物能产生大量的多胺物质(Alczaretal., 2006; Marcoetal., 2011)。植物外源施加多胺或者异源表达多胺合成酶,都能增强植物抵御胁迫的能力。多胺合成涉及多个酶促反应,相关酶基因的作用及其对多年生木本植物的影响有待研究。
提供氨丙基的S-腺苷甲硫氨酸脱羧酶(SAMDC)在多胺合成途径上发挥着关键作用。因此,本研究通过对SAMDC4a过表达银腺杨84K植株进行干旱胁迫,分析了SAMDC4a基因差异表达导致植物体内源多胺含量变化对植株抗逆性的影响。干旱胁迫后,过表达SAMDC4a植株叶片呈现不同的响应状态,随着基因表达量的升高,其内源亚精胺和精胺含量都随之增加。这进一步说明过表达SAMDC4a植株基因表达差异影响内源多胺含量,SAMDC4a基因表达升高导致植株多胺含量增加,进而增强对干旱胁迫的耐受性。此外,为完全评估SAMDC4a基因在抗逆性中的作用,后期应对SAMDC4a进行基因沉默或RNA干扰,进一步研究SAMDC4a基因表达量降低对多胺含量及抗逆性的影响。
植物在受到胁迫时,电解质渗透率是分析植物发生损伤的重要标准,同时电解质渗透率的测定结果可用来评估逆境胁迫后的细胞活力(Busaidietal., 2015)。据报道在低温胁迫下,对黄瓜(Cucumissativus)幼苗施加外源腐胺和亚精胺,能有效降低黄瓜叶片的电解质渗透率,对低温产生相应的抗逆性(Zhangetal., 2009)。本试验在干旱胁迫后,未转基因银腺杨84K的电解质渗透率显著升高,而过表达SAMDC4a植株的渗透率变化幅度较小,且维持在较低水平,这表明通过改变植株内源多胺含量,可以对植株电解质的渗透有重要影响,稳定了细胞透性,减少了对细胞的伤害。叶片相对含水量是衡量植株响应干旱胁迫的重要生理指标,可用来评估植物抗旱能力。在正常供水情况下,过表达SAMDC4a转基因银腺杨84K植株与未转基因植株的叶片相对含水量无显著差异; 但在干旱胁迫下,转基因植株叶片相对含水量显著高于未转基因对照。与之相反,过表达SAMDC4a转基因植株在正常供水情况下的叶片失水率显著低于未转基因对照。这些结果表明,多胺通过增强叶片持水能力,减少叶片水分丢失,进而有效缓解植株叶片所受水分胁迫,从而改善叶肉细胞中水分状况,在有限的水分供应下保证植株正常生长(Farooqetal., 2009)。
本研究表明,多胺合成关键酶基因SAMDC4a过量表达加速了多胺(尤其是亚精胺、精胺)的合成,促进了多胺的积累,并间接引起多胺相关的生理、生化变化。H2O2作为多胺代谢相关产物之一(Groppaetal., 2008; Alczaretal., 2010),在植物应对逆境胁迫中具有重要影响。在多种胁迫条件下,多胺能够缓解活性氧(ROS)带来的损伤、激活抗氧化机制,对各种胁迫产生抗逆性(Seoetal., 2019)。H2O2作为活性氧主要类型之一,在植株干旱胁迫下的积累是造成植株自身损伤的一个重要途径。对植株施加外源多胺时,会导致植株H2O2含量下降、抗氧化机制的激活、ROS水平显著降低等,可以缓解干旱胁迫的影响(Satishetal., 2018; Hassanetal., 2018; 2020)。本研究表明,过表达SAMDC4a银腺杨84K,在干旱胁迫下的H2O2含量变化幅度较小,且显著低于未转基因对照植株,进一步表明多胺可以减少活性氧积累,减轻干旱胁迫引起的氧化损伤程度。说明过表达SAMDC4a基因能增强杨树抗旱能力,增强对环境胁迫的抗逆性。
4 结论
S-腺苷甲硫氨酸脱羧酶基因(SAMDC4a)过表达的银腺杨84K可通过提高植株内源多胺含量,从而增强叶片的持水能力,减轻ROS积累,维持正常的细胞渗透能力,在干旱胁迫下增强对干旱胁迫的抗性,说明内源多胺在木本植物响应干旱胁迫过程中有重要作用。这为揭示杨树多胺在抗逆性方面的作用提供了重要依据,对选育抗逆新品种具有潜在的应用价值。
