不同林龄杉木林土壤磷形态与解磷菌变化*
2022-05-11王淑真梁晶晶包明琢周垂帆
王淑真 梁晶晶 包明琢 潘 菲 周垂帆
(福建农林大学林学院 人工林可持续经营福建省高校工程研究中心 福州 350002)
第九次全国森林资源清查数据显示,我国杉木(Cunninghamialanceolata)人工林面积达1 138.66万hm2,占全国乔木林面积的6.33%(孙长山, 2020),其在调节气候、涵养水源和提供用材等方面发挥着重要作用。由于育林措施和经营制度不甚合理,当前杉木人工林面临土壤退化、肥力下降、生产力降低等一系列问题(田甜等, 2019)。
磷素是植物生长不可缺少的大量元素。在短时间内,磷素主要来源于植物凋落物分解和有机质矿化,在土壤中迁移与转化后,被植物吸收利用或以可溶性磷的形式流失(陈美领等, 2016)。在亚热带森林生态系统中,土壤总磷含量较高,但其中闭蓄态磷所占比例较大,可被林木吸收的有效磷含量处于较低水平。作为重要速生树种,杉木产生凋落物较少且难分解,养分归还量少,同时酸性红壤中的铁、铝氧化物对磷素的固定吸附作用强烈,土壤磷素不足成为限制杉木生长的主要因子(Zhangetal., 2018)。研究表明,土壤磷素含量和微生物利用特征等与林龄联系紧密。曹小玉等(2019)发现随杉木林龄增加,土壤全磷含量先减后增,中龄林时最低。Wu等(2020)的研究表明,杉木人工林土壤磷素生物有效性和循环利用率随林龄增加而增大。
微生物活动与土壤磷素循环密切相关,对磷素形态的转化和有效性有极重要影响。土壤中存在一类可将难溶性磷酸盐转变为可供植物利用的可溶性磷化合物的微生物,称为解磷菌(骆韵涵等, 2020)。众多研究表明,解磷菌可通过自身生命活动产生质子、CO2等,降低土壤pH值,引起无机磷酸盐溶解; 通过产生酶类水解有机磷; 通过产生有机酸,溶解难溶的磷酸盐,或争夺吸附位点等,对土壤磷素进行活化(滕泽栋等, 2017)。如Churgopost等(2021)发现施加解磷菌生物肥料提高了Takhli土系中磷素的有效性。然而,以往研究多聚焦在杉木林土壤养分循环特征、理化性质和微生物群落多样性的变化,以及施加解磷菌生物肥料对土壤磷素有效性的影响,自然条件下不同林龄杉木纯林土壤中解磷菌群落组成和磷素形态转化规律的研究鲜有报道。
碱性磷酸酶(ALP)可催化酯-磷酸键的水解(Nannipierietal., 2011),主要由土壤细菌、真菌及土壤动物(蚯蚓等)分泌(Nakasetal., 1987),可将土壤有机磷水解为可供植物利用的无机磷。ALP活性与phoD基因丰度呈显著正相关(Wuetal., 2015)。在当前已测序的编码ALP的3种基因(phoA、phoD、phoX)中,phoD基因在土壤样品分析中较常见(Tanetal., 2013),已成为研究陆地生态系统土壤有机磷转化的关键生物指示物。目前,利用高通量测序对土壤中解磷菌的种类、丰度、分布状况和群落组成进行分析,有助于了解人工林不同生长阶段的土壤微生物群落结构以及土壤功能变化。
鉴于此,本文基于高通量测序技术对PCR扩增的phoD基因进行测定,对福建省南平市杉木人工林生长过程中不同分类水平上解磷菌群落结构及多样性进行研究,以揭示土壤解磷菌群落结构和多样性随林龄的变化规律及磷素形态与不同分类水平上解磷菌群落的相关性,为杉木人工林磷素有效利用和可持续经营提供参考。
1 材料与方法
1.1 研究区概况与样品采集
福建省南平市是我国杉木中心产区之一,地处亚热带季风气候区,年均温19.3 ℃,相对湿度83%,土壤是由花岗岩发育的红壤。土样采自南平市溪后村(26°39′N,117°55′E),采样时间为2018年7月27、28日。采用空间代时间法,选择立地相似的不同林龄(4、15、24、43、100年)杉木人工林。其中,4年为幼龄林,15年为中龄林,24年为近熟龄,43年和100年为过熟林。乔木层以杉木为主,不同林龄杉木林的林下植被种类和覆盖度不同。4年林下植被覆盖度较低,以芒萁(Dicranopterisdichotoma)为主; 15年和24年林下植被丰富度上升,灌木层主要有山苍子(Litseacubeba)、毛冬青(Ilexpubescens)、盐肤木(Rhuschinensis)等,草本层主要有芒萁、乌毛蕨(Blechnumorientale)等; 43年和100年的林下植被更为丰富,灌木层主要有粗叶榕(Ficushirta)、毛冬青等,草本层主要有观音莲座蕨(Angiopterisfokiensis)、淡绿短肠蕨(Allantodiavirescens)等。每个林龄选择坡度和其他立地条件相近的3片林地作为重复,每片林地选择坡度30°~40°、坡向东南的土坡进行采样。每个土坡上划定20 m×20 m的采样地,沿S形选择6个采样点,在去除地表枯枝落叶层后,用土钻采集0~20 cm的表层土壤,并将采集点的土样等量混合为1份样品(张芸等, 2016)。每份土壤样品过20目筛后放入4 ℃的冰箱中冷藏保存,一部分用于土壤磷素形态检测,一部分用于解磷菌相对含量测定。
1.2 土壤磷素测定
土壤磷组分测定采用改进的Hedley磷素分级法(Guoetal., 1998; Börlingetal., 2001)。称取0.5 g土样(< 2 mm)于50 mL离心管中,逐级加入蒸馏水、0.5 mol·L-1NaHCO3、0.1 mol·L-1NaOH、1 mol·L-1HCl溶液30 mL后,振荡16 h,4 500 r·min-1离心后取上清液。取浓硫酸和0.5 mol·L-1的H2O2溶液,将剩余土样在270 ℃下消煮,测定Residual-P。活性磷包括水溶性无机磷(H2O-inorganic phosphorus, H2O-IP),水溶性有机磷(H2O-organic phosphorus, H2O-OP),碳酸氢钠态无机磷(NaHCO3-inorganic phosphorus, NaHCO3-IP)和碳酸氢钠态有机磷(NaHCO3-organic phosphorus, NaHCO3-OP)。中等活性磷包括氢氧化钠态无机磷(NaOH-inorganic phosphorus, NaOH-IP)和氢氧化钠态有机磷(NaOH-organic phosphorus, NaOH-OP)。稳定态磷包括盐酸态无机磷(HCl-inorganic phosphorus, HCl-IP),盐酸态有机磷(HCl-organic phosphorus, HCl-OP)和Residual-P。土壤无机磷总量TIP(Total inorganic phosphorus)=H2O-IP+NaHCO3-IP+NaOH-IP+HCl-IP+Residual-P,有机磷总量TOP(Total organic phosphorus)=H2O-OP+NaHCO3-OP+NaOH-OP+HCl-OP。
1.3 土壤解磷菌测定
基因组DNA抽提完成后,利用1%琼脂糖凝胶电泳检测抽提的基因组DNA,采用引物F733(5′-TGGGAYGATCAYGARGT-3′)和R1083(5′-CTGSGCSAKSACRTTCCA-3′)(Chenetal., 2019)对其中的碱性磷酸酶基因(phoD)部分进行PCR扩增。每个样本3个重复,将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测,使用AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)切胶回收PCR产物。回收的产物经过连接 “Y”字形接头、去除接头自连片段、再次使用PCR进行文库模板的富集、氢氧化钠变性,产生单链DNA片段之后,完成高通量测序文库的制备。将符合要求的测序文库在Illumina Miseq平台上进行双末端测序(Paried-End),对基因进行物种注释后得到各分类水平上的解磷菌的相对丰度。
1.4 数据分析
根据OTU丰度信息绘制Venn图,可较直观地显示不同林龄杉木林土壤中共有与特有的OTU数目。为研究组间群落组成的异同,进行主坐标分析(PCoA)和利用Qiime软件绘制基于bray curtis的多样本聚类树。通过LefSe分析,进一步分析组间丰度上有显著差异的物种。通过Microsoft Excel 2006初步整理土壤磷组分原始数据,制作不同形态磷素含量三线表和面积图。使用SPSS对组间土壤磷素含量和解磷菌Alpha多样性指数分别进行单因素方差分析,选择SNK法进行显著性检验,将土壤磷素含量和微生物Alpha多样性指数进行双变量相关分析。使用Origin 2018绘制解磷菌Alpha多样性折线图。土壤磷素含量及解磷菌群落相对丰度冗余分析(RDA)使用Canoco 4.5。
2 结果与分析
2.1 不同林龄杉木林土壤磷素形态特征
由图1可知,林龄对土壤中不同形态磷素的含量影响显著。随林龄增加,活性磷含量逐渐增大。有机磷总量(TOP)随林龄变化表现为100年>24年>43年>15年>4年。总无机磷(TIP)和稳定态的磷含量变化趋势相同,即15年>4年>24年>100年>43年。

图1 不同林龄杉木林土壤有机磷、无机磷和不同活性磷素含量
由表1可知,不同形态磷素含量随林龄增加的变化趋势不同。在同一林龄,含量较高的磷素分别是Residual-P、NaOH-IP、NaHCO3-IP和NaOH-OP。土壤中H2O-IP含量随林龄增加而显著升高(P<0.05); NaOH-IP含量在24年时最大; NaHCO3-IP含量随林龄增加先升后降,在43年时最大。NaOH-OP含量随林龄变化明显,24年时较15年提升了78.69%,在100年时含量显著高于其他林龄(P<0.05); HCl-OP含量随林龄增加整体上逐渐增大。
2.2 不同林龄杉木林土壤中解磷菌绝对丰度分析
根据林龄分5组进行Venn图分析,共产生11 939个OTU,经抽平处理后剩余11 026个,其中4、15、24、43、100年中平均包含1 784、1 878、2 066、3 097、2 347个,4、15、24、43、100年中特有的OTU分别为1 262、594、830、2 524、1 284个,5组共同包含471个。不同林龄土壤包含的解磷菌OTU数目为43年>100年>24年>15年>4年。Venn图分析结果显示,43年林龄包含的解磷菌类群独特性最明显。整体来看,不同林龄杉木林土壤中的解磷菌种类差异显著。
表1 不同林龄杉木林土壤磷素形态特征①

图2 不同林龄杉木林土壤解磷菌OTUs分布
PCoA中样品点的距离可用来估计组间解磷菌群落结构与多样性的差异,样品点距离越近说明越相似,距离越远说明差异越大。15年和24年林龄土壤中解磷菌群落结构和多样性相近,4年林龄土壤中解磷菌群落与其他林龄差异较大(图3)。
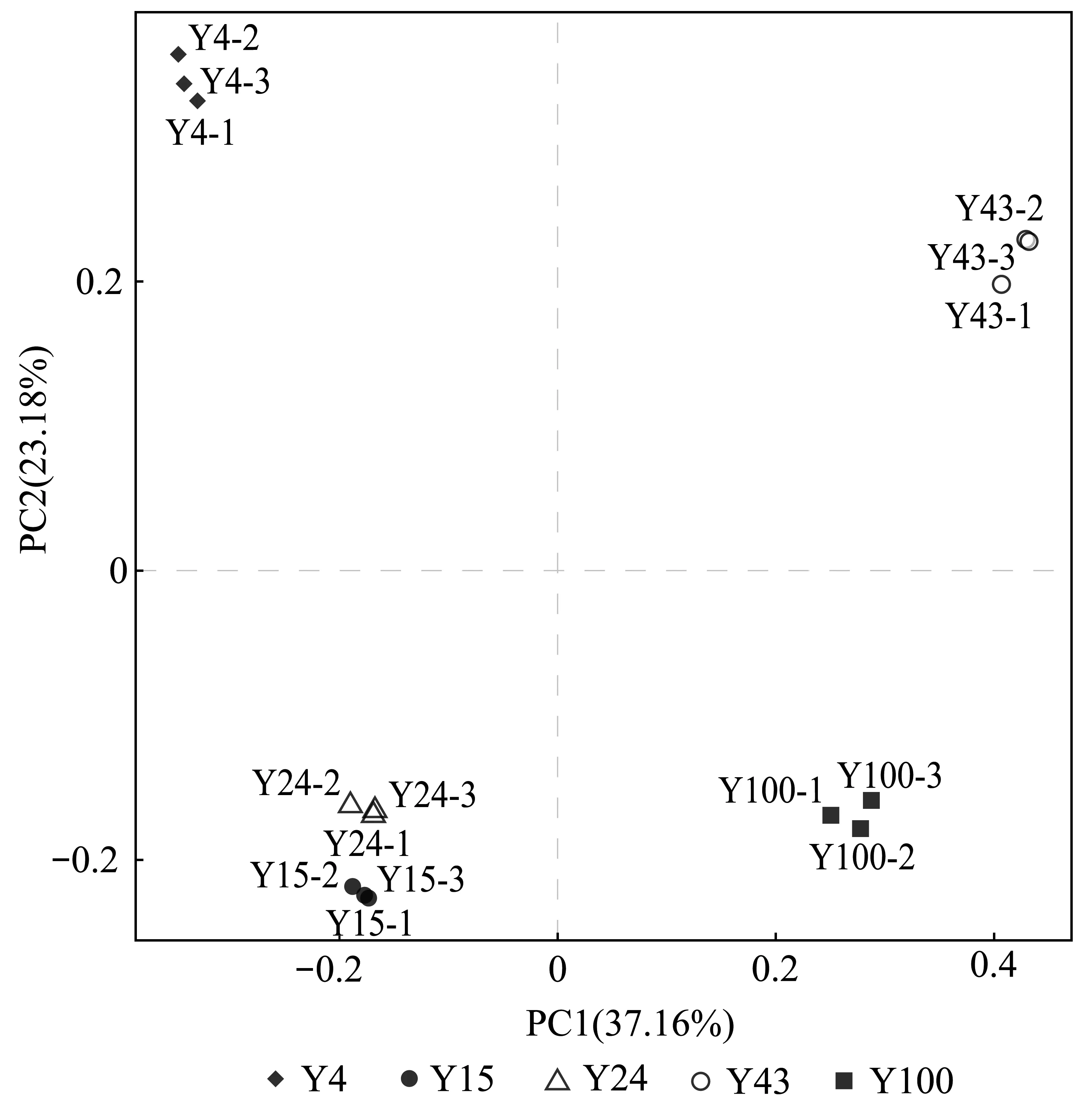
图3 不同林龄杉木林土壤解磷菌群落PCoA分析
2.3 不同林龄杉木林土壤中解磷菌多样性分析
由图4可知解磷菌Alpha多样性指数变化,土壤中解磷菌丰富度总体表现为43年>100年>24年>15年>4年,解磷菌种群多样性总体表现为43年>100年>15年>24年>4年。
用Hcluster分析的多样本聚类树的树枝结构可比较不同林龄土壤中解磷菌群落结构和多样性的异同。如图5所示,根据树枝长度大致可将样本分为4类: 4年,100年,43年,15和24年,即15年和24年林龄土壤中解磷菌组成相近,结构差异较小,4年林龄土壤中解磷菌独特性较高,与PCoA分析结果相同。

图4 不同林龄杉木林土壤解磷菌Alpha多样性指数

图5 不同林龄杉木林土壤中解磷菌基于bray curtis多样本聚类树
2.4 不同林龄杉木林土壤解磷菌含量特征
由图6可知,土壤解磷菌在门分类水平上,相对含量最高的类群为变形菌门(Proteobacteria),随林龄增加,变形菌门、慢生根瘤菌属(Bradyrhizobium)的含量整体呈下降趋势,表现为4年>24年>15年>43年>100年; 属分类水平,优势类群为慢生根瘤菌属、Pseudolabrys、假单胞菌属(Pseudomonas)、异常球菌属(Deinococcus)、甲基杆菌属(Methylobacterium)、Ramlibacter。慢生根瘤菌属整体呈下降趋势,100年相对含量最低,4年相对含量最高; 种分类水平上,慢生根瘤菌属的erythrophlei相对含量随林龄变化明显,24年和43年之间降低显著;Pseudolabrys_taiwanensis在43年处显著升高。随林龄增加,异常球菌属、Ramlibacter、极地单胞菌属(Polaromonas)变化趋势相同,均在15年林龄有最大值,4年林龄有最小值。阿菲波菌属(Afipia)变化趋势为4年>24年>43年>15年>100年; 放线菌门(Actinobacteria)变化趋势为43年>100年>15年>4年>24年; 厚壁菌门(Firmicutes)相对含量在100年林龄最高(0.45%); 假单胞菌属相对含量为4年>15年>24年>100年>43年; 甲基杆菌属43年林龄明显升高到1.75%。此外,本研究还发现100年林龄厚壁菌门的相对含量显著升高(0.45%),蓝细菌门(Cyanobacteria)和Gloeobacter在43年林龄含量明显高于其他林龄(1.08%),马赛菌属(Massilia)相对含量变化表现为 43年>15年>100年>24年>4年,在43年林龄最高,相对含量达到1.14%。
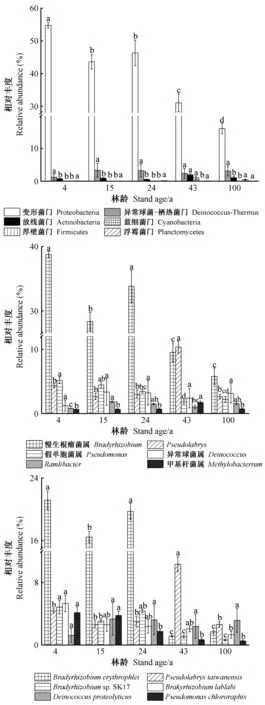
图6 不同分类水平上土壤解磷菌相对丰度
LefSe分析图由内至外辐射的圆圈代表了由门至属(或种)的分类级别。不同颜色节点分别代表在该组别中起重要作用的微生物类群,无显著差异的物种统一着色为黄色,差异物种跟随组进行着色。如图7所示,100年所对应的放线菌门、丙型变形菌纲(Gammaproteobacteria)、β-变形菌纲(Betaproteobacteria)、伯克霍尔德氏菌目(Burkholderiales)、丛毛单胞菌科(Comamonadaceae)呈红色,说明其在100年林龄杉木林中的相对丰度显著高于其他组; 同理,α-变形菌纲(Alphaproteobacteria)在4年杉木林中的相对丰度显著高于其他组; 蓝细菌门中的Gloeobacteraceae在43年林龄杉木林中相对丰度显著高于其他组。
2.5 不同形态磷素含量与解磷菌丰度的冗余分析
将土壤样品中的TIP、TOP、活性磷、中等活性磷和稳定态磷的含量与解磷菌的Alpha多样性指数进行相关分析(表2),可知解磷菌丰富度与多样性指数与活性磷含量呈正相关,Chao1和Observed_species指数(丰富度)与稳定态磷含量呈显著负相关(P<0.05)。
对土壤的不同磷素形态和解磷菌相对丰度进行冗余分析。图8a、8b、8c分别表示不同磷素形态与解磷菌门、属和种水平上优势群落的相关性大小。二者之间的相关性与夹角的余弦值有关,夹角越接近0°,表示二者呈显著正相关; 夹角越接近180°,代表二者呈显著负相关。土壤NaHCO3-IP与厚壁菌门相对含量呈显著正相关,H2O-IP与变形菌门相对含量呈显著负相关; NaOH-IP与放线菌门、芽单胞菌门(Gemmatimonadetes)呈显著负相关; HCl-OP与酸杆菌门(Acidobacteria)呈显著负相关(图8a)。 H2O-OP、NaHCO3-IP与慢生根瘤菌属呈显著负相关(图8b); NaHCO3-OP与阿菲波菌属、红游动菌属(Rhodoplanes)呈显著负相关。Bradyrhizobiumsp.SK17与Residual-P呈显著正相关,与H2O-OP呈显著负相关(图8c)。
3 讨论
3.1 不同林龄杉木林土壤磷素形态变化
Hedley磷素分级研究,为分析杉木人工林土壤中磷素供应及其生物有效性提供了一定参考。在本研究中,H2O-IP和NaHCO3-P含量逐年上升。H2O-P主要指与钾、钠结合的正磷酸盐(主要是HPO42-),可被植物直接吸收利用。NaHCO3-P包括吸附在土壤表面的无机磷和易于矿化的有机磷(孙桂芳等, 2011)。上述2种形态的磷素被认为是作物吸收的最有效磷源(春雪等, 2020)。据此分析可知,随着林龄增加,可供杉木生长的有效磷素水平不断提高。陈阳(2014)在对闽西北杉木人工林土壤理化性质的研究中发现,杉木纯林在0~20 cm土层中有效磷含量随林龄增加整体呈上升趋势。本研究发现NaOH-P含量随林龄变化波动明显,表现为24年>100年>43年>4年>15年。与王留芳等(2019)的研究相似,在湘中地区的植被恢复过程中土壤NaHCO3-P、NaOH-P快速累积。NaOH-P是土壤潜在磷源(Zhangetal., 2018),主要指通过化学吸附紧密结合在土壤铁铝化合物表面的有机磷和无机磷(余定坤, 2013),难以被植物直接吸收利用。土壤中的正磷酸盐阴离子(主要是H2PO4-)和高比表面积的铁铝氧化物之间强烈的吸附与解吸,对红壤中的磷素水平起着重要调控作用(Taghipouretal., 2013)。Andrea等(2011)研究表明,铁、铝可保护部分磷酸单酯免受土壤微生物降解。杉木林土壤中游离态铁铝氧化物含量最高,游离态铁铝氧化物与黏土矿物结合,提高了土壤团聚体的稳定性(赵友朋等, 2018)。与此同时,随着植被恢复年限增加,土壤非晶型、游离态和络合态铁铝氧化物含量逐渐增高(吴敏敏等, 2020),土壤潜在的供磷能力也将随之变化。据此分析,杉木人工林近熟林和过熟林土壤的潜在供磷能力高于幼龄林和中龄林,这可能是游离态铁铝氧化物含量随林龄增加逐渐升高,土壤团聚体稳定性增强所致。
本研究结果显示,随林龄增加,杉木林土壤中有机磷总量表现为100年>24年>43年>15年>4年。这一结论与曹娟等(2014)对湖南会同8、18、26年杉木人工林土壤中总有机磷含量比3年幼林含量高的研究结果相似。杉木近熟林的生物量最高,凋落物随成熟林林龄增加分解速率逐渐加快(郭琦等, 2014),在土壤表层有机质含量增加,促进了磷元素循环,成为有机磷的主要来源。本研究表明,杉木幼龄林、中龄林、近熟林中土壤磷素活性表现为稳定态磷>中等活性磷>活性磷,过熟林磷素活性表现为稳定态磷>活性磷>中等活性磷。这与林开淼等(2020)的研究结果相近,36年杉木林0~20 cm土壤中不同形态磷素含量表现为闭蓄态磷>中活性磷>易活性磷。在本研究中,稳定态磷包含HCl-P和Residual-P,此部分磷素在土壤中稳定存在,即闭蓄态磷。闭蓄态磷是土壤中由水化氧化铁胶膜包被的磷酸盐,其含量与土壤pH值、有机质、微生物等有关(孙桂芳等, 2011)。植物根系和部分土壤微生物通过产生有机酸和相关酶,提高土壤中难溶性含磷化合物的利用率(Tanetal., 2013)。此外,部分土壤微生物也可以通过形成菌丝,缩短磷扩散距离和增加吸收表面积,提高磷素吸收量(Shenoyetal., 2005)。稳定态磷含量与解磷菌丰富度呈显著负相关(表2)。因此土壤中稳定态磷的酸解与矿化,可能是导致土壤磷素活性变化的重要原因。
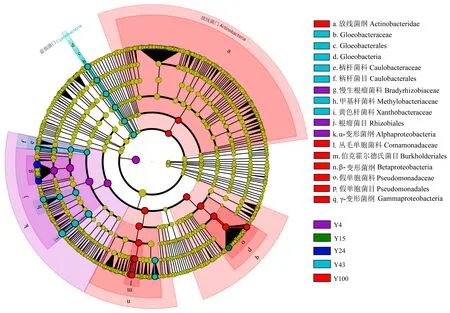
图7 土壤解磷菌群落LefSe分析
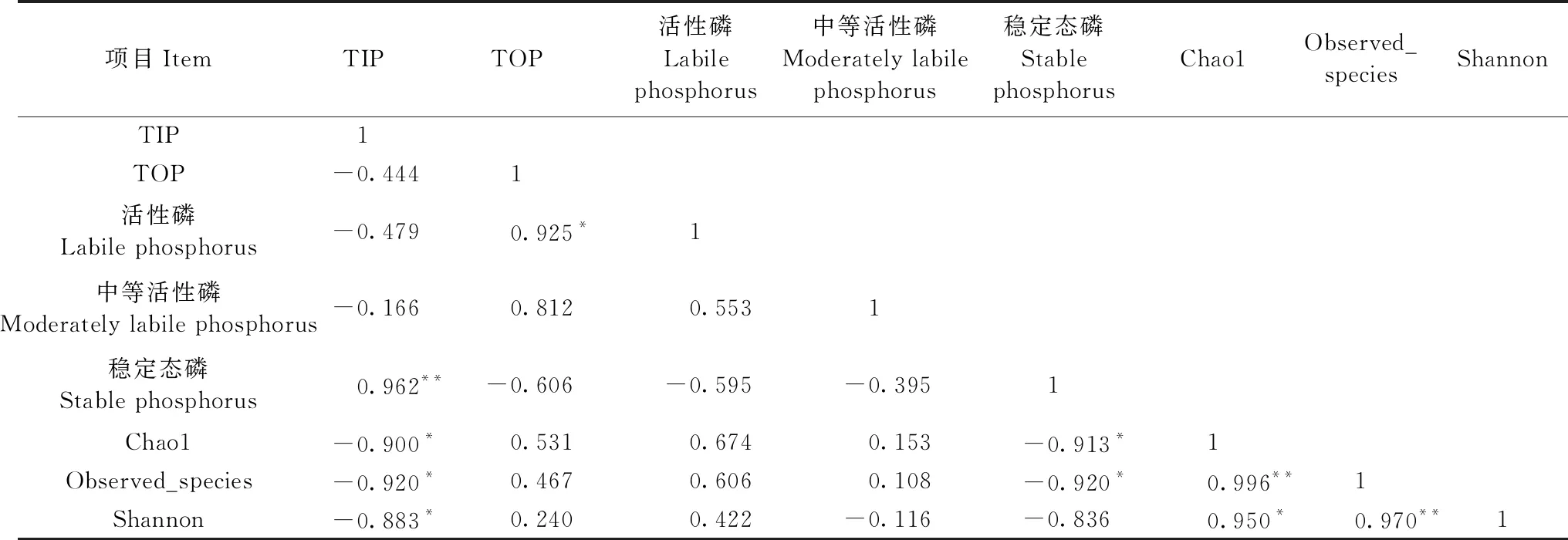
表2 磷素形态与解磷菌Alpha多样性指数的相关性分析①
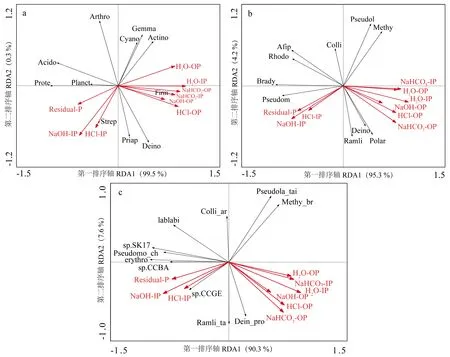
图8 不同磷素形态与土壤解磷菌相对丰度相关分析
3.2 不同林龄杉木林解磷菌群落结构与多样性变化
本研究结果表明,变形菌门作为门分类水平上的优势类群,随林龄增加其相对丰度整体呈现下降趋势,具体表现为4年>24年>15年>43年>100年; 放线菌门相对丰度变化为43年>100年>15年>4年>24年; LefSe分析结果显示,4年林龄杉木林中甲型变形菌纲的相对丰度显著高于其他组; 100年林龄杉木林中丙型变形菌纲相对丰度显著高于其他组。土壤铵态氮含量直接影响变形菌门的相对丰度,铵态氮含量越高,变形菌门相对丰度越大(李聪杰等, 2016)。根据本研究组之前对杉木林下土壤理化性质的分析,土壤铵态氮含量随林龄增长依次为11.05、11.00、9.66、8.64、8.39 mg·kg-1(曹升等, 2021),铵态氮含量变化与土壤变形菌门相对丰度变化一致。土壤总有机磷含量随林龄增加总体上逐渐升高(图1)。由于有机磷需经微生物矿化为无机磷才能被植物吸收,因此推测变形菌门中的某些类群在磷限制条件下对有机磷的矿化作用可能是林木对贫磷条件的一种适应机制。放线菌为K对策者,在资源有限的条件下,选择可维持长期生存状态的繁殖策略(Andreaetal., 2011)。甲型变形菌纲和丙型变形菌纲的是典型的r对策者,可在特殊环境下生存,一旦环境好转,繁殖速率会在短时间内迅速提升(Fuchsetal., 2000)。林媚媚(2009)对福建南靖和溪南南亚热带季风常绿阔叶林演替过程的研究发现,斑块内演替前期物种数目变化剧烈,经历急剧增长后,物种丰富度趋于平缓,各生长型和生活型组成比例趋于稳定,从演替初期逐渐向顶级斑块发展。区别于自然演替,人类活动对群落演替的影响不容忽视。在本研究区域,除幼龄期施加必要的抚育措施外,其他阶段人类活动对杉木林的影响较小。杉木林在不同生长阶段与生态学上的次生演替过程有一定程度相似。初期环境条件的变化对杉木生长影响较大,4年杉木林的林下植被较少,土壤微生物对资源、空间竞争等不激烈,适合甲型变形菌纲菌群生长; 随着林冠逐渐郁闭,林下植被正常生长受限,继而影响到微生物的生长繁殖。成熟林生态系统的自我调节机制使得林冠郁闭度下降,微生物丰度得到一定程度的恢复。在这个过程中,微生物种群也会根据环境条件的变化及时做出反馈调节。
杉木林土壤中解磷菌丰富度随林龄变化呈现出43年>100年>24年>15年>4年,Shannon指数变化与之类似,但15年杉木林土壤中解磷菌的多样性要高于24年。解磷微生物通过酶解和酸解作用,增加了土壤中有效磷形态含量,促进磷形态转化(春雪等, 2020)。如Wei等(2019)研究表明,土壤中解磷菌丰度和无机磷浓度呈正相关,与本文结果相印证。王蕊等(2020)研究发现,编码碱性磷酸酶细菌群落的Shannon指数与pH值呈极显著正相关,说明解磷菌群落的结构与功能受到土壤理化性质的影响。
整体来看,随杉木林龄增加,土壤中酸杆菌门含量逐渐降低,并且在100年林龄杉木林中相对含量近乎为0。一般认为,酸杆菌门的丰度与土壤pH值呈显著负相关(Jonesetal., 2009),且较多在贫营养型环境中生存较好。然而Tersia等(2020)研究显示,部分酸杆菌在pH值和有机质含量较高的环境中相对丰度增加。据本研究组之前对杉木林土壤理化性质的分析,土壤pH值随林龄增加波动明显,依次呈现出4.50、4.67、4.42、5.12和4.6(曹升等, 2021),酸杆菌门相对丰度随林龄变化趋势与土壤pH值变化大致呈负相关。据此分析,生存环境改变(pH值等)可能是引起酸杆菌门相对丰度变化的主要原因,但也有可能是不同菌类之间由于空间、资源等的竞争所致。在杉木生长过程中,稀有菌类(如Afipia、Massilia等)相对含量变化趋势明显,可能比优势菌类对环境的适应性更好,从而在不同形态磷素转化过程中发挥关键作用(Weietal., 2019)。
3.3 磷素形态变化与解磷菌的关系
杉木人工林土壤中磷素形态的转化与解磷菌有密切联系。根据不同林龄杉木林土壤解磷菌群落LefSe分析结果,可以看到100年林龄杉木林中伯克霍尔德氏菌目、丛毛单胞菌科相对丰度高于其他组。100年杉木林土壤中活性磷和中等活性磷的含量均处于较高水平,说明伯克霍尔德氏菌目的细菌的存在有助于土壤中磷素的释放,这一研究结果与骆韵涵等(2020)对罗源湾红树林根际土壤中解磷菌的研究结果相近,土壤pH值在一定程度上与解磷量呈负相关。伯克霍尔德菌属通过产生有机酸,特别是葡萄糖酸,使得磷酸盐被溶解。乔木层元素的累积量影响凋落物中元素含量。在杉木生长发育过程中,对P的需求量较N、K低,且随林龄增加,乔木层的P元素积累量逐渐升高(周玉泉等, 2019)。从幼龄林到中龄林,杉木林冠逐渐郁闭,土壤细菌、真菌数量减少,酶活性逐渐降低; 从中龄林到成熟林,林分密度及郁闭度下降,林下植被逐渐恢复,土壤细菌总数、磷酸酶等含量上升明显(焦如珍等, 1997)。杉木林下植被和凋落物的存在对土壤养分和微生物活性的维持具有重要意义。凋落物分解产生的有机酸(主要是草酸),使土壤pH值随林龄增加逐渐降低(邹显花等, 2017)。丛毛单胞菌属严格好氧,属化能有机营养型,对有机酸有良好的矿化作用,在中性或弱酸性环境下的降解能力较好(王巧蕊等, 2018)。这两类解磷菌可从一定程度上反映杉木林土壤磷素的转化机制和土壤的供磷能力。
4 结论
随杉木林林龄增加,H2O-IP和NaHCO3-P含量逐渐上升,NaOH-P含量波动明显,表明土壤活性磷含量不断提高,近熟林和过熟林土壤的潜在供磷能力高于幼龄林和中龄林; 总体来看,随杉木林林龄增加,解磷菌丰富度增大,从而对稳定态磷的矿化作用增强,导致稳定态磷向活性磷转化,因此土壤磷素有效性水平逐渐提高; 解磷菌丰富度经历急剧增长后,在43年时的物种丰富度和特有OTU数目均达到最大; 放线菌门相对含量与NaOH-IP呈显著负相关,表现为43年>100年>15年>4年>24年; 酸杆菌门相对丰度与NaOH-OP含量呈显著负相关。
