腹腔注射亚精胺对小鼠卵巢组织多胺含量及代谢相关基因表达的影响
2017-08-07陈咨余姜冬梅龙诗韵易治鑫徐麒麟
陈咨余, 姜冬梅, 康 波, 管 成, 龙诗韵, 易治鑫, 徐麒麟
(四川农业大学 动物科技学院/畜禽遗传资源发掘与创新利用四川省重点实验室, 四川 成都 611130)
腹腔注射亚精胺对小鼠卵巢组织多胺含量及代谢相关基因表达的影响
陈咨余, 姜冬梅, 康 波, 管 成, 龙诗韵, 易治鑫, 徐麒麟
(四川农业大学 动物科技学院/畜禽遗传资源发掘与创新利用四川省重点实验室, 四川 成都 611130)
【目的】研究腹腔注射亚精胺对鼠卵巢组织多胺代谢的影响。【方法】给鼠腹腔注射不同剂量亚精胺[0(对照组、0.05、0.10和0.15 mg·g-1],应用实时荧光定量PCR检测多胺代谢关键基因表达量, 应用高效液相色谱检测鼠卵巢组织中多胺含量。【结果】注射0.15 mg·g-1亚精胺时,卵巢组织ODC、OAZ1、SPMS、SSAT、PAOX和SMOX基因表达量显著高于其他3组;注射亚精胺组鼠卵巢组织SPDS表达量均显著低于对照组;注射0.10 mg·g-1亚精胺组鼠卵巢组织中腐胺和亚精胺含量均显著高于对照组,而注射亚精胺对卵巢中精胺含量无显著影响。【结论】外源性亚精胺的腹腔注射可导致鼠卵巢组织中多胺含量以及多胺代谢相关基因表达量发生变化,且具有剂量依赖性,提示外源性亚精胺可通过介导卵巢组织的多胺代谢来参与调控卵巢功能。
亚精胺; 鼠; 卵巢; 多胺代谢; 繁殖; 基因表达; 腹腔注射
多胺(天然多胺主要包括腐胺、亚精胺和精胺)在动物繁殖过程中具有重要调控作用[1]。多胺通过影响雄性动物精子发生和精子活性来参与调控雄性动物繁殖功能[2];在雌性动物中,生殖相关激素变化可影响多胺合成限速酶鸟氨酸脱羧酶(Ornithine decarboxylase,ODC)的活性,进而导致多胺含量发生变化[2-3]。老龄鼠卵泡组织ODC活性显著低于青年鼠,且其胚胎染色体的非整倍性显著高于青年鼠,当补充外源性腐胺后,鼠胚胎染色体非整倍性显著降低,表明多胺对维持雌鼠繁殖功能具有重要作用[4-5]。
亚精胺可为真核翻译起始因子eIF5A提供氨基丁基,替换1个赖氨酸残基,使eIF5A成为具有活性的成熟蛋白分子,是eIF5A翻译后羟腐胺赖氨酸修饰过程的重要底物。eIF5A的hypusine修饰过程对于其功能和细胞增殖具有重要作用[1,6]。研究表明,外源性亚精胺对细胞免疫、生物抗逆性以及细胞增殖分化均具有显著作用,且该作用表现出剂量和时间依赖性[7]。然而,外源性亚精胺对雌性动物繁殖功能的影响尚不清楚。本研究通过给小鼠腹腔注射不同浓度亚精胺,检测鼠卵巢组织中多胺含量及多胺代谢关键基因表达量,揭示外源性亚精胺对鼠卵巢组织多胺代谢的影响,以期为多胺调控雌性动物繁殖功能的研究奠定基础。
1 材料与方法
1.1 试验动物与处理
8周龄的SPF级昆明鼠(购自成都达硕实验动物公司)32只,体质量(30±5) g随机分成4组,饲养环境为室温20~22 ℃,光照充足,给予充足的标准饲料和饮用水。按照鼠体质量计算每组鼠所需亚精胺剂量,设置对照组(生理盐水)、低剂量组(0.05 mg·g-1)、中剂量组(0.10 mg·g-1)、高剂量组(0.15 mg·g-1),每只鼠注射总剂量为300 μL,给药后自由饮水、采食,24 h后采用,采用颈椎脱臼法将鼠处死,采集卵巢组织样品,液氮冷冻后置于-80 ℃冰箱保存备用。注射用亚精胺购自Sigma公司。
1.2 实时荧光定量PCR
按照RNAiso Plus试剂盒(购自TaKaRa公司)说明书分别提取对照组和亚精胺处理组小鼠的卵巢组织中总RNA,并按照PrimeScript RT reagent Kit with gDNA Eraser试剂盒(购自TaKaRa公司)说明书制备cDNA。利用实时荧光定量PCR检测ODC、鸟氨酸脱羧酶抗酶(Ornithine decarboxylase antizyme1,OAZ1)、S-腺苷甲硫氨酸脱羧酶(S-adenosylmethionine decarboxylase,SAMDC)、亚精胺合成酶(Spermidine synthase,SPDS)、精胺合成酶(Spermine synthase,SPMS)、亚精胺/精胺N1-乙酰基转移酶(Spermine/spermine N1-acetyltransferase,SSAT)、多胺氧化酶(Polyamine oxidase,PAOX)和精胺氧化酶(Spermineoxidase,SMOX)基因表达量, 引物序列见表1。实时荧光定量PCR反应体系为:iQTMSYBR Green Supermix(购自BioRad公司) 25 μL,上、下游引物(10 μmol·L-1)各1.0 μL,cDNA模板2.5 μL,用RNaseFree水补充至50 μL。循环参数:95 ℃预变性3 min;95 ℃变性10 s,60 ℃退火30 s,72 ℃延伸30 s (采集荧光信号),40个循环。并绘制熔解曲线。每个样品进行3次重复,用GAPDH作为内参基因。
表1 实时荧光定量PCR引物信息
Tab.1 Sequences of primer pairs used in real-time fluorescent quantitative PCR
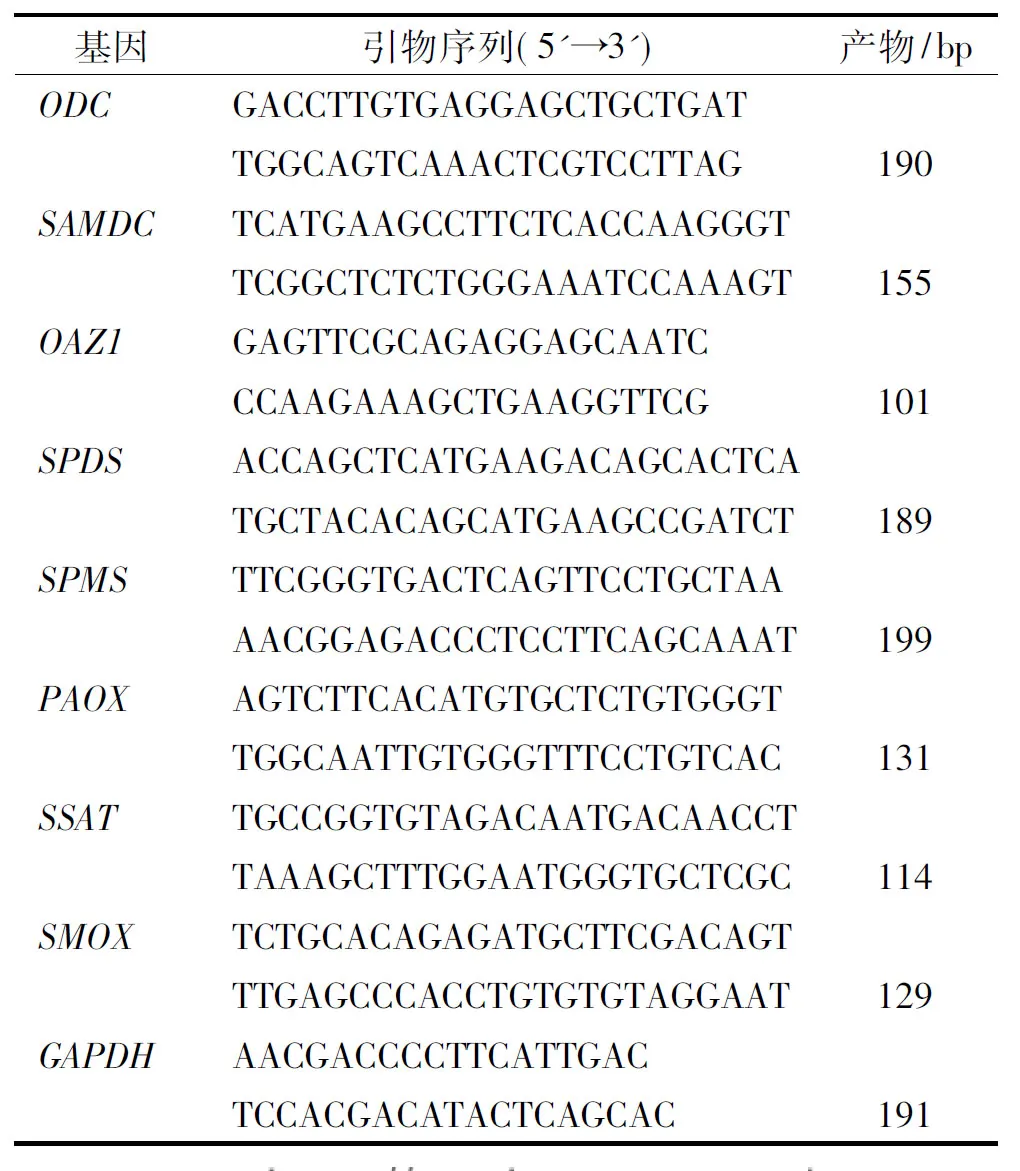
基因引物序列(5'➝3')产物/bpODCGACCTTGTGAGGAGCTGCT-GATTGGCAGTCAAACTCGTCCT-TAG190SAMDCTCATGAAGCCTTCTCAC-CAAGGGTTCGGCTCTCTGGGAAATC-CAAAGT155OAZ1GAGTTCGCAGAGGAGCAATCCCAAGAAAGCTGAAGGTTCG101SPDSACCAGCTCATGAAGACAGCACTCATGCTACACAGCATGAAGCCGATCT189SPMSTTCGGGTGACTCAGTTCCTGCTAAAACGGAGACCCTCCTTCAGCAAAT199PAOXAGTCTTCACATGTGCTCTGT-GGGTTGGCAATTGTGGGTTTCCT-GTCAC131SSATTGCCGGTGTAGACAATGACAACCTTAAAGCTTTGGAATGGGT-GCTCGC114SMOXTCTGCACAGAGATGCTTCGA-CAGTTTGAGCCCACCTGTGTGTAG-GAAT129GAPDHAACGACCCCTTCATTGACTCCACGACATACTCAGCAC191
1.3 高效液相色谱检测多胺含量
称取冻存卵巢组织,加入1.0 mLφ为 5%的HClO4,电动匀浆机匀浆,加入500 μg·mL-1的内标工作液,超声波破碎10 min,冷冻离心机(4 ℃,12 000 r·min-1)离心10 min,取上清,加入2 mL 2.5 mol·L-1的NaOH和4 μL的苯甲酰氯,漩涡震荡后置于40 ℃的水浴锅中衍生60 min。衍生液经C18固相萃取柱过柱后,用0.5 mL色谱甲醇洗脱样品,用0.22 μm针头滤器过滤洗脱液,4 ℃避光保存待检测。高效液相色谱检测条件为:流动相为V(甲醇)∶V(水)=66∶34;流速为0.9 mL·min-1;紫外检测器检测波长为229 nm;柱温为25 ℃。制作多胺标准曲线所用腐胺、亚精胺和精胺均购自Sigma公司。
1.4 数据分析
采用2-△△Ct法处理荧光定量数据,计算各处理组卵巢组织中目的基因的相对表达量。利用SAS9.2统计分析软件MEANS过程进行描述性统计分析,ANOVA过程进行方差分析和Duncan’s多重比较,试验数据用平均值±标准误表示。
2 结果与分析
2.1 鼠卵巢组织多胺合成代谢相关基因表达
由图1可知,腹腔注射不同浓度亚精胺对鼠卵巢组织中SAMDC基因表达无显著影响(P>0.05)。注射高剂量亚精胺组鼠卵巢组织中OAZ1、ODC和SPMS基因的表达量均显著高于对照组(P<0.05)。注射不同剂量亚精胺组鼠卵巢组织中SPDS基因表达量显著低于对照组,且不同剂量处理组之间SPDS表达量无显著差异。
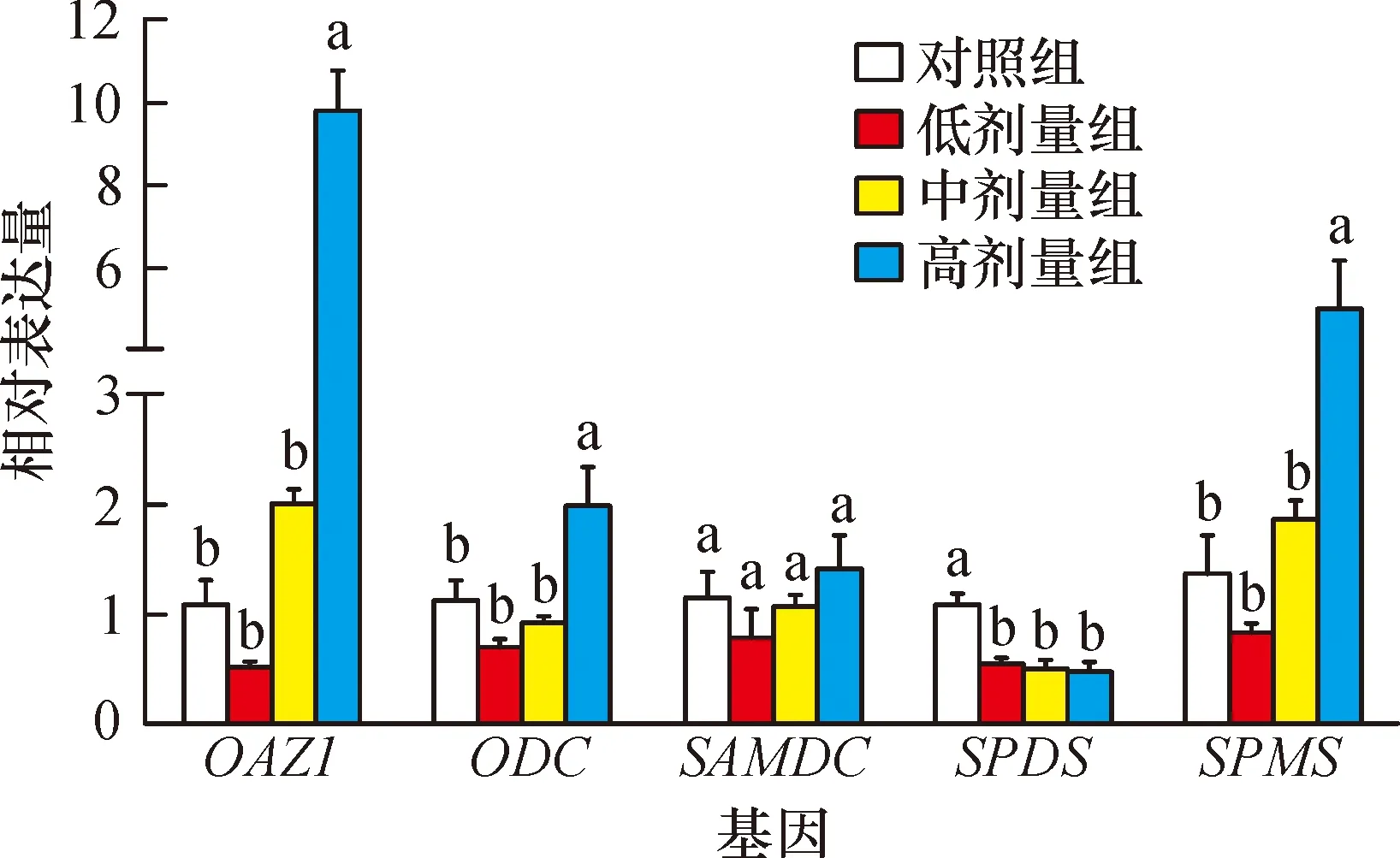
相同基因不同柱子上凡具有一个相同小写字母者,表示不同处理间差异不显著(Duncan’s法,P>0.05,n=6)。
图1 亚精胺对小鼠卵巢多胺合成代谢相关基因表达的影响
Fig.1 Effect of spermidine on polyamine biosynthesis-related gene expression in mouse ovary
2.2 鼠卵巢组织多胺分解代谢相关基因表达
由图2可知,腹腔注射低剂量亚精胺后,卵巢组织中SSAT、PAOX和SMOX表达量与对照组无显著差异;随着亚精胺剂量的逐渐升高,SSAT和SMOX表达量也逐渐升高,中剂量组表达量显著高于对照组和低剂量组,PAOX中剂量组表达量显著高于对照组;3个基因高剂量组表达量均显著高于其他3组。

相同基因不同柱子上凡具有一个相同小写字母者,表示不同处理间差异不显著(Duncan’s法,P>0.05,n=6)。
图2 亚精胺对鼠卵巢多胺分解代谢相关基因表达的影响
Fig.2 Effect of spermidine on polyamine catabolism-related genes expression in mouse ovary
2.3 鼠卵巢组织多胺水平
由图3可知,随着腹腔注射亚精胺剂量的增加,卵巢组织中腐胺的含量呈现先上升后下降的趋势,中剂量亚精胺处理组腐胺的含量显著高于对照组,低、高剂量亚精胺处理时,腐胺的含量和对照相比变化不显著。中剂量亚精胺处理组中,卵巢的亚精胺含量显著高于其他3组,高剂量组的亚精胺含量显著低于其他3组。各处理组间,卵巢中精胺含量无显著差异。
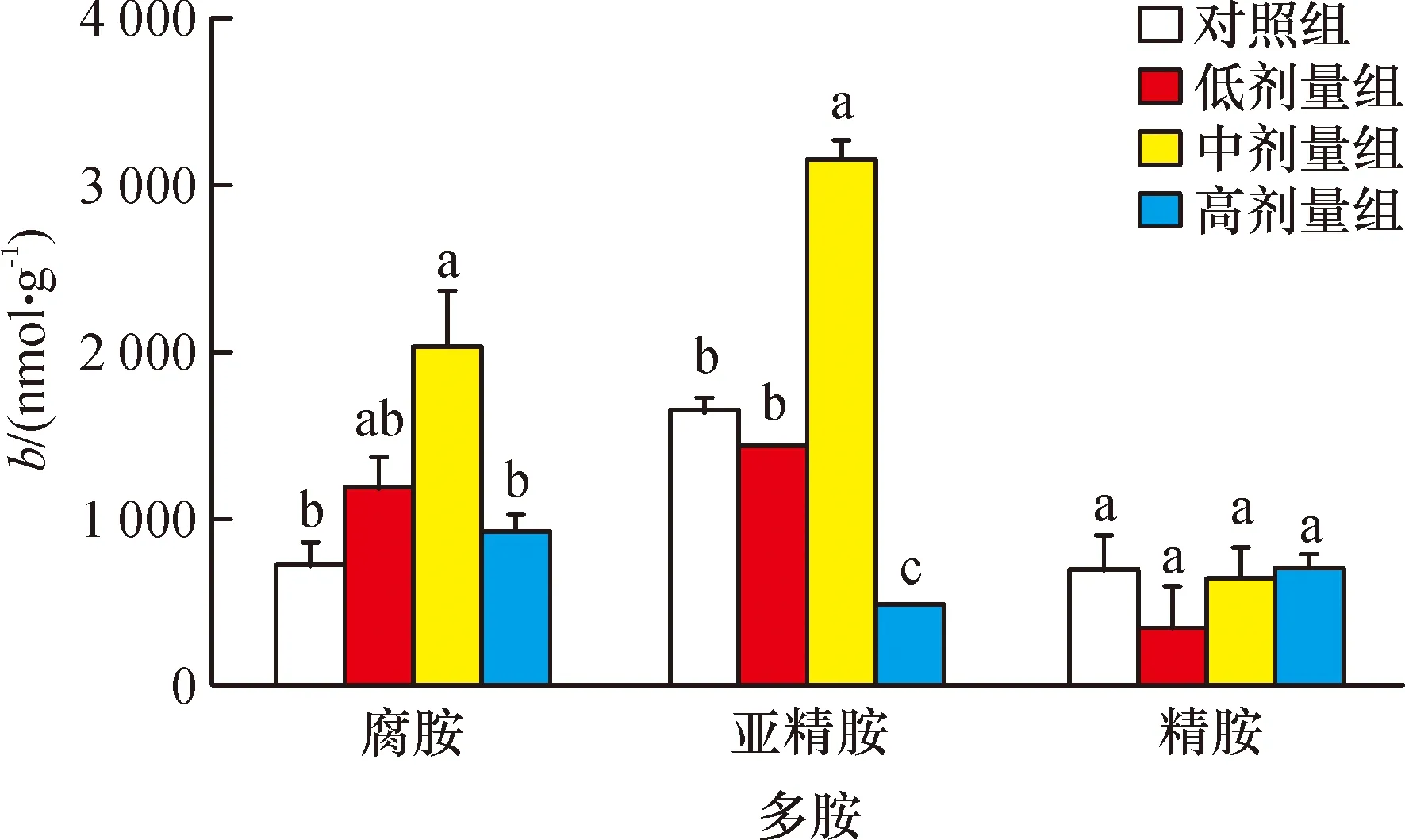
相同基因不同柱子上凡具有一个相同小写字母者,表示不同处理间差异不显著(Duncan’s法,P>0.05,n=3)。
图3 亚精胺对鼠卵巢多胺的含量的影响
Fig.3 Effect of spermidine on polyamine concentration in mouse ovary
3 讨论与结论
3.1 外源性亚精胺对鼠卵巢组织中多胺合成基因表达的影响
生物体通过调节多胺代谢过程中关键酶的表达和活性来实现对多胺水平的精密调控[8]。多胺浓度升高可触发细胞内OAZ1+1移码机制,即在多胺的刺激下,OAZ1 mRNA翻译全长功能蛋白质,降低ODC活性,进而反馈抑制多胺生物合成途径,以维持机体内多胺稳态[9]。此外,OAZ1还能影响细胞膜上的多胺转运载体,促进细胞内多胺的外排,进而降低细胞内的多胺水平[10]。腹腔注射高剂量亚精胺组鼠卵巢组织中OAZ1基因表达量显著高于对照组,提示外源性亚精胺可通过介导卵巢OAZ1+1移码机制来维持卵巢内多胺水平的稳定。高剂量的亚精胺注射小鼠后,卵巢中ODC基因的表达量显著升高,推测由于OAZ1基因的升高过度抑制了ODC的活性,机体通过上调ODC基因的表达来代偿OAZ1对ODC活性的抑制。在试验过程中,我们还发现当注射高剂量亚精胺时,鼠卵巢中SPDS基因表达量显著降低,而SPMS基因表达量显著升高,说明机体通过抑制亚精胺合成、促进亚精胺分解这2条途径共同来维持卵巢组织亚精胺水平的稳定。亚精胺注射后,处理组卵巢中SPDS基因的表达量均显著降低,推测小鼠卵巢中由于对亚精胺的吸收引起自身亚精胺含量的上升,机体一方面通过合成精胺来降低亚精胺水平,同时抑制亚精胺的合成以维持卵巢中亚精胺的正常水平。Shi等[11]研究发现,心脏过表达SAMDC基因的小鼠,心脏中亚精胺含量低于对照组,而心脏同时过表达SPDS和SAMDC基因的小鼠与仅心脏过表达SAMDC基因的小鼠相比,心脏亚精胺含量变化不显著,表明小鼠心脏中多胺的积累并非受到SPDS和SAMDC活性的调控。然而,本研究发现外源注射亚精胺对鼠卵巢SAMDC基因表达无显著影响,推测外源性亚精胺不会导致SAMDC在转录水平发生改变,可能通过翻译水平或酶的活性的变化来调控多胺水平。综上所述,外源性亚精胺主要通过上调ODC、OAZ1和SPMS基因,下调SPDS基因表达来维持机体多胺池的稳态。
3.2 外源性亚精胺对鼠卵巢组织中多胺分解代谢基因表达的影响
研究发现,多胺浓度升高可诱导核仁素亚型降解,解除其对SSAT翻译的抑制,提高SSAT活性,促进多胺代谢,从而校正多胺浓度[12]。本研究发现,随着亚精胺剂量的升高,卵巢中SSAT基因表达量逐渐升高,表明组织中多胺的分解代谢活动随着亚精胺剂量的上升而逐渐加强。注射高剂量亚精胺组鼠卵巢PAOX基因表达量显著高于其他组,提示多胺分解代谢程度可能对多胺浓度具有依赖性。精胺能够通过SMOX的作用直接转变为亚精胺,并且生成H2O2和3-氨基丙醇[13]。本研究发现,随着亚精胺注射剂量的升高,SMOX基因表达量也逐渐上升,推测由于机体内亚精胺过多导致精胺含量增加,机体为维持亚精胺/精胺比例,从而高表达SMOX基因以促进精胺分解。高表达SMOX基因可导致活性氧自由基(Reactive oxygen species,ROS)升高进而造成细胞氧化应激和DNA损伤[14]。Kwak等[15]研究表明,亚精胺和精胺氧化时产生的丙烯醛能激活细胞中抗氧化途径相关基因的表达,吴婉玲[16]研究发现,外源性精胺处理导致Jurkat细胞内精胺氧化酶活性高,增加H2O2的产生,从而造成细胞凋亡。因此,注射高剂量亚精胺鼠卵巢组织中多胺分解活动加强,多胺分解代谢产物累积,导致卵巢组织中ROS浓度升高,从而造成组织损伤。
3.3 外源性亚精胺对鼠卵巢组织中多胺水平的影响
随着亚精胺剂量的增加,鼠卵巢组织腐胺含量先升高后下降,注射0.10 mg·g-1亚精胺组鼠卵巢组织中腐胺含量最高,而0.15 mg·g-1组腐胺含量又下降至正常水平。这些结果表明,随着亚精胺剂量的增加,亚精胺向腐胺转化的效率相应增加,亚精胺优先转化成腐胺;随着亚精胺剂量进一步增加,多胺代谢基因表达量显著升高,亚精胺代谢活动增强,过多的亚精胺又在SSAT和PAOX的作用下转化为乙酰基亚精胺。注射0.10 mg·g-1亚精胺时,卵巢组织中亚精胺含量最高,推测卵巢对外源亚精胺具有一定的耐受能力。当亚精胺剂量继续增加到0.15 mg·g-1时,机体加速对亚精胺的分解,从而降低亚精胺的含量。另外,根据基因定量的检测结果,高剂量亚精胺组中SSAT、PAOX基因表达量均显著高于其余各组,提示SSAT和PAOX基因高表达促进了亚精胺的快速分解。研究表明,亚精胺和精胺分解过程中会伴随着生成ROS,后者将导致细胞功能受损[17-18]。天然存在的3种多胺中,精胺毒性最大,腐胺最小,亚精胺毒性居中[19]。本研究发现,不同剂量亚精胺注射对鼠卵巢组织中精胺含量均无显著影响,其原因可能是卵巢组织对精胺变化更加敏感,外源性亚精胺可能促使精胺合成和分解代谢活动增强,进而导致精胺合成与分解代谢活动加快,从而维持卵巢组织精胺池的稳态,然而其详细机制仍有待进一步研究阐明。
综上所述,机体内多胺含量受到多种因素影响,多胺稳态调节也是一个动态的变化过程。本研究结果表明,外源性亚精胺注射可导致鼠卵巢组织中腐胺和亚精胺含量升高;多胺合成酶OAZ1、ODC、SPDS基因表达量上升,亚精胺合成酶表达量下降;多胺分解代谢酶SSAT、PAOX和SMOX基因表达量升高,且具有剂量依赖性,提示外源性亚精胺可通过介导卵巢组织的多胺代谢来参与调控卵巢功能。
[1] PEGG A E. Functions of polyamines in mammals[J]. J Biol Chem, 2016, 291(29):14904-14912.
[2] LEFEVRE P L, PALIN M F,MURPHY B D. Polyamines on the reproductive landscape[J]. Endocr Rev, 2011, 32(5):694-712.
[3] RAMOS RDOS S, MESQUITA F S, D′ALEXANDRI F L, et al. Regulation of the polyamine metabolic pathway in the endometrium of cows during early diestrus[J]. Mol Reprod Dev, 2014, 81(7):584-594.
[4] TAO Y,LIU X J. Deficiency of ovarian ornithine decarboxylase contributes to aging-related egg aneuploidy in mice[J]. Aging Cell, 2013, 12(1):42-49.
[5] LIU D, MO G, TAO Y, et al. Putrescine supplementation duringinvitromaturation of aged mouse oocytes improves the quality of blastocysts[J]. Reprod Fertil Dev, 2016. doi:10.1071/rd16061.
[6] NISHIMURA K, LEE S B, PARK J H, et al. Essential role of eIF5A-1 and deoxyhypusine synthase in mouse embryonic development[J]. Amino Acids, 2012, 42(2/3):703-710.
[7] EISENBERG T, KNAUER H, SCHAUER A, et al. Induction of autophagy by spermidine promotes longevity[J]. Nat Cell Biol, 2009, 11(11):1305-1314.
[8] NOWOTARSKI S L, WOSTER P M,CASERO R A JR. Polyamines and cancer: Implications for chemotherapy and chemoprevention[J]. Expert Rev Mol Med, 2013, 15:e3.
[9] RATO C, AMIROVA S R, BATES D G, et al. Translational recoding as a feedback controller: Systems approaches reveal polyamine-specific effects on the antizyme ribosomal frameshift[J]. Nucleic Acids Res, 2011, 39(11):4587-4597.
[10]PETROS L M, HOWARD M T, GESTELAND R F, et al. Polyamine sensing during antizyme mRNA programmed frameshifting[J]. Biochem Biophys Res Commun, 2005, 338(3):1478-1489.
[11]SHI C, WELSH P A, SASS-KUHN S, et al. Characterization of transgenic mice with overexpression of spermidine synthase[J]. Amino Acids, 2012, 42(2/3):495-505.
[12]PEREZ-LEAL O, BARRERO C A, CLARKSON A B, et al. Polyamine-regulated translation of spermidine/spermine-N1-acetyltransferase[J]. Mol Cell Biol, 2012, 32(8):1453-1467.
[13]CHATURVEDI R, DE SABLET T, PEEK R M, et al. Spermine oxidase, a polyamine catabolic enzyme that linksHelicobacterpyloriCagA and gastric cancer risk[J]. Gut Microbes, 2012, 3(1):48-56.
[14]AVERILL-BATES D A, KE Q, TANEL A, et al. Mechanism of cell death induced by spermine and amine oxidase in mouse melanoma cells[J]. Int J Oncol, 2008, 32(1):79-88.
[15]KWAK M K, KENSLER T W,CASERO R A JR. Induction of phase 2 enzymes by serum oxidized polyamines through activation of Nrf2: Effect of the polyamine metabolite acrolein[J]. Biochem Biophys Res Commun, 2003, 305(3):662-670.
[16]吴婉玲. 探讨多胺诱发血癌细胞凋亡的作用机制[D]. 台湾:高雄医学大学生物化学研究所, 2013.
[17]CERVELLI M, PIETROPAOLI S, SIGNORE F, et al. Polyamines metabolism and breast cancer: State of the art and perspectives[J]. Breast Cancer Res Treat, 2014, 148(2):233-248.
[18]YANG L, ZHAO J, ZHU Y, et al. Reactive oxygen species (ROS) accumulation induced by mononaphthalimide-spermidine leads to intrinsic and AIF-mediated apoptosis in HeLa cells[J]. Oncol Rep, 2011, 25(4):1099-1107.
[19]TURECKI G. Polyamines and suicide risk[J]. Mol Psychiatry, 2013, 18(12):1242-1243.
【责任编辑 庄 延】
Effects of intraperitoneal spermidine injection on polyamine content and metabolism-related gene expression in mouse ovary
CHEN Ziyu, JIANG Dongmei, KANG Bo, GUAN Cheng, LONG Shiyun, YI Zhixin, XU Qilin
(College of Animal Science and Technology, Sichuan Agricultural University /Farm Animal Genetic Resources Exploration and Innovation Key Laboratory of Sichuan Province, Chengdu 611130, China)
【Objective】 To observe the effects of intraperitoneal injection of spermidine on polyamine metabolism in mouse ovary.【Method】 Mice were injected with different doses of spermidine[0(control), 0.05, 0.10 and 0.15 mg·g-1]. Real-time fluorescent quantitative PCR was used to determine the ovarian expression levels of key genes in polyamine metabolism. High performance liquid chromatography (HPLC) was used to detect the polyamine levels in mouse ovarian tissues.【Result】 After administration of 0.15 mg·g-1spermidine, the expression ofODC,OAZ1,SPMS,SSAT,PAOXandSMOXgenes in ovarian tissues were significantly higher compared to other three groups.SPDSgene expressions of all three spermidine administration groups were significantly lower compared to control. After administration of 0.10 mg·g-1spermidine, the putrescine and spermidine contents in mouse ovary were significantly higher compared to control, while spermidine administration had no significant effect on spermine content in ovary. 【Conclusion】 Exogenous spermidine injected intraperitoneally can change polyamine content and polyamine metabolism-related gene expression in mouse ovary. Such effects are dose-dependent. Exogenous spermidine can potentially participate in the regulation of ovarian function through adjusting polyamine metabolism.
spermidine; mouse; ovary; polyamine metabolism; reproduction;gene expression; intraperitoneal injection
2016- 09- 01 优先出版时间:2017- 06-21
陈咨余(1993—),女,硕士研究生,E-mail: oleander0809@163.com; 通信作者:姜冬梅(1978—),女,讲师,E-mail: jiangdm9277@163.com;康 波(1978—),男,副教授,博士,E-mail: bokang@sicau.edu.cn
国家自然科学基金(31201798);高等学校博士学科点专项科研基金资助课题(20105103120003)
S814.1
A
1001- 411X(2017)04- 0052- 05
优先出版网址:http://kns.cnki.net/kcms/detail/44.1110.s.20170621.1924.018.html
陈咨余, 姜冬梅, 康 波, 等.腹腔注射亚精胺对小鼠卵巢组织多胺含量及代谢相关基因表达的影响[J].华南农业大学学报,2017,38(4):52- 56.
