我国森林土壤微生物生物量碳的纬度分布特征及影响因子*
2022-05-11马寰菲王洁莹任成杰郭垚鑫赵发珠
周 莎 马寰菲 王洁莹 任成杰 郭垚鑫 王 俊,4 赵发珠
(1.西北大学城市与环境学院 陕西省地表系统与环境承载力重点实验室 西安 710127; 2.西北农林科技大学农学院 杨凌 712100; 3.西北大学生命科学学院 西安 710127; 4.中国科学院水利部水土保持研究所 杨凌 712100)
土壤微生物生物量碳(microbial biomass carbon,MBC)含量占土壤有机碳含量的0.5%~13%(Heetal., 2020),可灵敏反映环境和气候的变化,也可直接表征微生物群落状态与功能变化(何越等, 2021),是评价土壤质量的重要指标(Mazzarinoetal., 1991)。在森林生态系统中,土壤微生物生物量碳是有生物活性的养分,一方面作为相对易分解和周转较快的营养库,对碳、氮、磷等化学元素的生物地球化学循环起着重要的驱动作用(李品等, 2019),另一方面作为活性碳库的一部分,对碳的固持和累积具有重要意义(Tayloretal., 1999)。虽然土壤微生物生物量碳含量占土壤有机碳含量比例相对较小,但是对森林生态系统碳库平衡和养分循环意义重大。
森林生态系统中关于微生物生物量碳的研究已有大量报道。例如,曹润(2019)等研究表明,热带森林中土壤微生境(温度与水分)时空变化及土壤碳/氮养分状况对土壤微生物生物量碳含量有显著影响。Xu等(2006)对温带火山森林土壤的研究表明,MBC含量与树种类型、气温、土壤碳氮比、土壤酸性具有一定相关性。此外,温带森林的冻融循环(高珊等, 2018)和降水量变化(方熊等, 2012)、暖温带天然林的土层变化(Anaetal., 2011)等对MBC含量也有较大影响。Tan等(2008)研究表明,寒带针阔混交林中的MBC含量对土壤压实度和地表植物残体去除的响应较敏感。目前,微生物生物量碳含量在不同森林类型中受不同因素影响,即便在相同土壤、气候条件下,不同森林类型土壤微生物生物量碳含量也存在较大差异。在森林土壤微生物生物量碳变化规律研究中,明确其影响因素很重要。
森林土壤微生物生物量碳具有较强的温度敏感性(吴明辉等, 2021); 也有研究表明,植被类型(Bergetal., 2009)、凋落物多样性(Marceletal., 2008)、土壤理化性质(Salvadoretal., 2017)、土壤有机碳化学结构的差异(Rumpeletal., 2002)等都会对土壤微生物生物量碳造成影响。植被类型变化导致凋落物、根系分泌物等外源有机碳输入产生差异,进而影响土壤有机碳化学结构组分的变化(Quideauetal., 2001; 孙筱璐, 2017)。在人工林中,土壤微生物生物量的差异主要受不同树种之间凋落物烷基碳组分的差异影响(张冰冰等, 2021)。
目前森林土壤微生物生物量碳的研究多集中于单一林分,对沿纬度梯度变化的气候、土壤理化性质等引起的森林微生物生物量碳含量变化研究还较少。本研究以我国沿纬度梯度分布的10个典型森林生态系统为对象,通过分析表层(0~10 cm)土壤MBC含量,结合气候和土壤理化性质多个因素,探讨土壤MBC含量的纬度变化规律及其驱动因子,以期揭示我国沿纬度分布的森林生态系统中土壤MBC含量变化的效应及机制,为气候变暖条件下深入理解森林土壤MBC周转和固持提供理论依据。
1 研究区概况
选择具区域代表性的10个典型森林生态系统,从北到南依次是帽儿山、东灵山、富县、火地塘、茂县、贡嘎山、哀牢山、西双版纳、木论和尖峰岭。该森林样带有明显的水热梯度变化,年均气温3.1~23.15 ℃,年均降水量486~2 266 mm,气候和植被类型丰富。样带概况见表1。
2 研究方法
2.1 样点设置及样品采样
2019年7—8月进行样地设置和样品采集。在每个森林生态系统中选取区域代表性强且植被类型相对单一、人为干扰小的森林站点设置样地,每个森林生态系统中选取3块标准样地(50 m×50 m),在每块样地内用内径5 cm的土钻按“S”型采集0~10 cm表层土样9个,将同一样地内9个土样均匀混合。剔除石块、根系、凋落物、动植物残体等,过2 mm土壤筛后将样品一分为二:一部分低温保存带回实验室,置于-20 ℃冰箱保存,用于测定土壤微生物生物量碳含量和土壤酶活性;另一部分用塑封袋装好、常温带回实验室,自然风干、粉碎、过筛(0.25 mm)后,用于测定其他土壤理化性质。

表1 森林样带基本信息
2.2 土壤理化性质测定
土壤有机碳含量用H2SO4-K2Cr2O7外加热法测定;全氮含量用H2SO4凯氏定氮法测定;全磷含量用紫外分光光度计比色法测定(鲍士旦, 2000)。
土壤MBC含量用氯仿熏蒸-硫酸钾浸提法测定(Vanceetal., 1987)。将经过熏蒸和未熏蒸的新鲜土壤(10 g)分别用0.5 mol·L-1的K2SO4溶液浸提,使用TOC-VCPH仪测定浸提液的有机碳质量分数。利用下述公式求土壤微生物生物量碳含量(MBC,单位mg·kg-1):
MBC=(EC-EC0)/0.38。
式中:EC和EC0分别为熏蒸和未熏蒸土样的浸提液碳含量(mg·kg-1),0.38为校正系数(Vanceetal., 1987)。
采用微孔板荧光法(Saiya-Corketal., 2002)测量土壤中β-葡萄糖苷酶(β-1,4-glucosidase,BG)和纤维二糖水解酶(cellobiohydrolase,CBH)活性。用吸光光度法测定多酚氧化酶(polyphenol oxidase,PPO)和过氧化物酶(peroxidase,PER)活性。
氧化酶(Oy-EEAs)活性是PER和PPO活性之和,水解酶(Hy-EEAs)活性是BG和CBH活性之和。
采用13C固体核磁共振法测定土壤有机碳化学结构组分含量,主要包括土壤烷基碳组分含量,土壤含氧烷基碳组分含量,土壤芳香基碳组分含量,土壤羧基碳组分含量和土壤羰基碳组分含量(Kögel-Knabner, 1997)。
利用pH计(PHS-3C,中国上海)测量去离子水悬浮液(水土比为2.5∶1)中的土壤pH值,环刀法(100 cm3)测定土壤密度(鲍士旦, 2000)。用激光分散法测定土壤质地(Mccaveetal., 1986)。
2.3 数据处理
利用SPSS18.0软件处理数据。用单因素方差分析(ANOVA)对不同纬度梯度森林土壤MBC含量和土壤有机碳化学结构组分含量进行差异性检验。用Mantel test对影响MBC含量变化的因素进行相关性分析,评估土壤MBC含量沿纬度变化的主要影响因素。使用Origin 9.5软件绘图。
3 结果分析
3.1 土壤微生物生物量碳含量沿纬度梯度的变化
MBC含量呈沿纬度降低而降低的趋势(图1),在帽儿山最大,为(913.32±39.62 )mg·kg-1,西双版纳最小,为(200.57±13.99)mg·kg-1,最大值是最小值的4.6倍。但处于低纬度的木论的土壤MBC含量显著高于同样处于低纬度的尖峰岭和西双版纳,并显著高于火地塘、茂县、贡嘎山和哀牢山(P<0.05)。
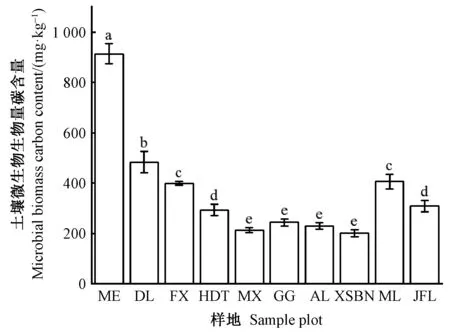
图1 中国森林样带土壤微生物生物量碳含量的变化
3.2 土壤有机碳化学结构组分含量沿纬度梯度的变化
不同有机碳化学结构组分含量沿纬度呈现不同变化规律(图2)。烷基碳含量在火地塘最大(13.7%±1.8%),在茂县最小(1.9%±0.3%)。含氧烷基碳含量最大值出现在西双版纳(38.0%±0.7%),最小值出现在木论(26.5%±1.3%)。芳香基碳含量变化在59.7%±0.2%和44.0%±0.3%之间,最大值和最小值分别出现在木论和贡嘎山。羧基碳含量在贡嘎山最大(8.9%±0.6%),在哀牢山最小(1.7%±1.7%)。羰基碳含量在茂县最大(3.3%±1.6%),在木论最小(0.7%±0.3%),且茂县羰基碳含量显著高于其他各组(P<0.05)。
3.3 土壤微生物生物量碳含量与非生物因素的相关性分析
Mantel test结果(表2)表明:MBC含量与土壤理化性质指标中的有机碳含量(r=0.64,P<0.01)、砂砾含量(r=0.48,P<0.01)、水解酶活性(r=0.48,P<0.01)、磷含量(r=0.47,P<0.01)、pH值(r=0.43,P<0.01)、氮含量(r=0.31,P<0.01)和碳化结构中的烷基碳含量(r=0.21,P<0.01)均极其显著正相关,与粉粒含量(r=0.15,P<0.05)、氧化酶活性(r=0.15,P<0.05)显著正相关,与土壤密度、黏粒含量及其他有机碳化学结构组分含量无显著相关。在气候条件中,MBC含量与年均气温极显著正相关(r=0.31,P<0.01),与年均降水量无显著相关。

图2 中国森林样带土壤有机碳化学结构组分含量变化

表2 土壤微生物生物量碳含量与气候及土壤理化性质的相关性
4 讨论
4.1 森林土壤MBC含量沿纬度的分布特征
本研究发现,MBC含量随纬度降低而降低(图1),与王春燕(2016)等的研究结果基本一致。MBC含量沿纬度的变化范围为(200.57±13.99)~(913.32±39. 62)mg·kg-1,与全球森林土壤微生物生物量碳含量(347±125)~(971±650)mg·kg-1基本一致(李品等, 2019)。MBC含量在帽儿山、东灵山、富县、火地塘、茂县、贡嘎山、哀牢山、西双版纳、木论和尖峰岭间存在较大差异,有明显的纬度梯度。先前研究也证明土壤MBC含量与多种因素有关,如吴明辉等(2021)研究认为高寒多年冻土区MBC含量受土壤温度、地下生物量和pH值影响。刘宝等(2019)在中亚热带森林生态系统研究中发现,凋落物数量和质量、土壤有机碳含量和土壤温度是导致各林分类型土壤MBC含量差异的主要因素。需要指出的是,本研究发现处于低纬度的木论喀斯特地区的MBC含量较高,这可能与喀斯特地区的森林类型及特殊地理条件有关。木论喀斯特为中亚热带石灰岩常绿落叶阔叶混交林,是森林植被顶级群落分布区(陈香碧等, 2018)(表1),其地上、地下有机物质输入量较大,在一定条件下,有机质输入量越多,土壤MBC含量越高(Linetal., 1999); 另一方面,喀斯特地区钙质丰富的石灰土(表1)环境为细菌、放线菌等微生物提供了异常良好的活动场所,丰富的凋落物进入土壤并不断被分解形成腐殖质,并与土壤中钙、镁等离子结合,形成高度缩合稳定的腐殖质钙,促进碳的积累,也更利于MBC含量的积累(李新爱等, 2006; 徐学池等, 2019)。除此之外,喀斯特地区高异质性的微生境(龙健等, 2021)、特殊的二元水文结构和复杂的地貌形态引起的土壤养分的多元性、高变异性等(范夫静等, 2014; 徐广平等, 2019),均影响土壤微生物及MBC的积累过程。
4.2 影响森林土壤MBC含量纬度变化的因素
土壤理化性质是驱动MBC含量沿纬度变化的主要因子(表2)。土壤理化性质作为微生物生存繁殖的重要环境条件,对微生物群落生长、MBC积累具有重要调控作用。本研究表明,土壤有机碳含量、全氮含量和全磷含量与MBC含量呈极显著正相关(P<0.01)(表2)。Monika等(2021)研究表明,温带森林土壤MBC含量与土壤有机碳含量具有显著正相关关系(P<0.05); 王全成等(2021)研究发现,亚热带森林土壤MBC含量与全氮含量、全磷含量均呈显著正相关(P<0.05),这与本研究结果一致。土壤MBC含量是土壤养分的供给源和储备库(Gregorichetal., 2000),土壤有机碳含量、全氮含量和全磷含量可直接影响土壤MBC含量。土壤有机碳、氮和磷是微生物生长的重要营养和能量来源(王全成等, 2021),土壤有机碳、全氮和全磷含量变化会影响微生物活性及其生物量。吴旭东等(2021)研究表明,在极端干旱条件下,盐池荒漠草原土壤MBC含量的积累,是微生物和植物竞争土壤中碳、氮养分的结果。土壤砂砾含量(P<0.05)、粉粒含量(P<0.01)分别与MBC含量显著和极显著相关(表2),这是因为土壤质地随纬度变化而变化,这种变化部分受不同纬度的植被类型差异影响(Wenetal., 2020),不同植被类型的根系机械作用、分泌物胶结土壤团聚体、凋落物输入等过程的差异会影响土壤质地(Xiaetal., 2020),而土壤质地影响MBC的固持(王群艳等, 2016)。除此之外,pH值也是影响MBC含量纬度分布的重要因素(P<0.01)(表2),这可能与pH值影响微生物活性及微生物储存碳的能力有关(Maliketal., 2018)。
本研究发现,土壤水解酶和氧化酶活性分别与MBC含量极显著(P<0.01)和显著正相关(P<0.05)(表2)。以往研究也表明,MBC含量与土壤胞外酶活性(解梦怡等, 2020)存在密切关系。在土壤的众多生化反应中,土壤胞外酶是有机碳分解速率的重要限速因子(Xuetal., 2017)。高纬度地区的低温低湿环境、针叶树种根系分泌物、凋落物中含量较高的难降解成分等资源限制及环境压力,迫使微生物在种群生长过程中将细胞投资侧重于压力耐受性和资源获取(以胞外酶生产的形式降解复杂的底物)方面,提高了生理策略的投资,减缓了碳分解速率(Maliketal., 2018)。相反,低纬度地区的高温湿润环境,阔叶树种根系分泌物及凋落物中较高含量的水溶性糖、加尼奇酸和氨基酸等易被微生物利用的养分(Prihaetal., 2001),土壤动物的频繁活动,导致微生物活性高,微生物以合成和分泌降解代谢产物、防御代谢产物为主(Bollmannetal., 2009),合成生物量碳次之(Liangetal., 2017 )。
另外,本研究发现,气候条件中年均气温对MBC含量的纬度分布有极显著影响(P<0.01)(表2)。这可能是因为温度沿纬度有巨大分异(表1),温度变化调控土壤中微生物群落的活性(刘纯等, 2014),当温度较高时,土壤微生物活性强、代谢速率高、生长繁殖旺盛(Brownetal., 2004),MBC的积累速率不足以抵消分解消耗速率,从而造成MBC含量降低。Fang 等(2020)得出亚热带森林土壤温度升高会引起土壤MBC含量降低的结论;吴明辉等(2021)认为高寒地区土壤MBC含量与土壤温度显著负相关,都印证了本研究结果。
本研究结果表明,土壤有机碳化学结构组分中烷基碳含量与MBC含量极显著正相关(P<0.01)(表2)。这与孙筱璐(2017)等在南亚热带、暖温带和温带3种典型森林的研究结果不一致。主要原因是纬度变化引起的植被类型不同导致土壤有机碳化学结构有所差异(Quideauetal., 2001)。一方面,森林生态系统中,丰富多样的植被类型及源源不断的凋落物、根系分泌物等外源有机碳的输入,促进了微生物对土壤中活性有机碳的利用,MBC作为活性有机碳源被微生物分解,烷基碳因难降解而富集(Nicoleetal., 2003)。另一方面,土壤烷基碳来自于微生物代谢产物和木栓质、角质等聚合物(Douetal., 2008),难以降解,较为稳定。相对于低纬度地区的阔叶林,高纬度地区针叶林的土壤中烷基碳含量较高(Chenetal., 2004),微生物活性相对较低(陈仕奇等, 2019),微生物分解利用活性碳源速率较慢,影响MBC的积累。
5 结论
森林土壤MBC含量有明显的纬度分布规律,随纬度的降低而降低。土壤质地、土壤营养和土壤酶活性和土壤烷基碳含量是森林土壤MBC含量纬度分布格局的主控因素,年均气温是次要影响因素。
