亚热带不同林龄杉木人工林红壤线虫群落结构特征*
2022-05-11郑荧枫王建青邹秉章王思荣施秀珍余再鹏黄志群
郑荧枫 王建青 邹秉章 王思荣 施秀珍 余再鹏 黄志群
(1.福建师范大学地理科学学院 福州 350007; 2.福建师范大学湿润亚热带生态-地理过程教育部重点实验室 福州 350007; 3.福建省上杭白砂国有林场 上杭 364205)
线虫是土壤中数量最丰富的微型后生动物之一,广泛分布于各种土壤中(Hoogenetal., 2019)。土壤线虫种类丰富,占据了土壤食物网的不同营养级,在揭示土壤环境变化等方面具有重要作用 (Yeatesetal., 1993; Wardleetal., 1995; Bongersetal., 1998; Wangetal., 2019)。研究表明,土壤线虫通过促进植物与土壤间的负反馈、改变根际微生物群落和促进养分循环而影响植物生长(Wilschutetal., 2021)。同时,植物特性对维持土壤线虫多样性有重要调节作用(Zhangetal., 2015)。凋落物、植物根系和根系分泌物为地下动物群落生存发展提供了基础资源(Fuetal., 2009)。植物还会通过改变土壤特性进而影响土壤线虫群落。土壤线虫群落受土壤养分含量(Bjrnlundetal., 2012)、pH值(Kitagamietal., 2020)、含水量(Bakonyietal., 2007)等土壤因子调控影响。因此,研究土壤线虫与环境因子的关系有助于进一步了解土壤线虫群落特征及其与植被之间的关系(Zhietal., 2009; 陈利, 2011)。森林生长会改变土壤特征和线虫食物资源,土壤线虫群落结构也必然随林龄变化。迄今为止,关于不同林龄土壤线虫群落组成的研究已有较多报道(Zhangetal., 2015; Guanetal., 2015; 佟富春等, 2009),如Panesar等(2001)在对温哥华岛不同林龄冷杉(Pseudotsugamenziesii)的研究发现线虫数量虽在幼林阶段较低但随时间推移逐渐增加到成过熟林水平。Zhang等(2015)研究发现,温带森林中线虫数量及多样性在中龄林阶段最高,而在老龄林阶段因植物多样性下降及线虫群落不同类群间的竞争增加而下降。林龄对线虫群落的影响可能会由于经营方式、种植年限和植被类型不同发生变化(Kardoletal., 2005; Huetal., 2016; Scheuetal., 2003)。
杉木(Cunninghamialanceolata)是我国主要的速生造林树种,在我国森林资源中占有重要地位(梁萌杰等, 2016)。杉木人工林面积现已达全国人工林优势树种首位,约占总人工林面积的30%(国家统计局, 2020)。杉木人工林面积增加和种植年限延长对土壤性质和土壤生物产生了不同程度的影响。在阔叶林皆伐地上营造杉木人工林会造成许多生态环境问题,如土壤养分流失、肥力下降、植被多样性降低等,从而影响土壤生物群落(张芸等, 2019; 梁萌杰等, 2016; 周德明等, 2012)。以往研究表明,随着杉木人工林生长,先是林分郁闭度和林下植被多样性增加,植物凋落物累积(张芸等, 2019; Suetal., 2020; 梁萌杰等, 2016); 但当杉木林发育到一定阶段时,由于土壤养分限制作用加强、土壤pH值降低等,使土壤质量下降(周德明等, 2012; 王振宇等, 2019)。因此,本研究假设如下: 在杉木林生长过程中,线虫密度、多样性均有所增加,瓦斯乐斯卡指数下降; 线虫群落结构也会随林龄增加发生明显变化。为验证以上假设,本研究在亚热带地区选取了5个生长阶段(5、8、21、27、40年)的杉木人工林,研究土壤线虫群落组成和结构特征以及其与土壤理化性质的关系受林龄的影响,为我国亚热带地区杉木人工林可持续经营提供科学依据和土壤动物多样性的基础数据。
1 研究区概况
研究区位于福建省龙岩市上杭县白砂国有林场(24°46′—25°27′N, 116°15′—116°56′E),海拔400~800 m,属中亚热带季风气候。年均降水量约1 600 mm,年均气温约20.1 ℃,年日照时数约1 800 h,无霜期277天左右。主要地貌类型为低山丘陵,坡度10°~40°,土壤是由花岗岩发育而来的红壤。林下植被主要有闽楠(Phoebebournei)、细枝柃(Euryaloquaiana)、毛冬青(Ilexpubescens)、梅叶冬青(I.asprella)、芒萁(Dicranopterisdichotoma)、江南卷柏(Selaginellamoellendorffii)、狗脊蕨(Woodwardiajaponica)、黄樟(Cinnamomumporrectum)、中华里白(D.chinensis)等(Suetal., 2020)。
2 研究方法
2.1 样地设置
利用空间替代时间的方法,选取了幼龄林(5年)、中幼龄林(8年)、近熟林(21年)、成熟林(27年)和过熟林(40年)5个发育阶段的杉木人工林,其中8年和21年设置3个重复,5、27和40年分别设置4个重复,共18块样地,每块样地间距大于500 m,样地面积为20 m×30 m。
2.2 研究方法
2.2.1 土壤线虫的采集与鉴定 2019年7月进行土壤线虫采集。采用土钻法,在每个样地内,用内径为3.5 cm的土钻沿“S”型在12个点,钻取0~10 cm土层的土壤,将各样地的土壤均匀混合成1个土壤样品,装入已编号的自封袋中运回实验室,一部分用于线虫分离提取,一部分过筛、风干后测定土壤理化性质。采用浅盘法从每个样地的100 g新鲜土壤中收集线虫个体。在显微镜下计数线虫总数,以估计每个样地的线虫种群数量(线虫种群大小)。随后,每个样本随机抽取150条线虫,主要依据尹文英(1998)的《中国土壤动物检索图鉴》和谢辉(2005)的《植物线虫分类学》,在光学显微镜下鉴定线虫科属(不足150条的全部鉴定)。将所有采集到的线虫,基于网页http:∥Nemaplex.ucdavis.edu提供的依据,按照取食习性和食道特征,分为4个主要的营养类群(Yeates, 2003): 食细菌线虫(Bacterivores)、食真菌线虫(Fungivores)、植物寄生线虫(Plant parasites)和捕食-杂食性线虫(Omnivores-predators); 根据线虫的生活史策略,分为从r-策略者向k-策略者过渡的5个c-p(colonizer-persister)类群,并赋予其c-p值(Bongersetal., 1998)。
2.2.2 土壤理化性质测定 土壤质量含水率用烘干法测定(105 ℃); 土壤pH值用电位法测定 (土水比为1∶2.5); 土壤总碳、总氮含量用元素分析仪(Elemental EL MAX CNS analyzer,德国)测定; 总磷含量用HClO4-H2SO4消煮法测定; 矿质氮素含量用总有机碳分析仪(TOC,日本岛津)测定。有效磷含量在NaHCO3提取后用连续流动分析仪测定(Skalar San++,Skalar,荷兰)。土壤基本特征见表1。

表1 不同林龄杉木人工林0~10 cm土壤基本特征①
2.2.3 线虫生态指数计算 土壤线虫数量用每100 g干土中线虫的个数表示。优势属为占总线虫数10%以上的属,常见属为占总线虫数1%~10%以上的属,稀有属为占总线虫数1%以下的属。土壤线虫生态指数计算公式如下: 1) 香农多样性指数(H′):H′=-ΣPi×lnPi,式中Pi是某个属(i=1,2,3,…)的相对丰度; 2) 植物寄生线虫成熟度指数(plant-parasites index,PPI): PPI=Σ(Ci×Pi),式中Ci为植物寄生线虫第i类群c-p值; 3) 瓦斯乐斯卡指数(wasilewska,WI): WI=(FF+BF)/PP,式中FF(relative abundance of fungivores)为食真菌线虫相对丰度,BF(relative abundance of bacterivores)为食细菌线虫相对丰度,PP(relative abundance of plant-parasites)为植物寄生线虫相对丰度。
2.2.4 数据处理 采用SPSS 21.0软件对所有数据进行单因素方差分析(One-way ANOVA),并进行显著性检验(LSD),以评价不同林龄之间线虫群落的差异,P< 0.05表示具有统计学意义上的显著差异。进行Pearson相关分析,检验土壤理化性质与线虫数量、各营养类群线虫数量和相对丰度、线虫生态指数的相关性。利用非度量多维度进行线虫群落排序分析,利用R 3.6.3和Excel制图。
3 结果与分析
3.1 土壤线虫的密度与群落组成
由表 2可知,从杉木人工林土壤中,共采集到线虫3 397条,经鉴定隶属于67个属,其中根结属(Meloidogyne)是除5年生杉木林以外所有林分的优势属,占总个体数的15.9%。常见属有丝尾属(Oxydirus)、螫属(Pungentus)、丝尾垫刃属(Filenchus)和茎属(Ditylenchus)等21属,合计占总个体数的68.3%。稀有属包括柱咽属(Cylindrolaimus)、三孔属(Tripyla)、原杆属(Protorhabditis)和巴氏属(Bastiania)等45属,合计占总个体数的15.8%。杉木林不同生长阶段的优势属有所差异,5年的优势属为丝尾垫刃属,8年的优势属为丝尾属、根结属和螫属,21年的优势属仅有根结属,27年和40年的优势属均为丝尾属和根结属。不同生长阶段杉木人工林土壤线虫总密度变化范围为464~659条·(100 g)-1干土,土壤线虫总密度在不同林龄间无显著差异(P> 0.05)(图1)。
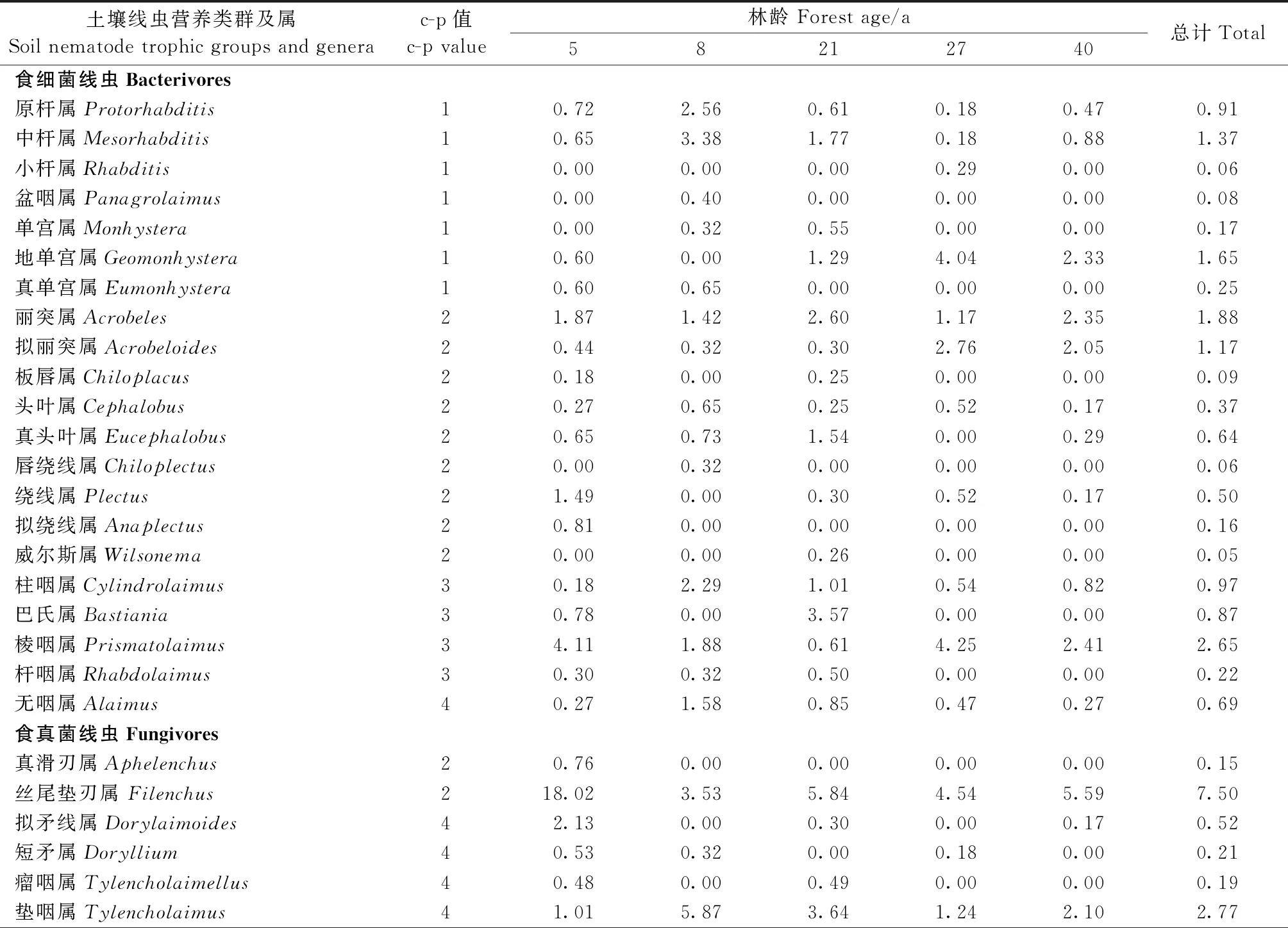
表2 不同林龄杉木人工林土壤线虫属的平均丰度
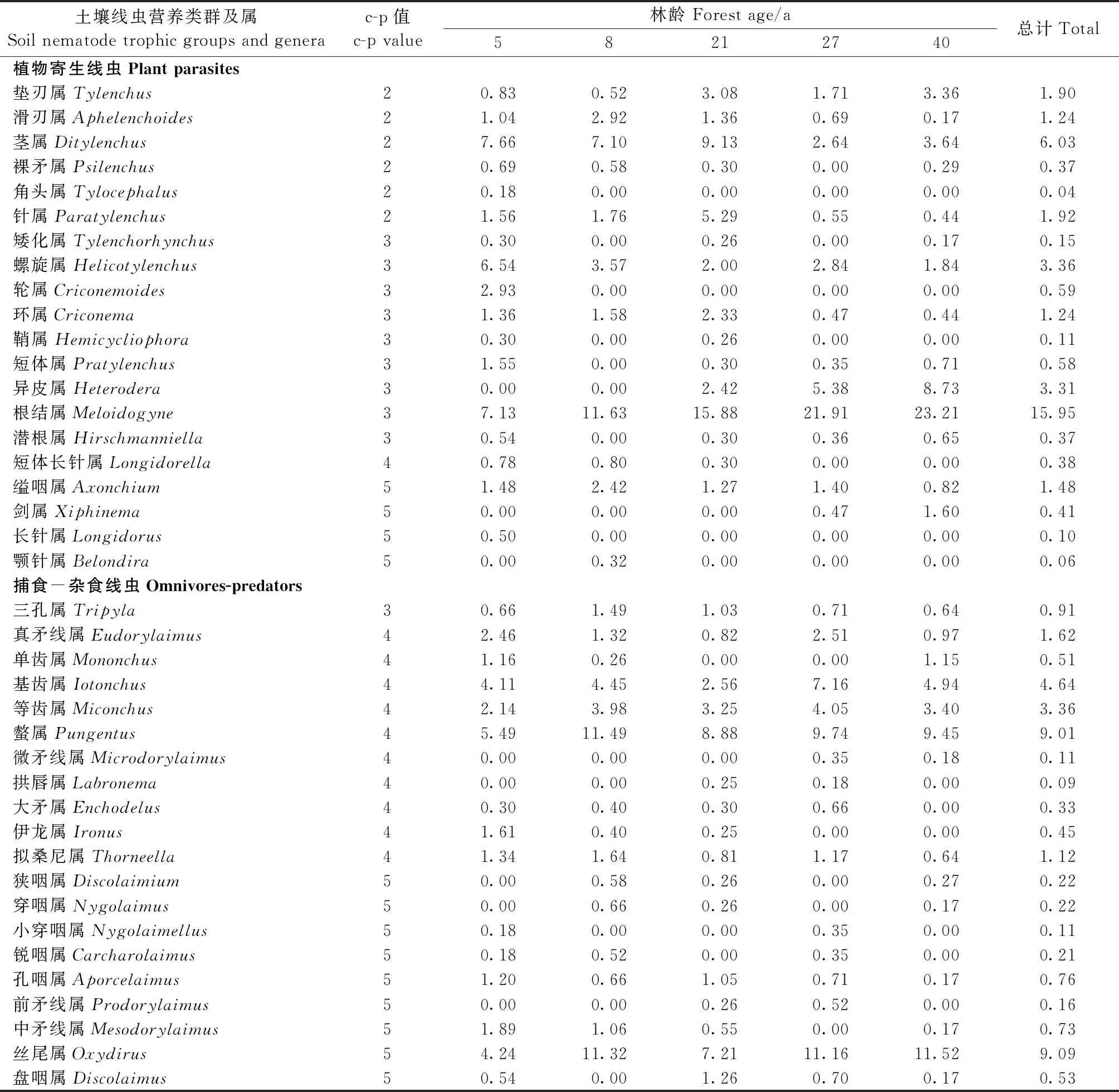
续表 Continued

图1 不同林龄杉木人工林土壤线虫密度
3.2 土壤线虫营养类群的林龄差异
4个营养类群中,植物寄生线虫的相对丰度最高,变化范围为23.98%~47.04%; 捕食-杂食线虫次之,为23.05%~30.93%; 食细菌线虫和食真菌线虫的相对丰度分别为12.21%~16.82%、16.02%~28.26%(图2)。由此可知,植物寄生线虫是本研究林分样地的主要功能类群。不同生长阶段杉木林土壤食细菌线虫、食真菌线虫和捕食-杂食类线虫的相对丰度无显著差异(P>0.05)。杉木林生长改变了植物寄生线虫相对丰度,在8年最低,之后逐渐增加,40年达到最高值(表3)。林龄对食细菌线虫、植物寄生线虫和捕食-杂食类线虫密度没有显著影响(P>0.05)。杉木林生长改变了食真菌线虫的密度,林龄为5年的林分显著高于其他林龄(P<0.05)。

图2 不同林龄杉木人工林土壤线虫各营养类群比例

表3 不同林龄杉木人工林线虫营养类群密度
3.3 土壤线虫群落结构特征
非度量多维尺度分析(NMDS)的应力系数(Stress)值小于 0.2(图3),表明NMDS分析可以反映不同林龄间线虫群落结构的差异程度。NMDS分析结果表明,林龄为40年土壤线虫群落结构与5年、8年存在明显差异。8年与27年土壤线虫群落结构差异显著。以上结果表明林龄可以改变土壤线虫群落结构。
3.4 土壤线虫生态指数的林龄差异
如表3所示,土壤线虫的香农多样性指数(H′)随林分生长未发生显著变化(P>0.05)。杉木林土壤的植物寄生线虫成熟度指数(PPI)在杉木林发育过程的变化范围为2.62~2.98。林龄为27年和40年的PPI显著高于21年(P<0.05)。瓦斯乐斯卡指数(WI)在杉木林生长过程的变化范围为0.44~1.09,林龄5年的WI值最高,显著高于40年(P<0.05),表现出WI随着林龄增加逐渐下降的趋势(表4)。
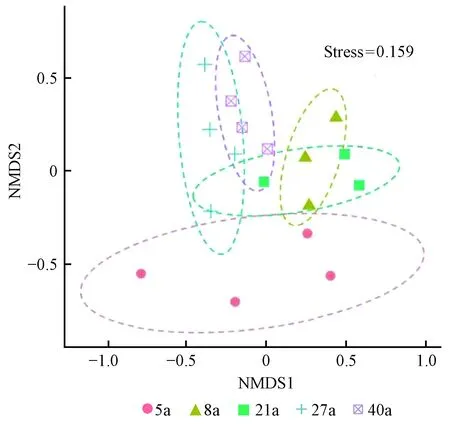
图3 土壤线虫群落非度量多维度排序

表4 不同林龄杉木人工林线虫生态指数
3.5 土壤理化性质与线虫群落特征的相关性
由表5可知,土壤pH值与WI呈显著正相关。TC与植物寄生线虫密度及相对丰度呈显著正相关。矿质氮与捕食-杂食线虫密度及相对丰度呈显著负相关。速效磷与食真菌线虫相对丰度呈显著正相关。土壤碳磷比与植物寄生线虫密度及相对丰度呈显著正相关,与捕食-杂食线虫相对丰度呈显著负相关。

表5 杉木土壤理化性质和线虫指标的相关系数①
4 讨论
大量研究表明,林龄显著影响土壤线虫密度及多样性(Zhangetal., 2015; Guanetal., 2015; Huetal., 2016)。但在本研究中,土壤线虫密度及多样性随林龄增加无显著变化,这与假设不一致。李树战等(2016)研究也发现,不同林龄杉木林的土壤线虫多样性没明显变化。有研究发现林分生长改善了土壤环境(含水量、孔隙度等),增加了线虫食物的可利用性(植物根系、根系分泌物等),直接或间接促进了土壤线虫密度增加(Huetal., 2016)。本研究中,由于林龄对土壤含水量、土壤碳、土壤氮和pH值的影响均未达到显著水平,因此土壤性质对线虫密度及多样性的影响较小,表现为线虫密度随林龄增加没有显著变化。Zhi等(2009)在研究固定沙丘线虫演替过程中也发现,线虫密度在森林演替过程中没有明显变化,这可能是由于资源限制(自下而上的限制)在明显调控线虫密度(Thorntonetal., 2002)。Zhang等(2015)对不同林龄温带森林土壤线虫的研究发现,土壤线虫总香农多样性指数在幼龄林最低,中龄林最高,产生这种变化的原因与造林树种多样性有关。而在本研究中,杉木人工林树种单一,林下植被多样性也没有显著变化,导致土壤线虫总香农多样性指数变化不大(Suetal., 2020)。根据非度量多维度分析,本研究40年林分土壤线虫群落结构与5年、8年林分差异明显。21年、27年与40年林分在土壤线虫群落结构上存在一定的相似性,说明杉木林在生长过程中线虫群落结构特点出现了变化,这与假设一致。这种变化反映了杉木人工林生长过程中地下资源输入和土壤环境的变化(Yeates, 2007; Guanetal., 2015)。
杉木林生长对各类群线虫的影响不同。林龄增加仅改变了食真菌线虫密度和植物寄生线虫相对丰度,而食细菌线虫和捕食-杂食性线虫的密度与相对丰度在不同林龄间没有明显差异。这表明在各营养类群中,植物寄生线虫和食真菌线虫对林龄变化更敏感。在所有营养类群中,植物寄生线虫包含个体数量最多,相对丰度最高,是本研究的优势营养类群。这一结果与陈利等(2011)对水曲柳(Fraxinusmandshurica)和兴安落叶松(Larixgmelinii)人工林土壤线虫的研究一致。在陆地生态系统中,植物根系发达,这导致植物寄生线虫成为优势营养类群(Zhangetal., 2015; Wangetal., 2021)。相关分析发现,植物寄生线虫密度及相对丰度与土壤总碳、土壤碳磷比呈明显正相关。本研究中,杉木林生长到成熟林阶段受土壤磷的限制(王振宇等, 2020)。由于这种限制有利于植物光合产物向根部分配,使根系能更好吸收养分,从而增加了植物根系生物量与分泌物量(韩畅等, 2017),因此植物寄生线虫相对丰度从8年到40年呈增加趋势。在本研究中,食真菌线虫保持在较低密度。
瓦斯乐斯卡指数(WI)反映了食微生物线虫对植物寄生线虫的比例,从而指示土壤健康状况(Wasilewska, 1994)。WI值小于1,表明以植物寄生线虫为主,土壤健康状况差; WI值大于1,表明以食细菌线虫和食真菌线虫为主,土壤健康状况良好(Wasilewska, 1994)。在本研究中,林龄5年的WI值显著高于40年,表现为随林龄增加而下降的趋势,这与假设相同,反映了随杉木林龄增加,植物寄生线虫比食微生物线虫的重要性增加,土壤健康状况变差。相关分析表明,WI与土壤pH值呈正相关。由于杉木种植年限增加积累了较多的含有酸性物质的凋落物,从而导致土壤pH值下降(陈春林等, 2012)。在本研究中,土壤pH值随林龄增大总体呈下降趋势,可能不利于食微生物线虫生长。
植物寄生线虫成熟度指数(PPI)可作为评价扰动后土壤恢复状况的指标之一(Bongersetal., 1997)。PPI值的大小是由植物组织对植物寄生线虫的承载能力决定的。PPI值越大,表明植被受植物寄生线虫的侵害越严重。在本研究中,PPI值的范围为2.62~2.87,这表明杉木人工林木受到一定程度的侵害。林龄27年和40年的土壤线虫PPI显著高于21年,说明林龄21年时的林木受植物寄生线虫侵害程度相对较轻。原因可能是杉木人工林在速生阶段的林木快速生长迅速提高了对植物寄生线虫的承载能力。但随着杉木人工林林龄继续增加,林木受植物寄生线虫侵害程度加重。
5 结论
对亚热带不同林龄杉木人工林红壤的线虫群落研究表明,随着林龄增加,土壤线虫密度及多样性均无明显变化,但植物寄生线虫的相对丰度显著提高,食真菌线虫的密度降低,改变了土壤线虫的群落组成。植物寄生线虫成熟度指数和瓦斯乐斯卡指数的变化表明,杉木人工林的长期种植可能导致土壤健康状况变差。对于未来杉木人工林的长期经营,应该重点关注土壤动物尤其土壤线虫群落的变化,以提高土壤的生态系统服务功能以及人工林的生产力。
