南亚热带常绿阔叶林4个常见树种的生物量分配特征与异速生长模型*
2022-05-11赵厚本周光益李兆佳邱治军吴仲民
赵厚本 周光益 李兆佳 邱治军 吴仲民 王 旭
(中国林业科学研究院热带林业研究所 南岭北江源森林生态系统国家定位观测研究站 广州 510520)
森林作为陆地生态系统的主体,其碳汇功能在调节全球碳平衡和应对全球气候变化方面发挥着重要作用。准确估算森林碳储量及其增长潜力是森林质量评价的重要内容,也是预测大气CO2浓度变化和全球气候变化的重要依据(Houghton, 2005; Yuetal., 2014)。森林生物量估算可用直接法(皆伐)和间接法(Brown, 2002)。直接法最精准但操作费时费力且具较大破坏性,不适用于大面积森林生物量估算,仅被少数学者采用(唐守正等, 2000)。采用遥感技术估测大面积森林生物量较为理想,但目前技术还不成熟(Xieetal., 2008)。生物量相对生长模型法通过采伐少量样本,将胸径、树高等简单易测的指标与生物量构建回归模型,有较好的估测精度,是迄今应用最广泛的估算方法(Hossainetal., 2016; Ubuyetal., 2018; 侯燕南等, 2016),也是校准新兴技术和新估算方法的基础(Juckeretal., 2017)。目前,全世界已构建了大量针对不对地区和树种的生物量模型(Chaveetal., 2005; Roxburghetal., 2016),也有研究构建适用于不同树种和森林类型的通用模型(Basukietal., 2009; Nafusetal., 2009)。但不同树种或同一树种不同地点的树木生物量分配格局、形态结构、木材密度等存在较大差异,导致其生物量相对生长模型也存在较大差异(Roxburghetal., 2016)。如采用通用模型估算特定森林的生物量,将会带来较大误差(Melsonetal., 2011)。为减小生物量估算误差和不确定性,需要构建针对特定森林类型和常见树种的专用生物量模型(Chaturvedietal., 2015; Hossainetal., 2016)。
我国亚热带常绿阔叶林具有很高的生物多样性,以壳斗科(Fagaceae)、樟科(Lauraceae)和山茶科(Theaceae)植物为主要优势物种,是亚热带大陆东岸季风气候区的典型植被类型(钟章成, 1988),覆盖了我国大概1/4的国土面积,其净初级生产力占全球森林的8%,在全球碳循环中占有重要地位(Yuetal., 2014)。在以往针对该地区森林生物量的研究中,由于没有成熟模型可用,一般采用平均标准木法,选择胸径树高接近所调查树木平均值的数株标准木测定生物量(曾曙才等, 2002; 林雯等, 2019; 谭家得等, 2012),这种方法用于人工同龄林精度尚可,但用于天然林或异龄林则存在较大误差。目前,己有学者在不同地区构建了针对亚热带常绿阔叶林不同树种的生物量模型(Linetal., 2017; Pengetal., 2017; 谢亭亭等, 2013; 左舒翟等, 2015),但多集中在地上生物量估算,地下生物量受限于根系取样困难,相关研究较少,不少学者采用地上和地下部分的比例估算地下生物量(Addo-Dansoetal., 2016),但树木地上地下生物量的比例在不同森林类型、树种、径级间存在较大差异,会导致估算结果的较大偏差(Xuetal., 2015)。针对南亚热带常绿阔叶林常见树种的生物量模型研究还不够完善。鉴于此,本研究选择南亚热带常绿阔叶林4个常见树种,实际测定生物量,分析其生物量的器官分配特征,构建各树种单株及各组分(地上部分和地下部分)生物量模型,以期提高生物量估算准确性。
1 研究区概况
研究区位于广东省广州市黄埔区福山村(113°30′17.9″—113°30′51.1″E,23°16′36.0″—23°17′11.8″N),距广州中心城区约26 km,为低山丘陵地貌,海拔128~153 m。广州市属南亚热带季风性气候,年平均气温21.7 ℃,年均降水量1 700 mm,降水集中在4—9月,约占全年降水量的80%,平均相对空气湿度为77%。地带性植被为南亚热带常绿阔叶林,试验点植被类型为砍伐后自然恢复的南亚热带常绿阔叶林,林龄约40年,土壤类型为红壤。
2 研究方法
2.1 样木选择
2016年,福山村一块40 hm2常绿阔叶林地块被规划为工业用地并办理了采伐手续, 本研究利用此契机,选择样木测定生物量。常见树种的判定以广州市2012年开展森林碳汇监测与计量研究项目设立的植物样地数据为依据,从共计312个植物样地中筛选出86块30 m×40 m的天然林样地,计算所有乔木的重要值并排序。对试验点进行踏查,选择数量和径级满足建立生物量模型条件且重要值排名前10的树种作为常见树种。最终选择了该地区常见的4个树种: 黧蒴(Castanopsisfissa)、中华锥(C.chinensis)、千年桐(Aleuritesmontana)和华润楠(Machiluschinensis),共79株样木,测定地上生物量。根据所选择样木位置情况,选择周围近距离内无其他树木且方便开展工作的64株样木,测定地下生物量。样木概况见表1。

表1 构建生物量相对生长方程的4个树种各胸径等级样本量
2.2 生物量测定
2016年9—10月进行样木生物量测定。地上部分生物量测定采用收获法: 用油锯在样木地上10 cm处伐倒,用皮尺测量准确树高,将树干从1.3 m处截断,然后每隔2 m截一段直至树冠。将树枝从树干上分离,将基部直径>3 cm的枝定义为大枝,在其直径3 cm处截开,截下部分为小枝。将所有树叶从树枝上分离。所有树干、大枝、小枝、叶的样品现场称鲜质量后取样,取样方法为: 每截树干锯1个厚度5 cm的圆盘;选择有代表性的3条大枝在基部和顶部各锯1个厚2 cm的圆盘;小枝用枝剪剪成小段取500 g样品;树叶取500 g样品。所有样品称鲜质量后装入布袋带回实验室。树干圆盘用浸水法测定体积用于计算木材密度ρ。所有样品65 ℃烘干到恒质量后计算含水率,根据含水率计算干质量。
树根生物量测定采用全挖法: 首先根据样木大小,以树桩为中心,用挖掘机挖一个直径1.5~3 m、深1.5~3 m的环形沟(挖出的土壤过2 cm孔径分样筛,以分离出根系),将树根连同土壤一起挖出,分离树根和土壤,所获根系分成4个部分: 根冠(树桩连同>5 cm粗根)、大根(2.5~5 cm)、中根(0.5~2.5 cm)和细根(<0.5 cm)。各部分现场称鲜质量,每部分采集500 g样品带回实验室,在65 ℃烘干到恒质量后称干质量,计算含水率并用于计算各部分干质量。
2.3 相对生长方程构建
幂函数(Y=aXb)或其对数变换形式(lnY=a+blnX)是最广泛使用的生物量相对生长模型(Linetal., 2017; Moussaetal., 2018)。由于生物量模型存在异方差,使用对数形式模型是消除异方差现象的方法之一。此外,在多元生物量模型中,自变量间可能高度相关从而使回归系数不稳定,使用对数形式模型进行线性回归可以通过方差膨胀系数(VIF)衡量多重共线性严重程度从而对自变量做出取舍。因此本研究采用对数形式的生物量模型。胸径(D)是推算生物量最重要的变量,树高(H)、木材密度(ρ)等其他变量通常作为第二变量或和D结合成单一变量应用在模型中(Carletal., 2017; Gouetal., 2017)。为评价H和ρ的加入对模型精度的影响,本研究选用以下5个生物量方程:
lnB=a+blnD;
(1)
lnB=a+bln(D2H);
(2)
lnB=a+blnD+clnH;
(3)
lnB=a+bln(D2ρ);
(4)
lnB=a+blnD+clnρ。
(5)
式中:B为器官或整株的生物量(kg);D为胸径(cm);H为树高(m);ρ为木材密度(g·cm-3),a、b和c为拟合系数。式(3)和(5)采用方差膨胀系数(VIF)衡量多重共线性程度(Dutcăetal., 2018)。采用决定系数(R2)和均方根误差(RMSE)进行模型效果评价和选优。
2.4 数据处理
一般认为,植物不同构件间的权衡关系为异速生长规律(陶冶等, 2014),因此用幂函数表示不同器官生物量占总生物量比例随胸径变化的关系。对各器官生物量占比的树种差异进行单因素方差分析(LSD法,α=0.05)。采用线性回归构建生物量模型。常规数据统计和作图在Excel 2013中完成,差异比较和回归分析在SPSS 21.0中完成。
3 结果与分析
3.1 生物量的各器官分配
不同器官生物量测定结果显示,4个树种均以叶生物量占总生物量比例最小,为5.34%~7.28%,华润楠叶生物量占总生物量比例显著低于黧蒴(P<0.05),其他树种间差异不显著; 树枝生物量占比为16.82%~24.20%,华润楠显著低于中华锥和黧蒴(P<0.05),其他树种间差异不显著; 树干生物量占比为47.22%~58.05%,中华锥显著低于其他3个树种(P<0.05); 树根生物量占比为14.25%~22.25%,黧蒴和千年桐显著低于中华锥和华润楠(P<0.05)(图1)。
随着胸径增大,黧蒴和中华锥的叶生物量占比呈上升趋势但不显著,千年桐和华润楠则呈极显著下降趋势(P<0.01); 各树种的树枝生物量占比均随胸径增大呈上升趋势,其中黧蒴和中华锥达到极显著水平(P<0.01),而千年桐达到显著水平(P<0.05); 各树种的树干生物量占比均随胸径增大呈下降趋势,其中黧蒴和中华锥达到了显著水平(P<0.05); 各树种的树根生物量占比和胸径均未表现出显著相关,表明树根生物量占比在树木生长过程中较稳定(图2)。

图1 各树种不同器官生物量占总生物量的比例
3.2 生物量相对生长模型
4个树种单株及各组分的生物量相对生长模型见表2。以D为单一自变量时,各树种单株及不同组分生物量模型均有较好拟合度,决定系数R2均值为0.955± 0.01,其中对单株和地上生物量拟合度较好,R2均值分别为0.972±0.007和0.971±0.009,对地下生物量拟合度较低,R2均值为0.920±0.025。以D为单一自变量时,单株及各组分生物量模型的拟合度以华润楠最高(R2=0.981±0.007),其次为黧蒴(R2=0.970±0.006)和千年桐(R2=0.947±0.027),中华锥最低(R2=0.921±0.029)。
当树高H以D2H形式和D整合成一个参数作为自变量时(公式2),所构建的地上部分生物量模型的拟合度较以D为单一自变量(公式1)的模型均下降,R2平均下降2.90%,RMSE平均上升32.54%; 对地下生物量拟合度虽有上升,但上升幅度较小,R2平均上升2.22%,RMSE平均下降15.67%; 对单株生物量则存在树种差异,对黧蒴和千年桐拟合度略有提高,对中华锥和华润楠则有所下降。总体来讲,若将H以D2H形式加入模型,弊大于利。
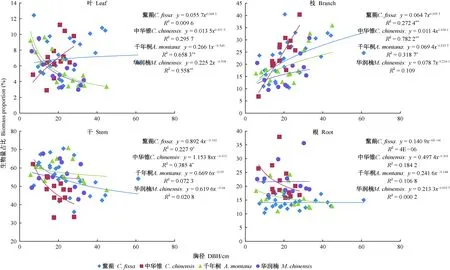
图2 各树种不同器官生物量占总生物量比例随胸径的变化趋势
当H作为独立第二自变量加入模型时(公式3),对4个树种地上生物量模型的拟合度影响较小,R2和RMSE的变化幅度都较小; 对地下生物量和单株生物量的拟合度均有所上升,R2分别平均上升1.75%和0.41%,RMSE分别平均下降11.10%和7.78%。总体来讲,将H作为单独的第二自变量加入模型,能提高模型精度但提升幅度有限。需指出的是,由于D和H存在极强的线性相关,模型的方差膨胀系数(VIF)较高(9.022±5.784),表明这2个系数存在较严重的多重共线性问题,这将大大降低模型在实际应用中的准确度。
当木材密度ρ以D2ρ形式和D整合成一个参数作为自变量时(公式4),所构建4个树种地上生物量和单株生物量模型的拟合度较D为单一自变量(公式1)模型的变化不大,R2变化幅度均低于1%; 对地下生物量拟合度的影响则存在树种差异,黧蒴拟合度略有提升,其他3个种有所下降(R2平均下降2.08%,RMSE平均上升18.14%)。总体上,将ρ以D2ρ形式加入模型将使模型精度略有下降。
当ρ作为独立第二自变量加入模型时(公式5),4个树种地上生物量和单株生物量的拟合度均有小幅提升,R2分别平均上升0.51%和0.18%,RMSE分别平均下降10.35%和3.54%; 对4个物种地下生物量拟合度均影响甚微。本研究中D和ρ没有显著相关,模型的VIF接近1(1.344±0.124),表明这2个系数不存在多重共线性问题。
4 讨论
如何有效分配生物量,对物种的生存和繁殖有重要意义(Weiner, 2004)。平衡生长假说认为,为了更有效地获取环境中的稀缺资源,植物会将资源分配到最需要的器官中,当某一构件获得收益时,另一构件就需付出适当代价(Mcconnaughayetal., 1999)。为了更好地适应生存环境,不同种类的植物有不同的生物量分配策略。一般情况下,耐荫树种倾向于将生物量更多地分配给枝条和叶片以增加树冠面积从而获取更多光照,而不耐荫树种则在茎部投入较多生物量以占据上层空间(Ntawuhiganayoetal., 2020)。本研究所选的4个树种基本符合这一规律,中华锥为耐荫树种,树冠生物量占比最大,表明该种倾向于产生大量枝叶以利于对横向空间的竞争; 千年桐为阳生树种,树干生物量占比最大,表明该种具有较高的树高,这有利于其占据林冠上层空间; 中华锥和华润楠的根生物量占比较高,表明这两个种拥有发达的根系,对土壤养分的竞争优势较强,对协迫的抗逆性也较好。
在生长过程中,植物的生物量分配策略会发生调整以适应不同阶段的竞争需求。随着胸径的增加,本研究所选4个树种的树干生物量分配比例呈下降趋势而树枝的生物量分配比例呈上升趋势,这表明这些种在幼树时分配较多生物量给树干以快速到达林冠层,达到一定高度后则分配更多生物量给树枝以获取更多光照。这种随个体生长调整生物量分配比例的策略在种间竞争极为激烈的南亚热带常绿阔叶林中是个有效的生存策略,但这种策略是否具有普遍性还需进一步研究,需获取更多树种数据的支持。在以往研究中,也有类似调整策略报道,如孟延山等(2019)报道青海云杉(Piceacrassifolia) 与祁连圆柏(Sabinaprzewalskii)的树干生物量占比随径级增加呈先下降后上升趋势。本研究所选4个树种的样木多集中在中幼龄阶段,未达到树木高度和冠幅稳定后树干持续生长阶段,因此未观测到树干生物量占比上升。
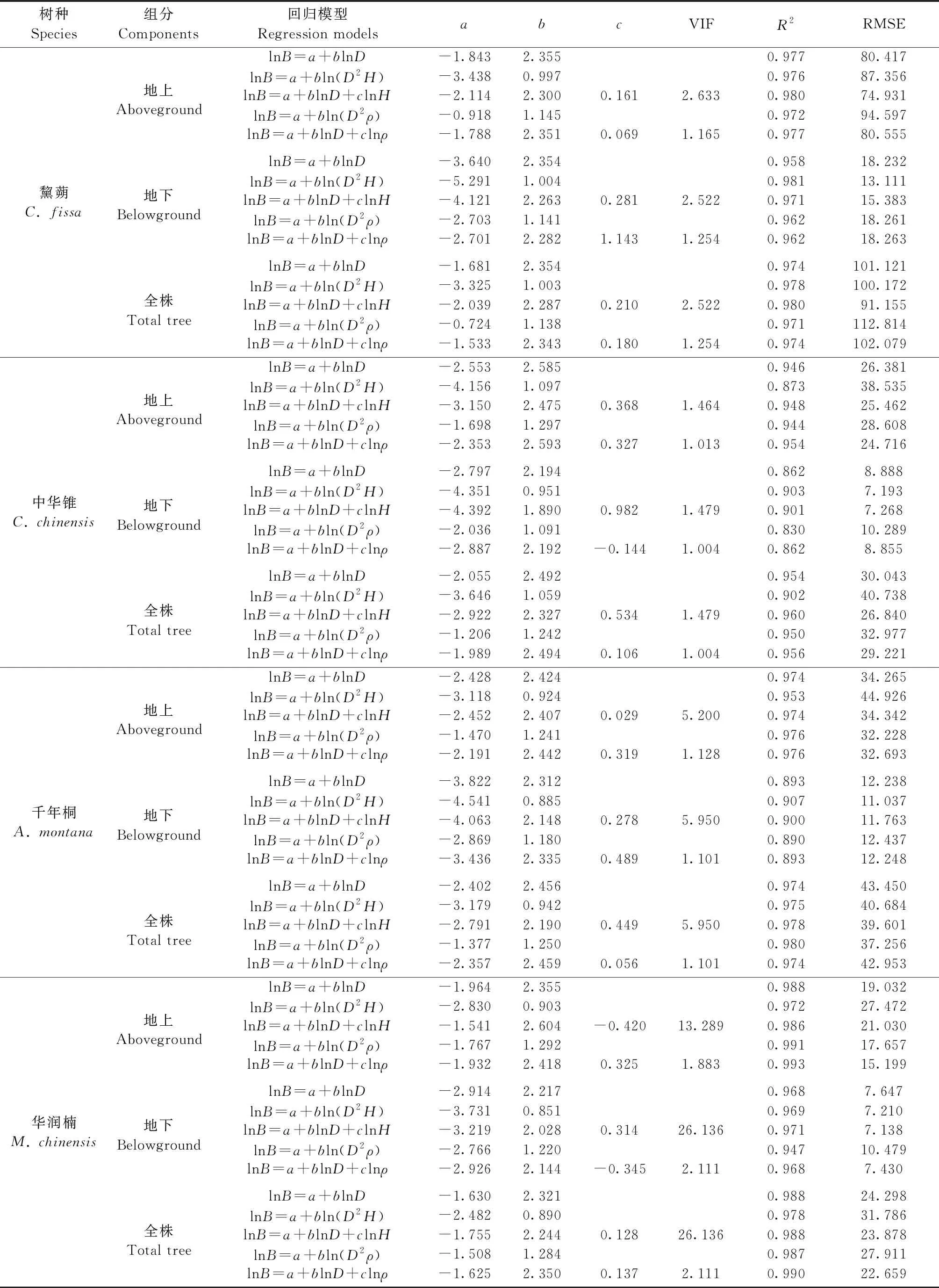
表2 不同树种各组分的生物量相对生长模型
胸径作为最常用和最容易被精确测量的指标,在生物量模型中应用最为广泛,且以D为单一变量的生物量模型往往可获得不错的精度(Linetal., 2017; 左舒翟等, 2015)。本研究以D为单一自变量的模型均有较高的R2和较低的RMSE,说明D和生物量存在极强的内在关联,是生物量模型的关键参数。
树高通常用来作为第二参数加入生物量模型以提高估算精度,但以往研究中H对提高模型精度的作用存在较大差异,既有H能提高模型精度(Mcnicoletal., 2015; Pengetal., 2017)的报道,又有能降低精度的报道,也有对精度无影响的报道(Gouetal., 2017; Xiangetal., 2016)。有研究表明,生物量模型中加入H能抵消区域效应(Chaveetal., 2005; Feldpauschetal., 2012),但Dutcă等(2018)的研究结果表明,H仅能减少树干生物量模型的区域效应却增加了地上和全株生物量的区域效应。本研究中将H以D2H的形式包含在模型中降低了地上和全株生物量的估算精度,对地下生物量的估算精度虽有提高但提高幅度很有限,这主要是因为H和生物量间的内在关联不够紧密,树木往往在早期生长阶段快速长高到达林冠层,之后再扩展横向空间,使得异龄林中不同径级树木往往具有相近的高度。将H作为第二自变量包含在模型中虽能提升估算精度,但增加了模型的多重共线性问题。共线性问题会加大模型预测参数的置信区间使模型不确定性增加,此外,回归数据中小的变化就会带来预测参数较大的改变从而影响模型精度(Dutcăetal., 2018)。在野外调查中,准确测量H也存在较大难度,目测法使用最多但误差也最大,激光测距仪、测高仪等设备虽然测量准确,但在常绿阔叶林中树冠的相互遮挡使寻找树木最高点极为困难。因此,不建议在估算生物量时将H包含在模型内。
木材密度是估算生物量的重要参数,特别是利用通用模型估算多树种组成的森林生物量时具有重要作用(Chaveetal., 2014),实际应用中也有较多ρ提高模型精度的报道(Pengetal., 2017; Xuetal., 2015),但也有将ρ加入模型不能提高精度的报道(Linetal., 2017)。本研究中,将ρ以D2ρ的形式包含在模型中对各部分生物量的估算精度均有所下降,但将ρ作为第二自变量包含在模型中则能提高估算精度,且D和ρ间不存在显著相关关系,模型也不存在多重共线性问题。因此,在条件允许时,使用D和ρ两个自变量构建生物量模型能提高预测精度。然而实际调查中较难准确获得ρ数据,如将ρ引入模型将大大增加调查难度从而降低模型实用性; 而且增加ρ为第二参数后,模型的解释能力提高幅度也不大。因此应用中需根据实际情况和调查目的,综合考虑准确性和工作难度间的平衡,选择合适的模型。
生物量异速生长模型具有高度的物种和区域特异性,构建特定地区特定物种的生物量模型对森林生态评价和区域碳汇计量具有重要意义。本研究所选4个树种是根据广州地区森林样地调查结果统计出的优势度较高的树种,有一定代表性。建模样本取自广州市次生常绿阔叶林,为华南地区最有代表性的林型。模型适用范围为南亚热带相似水热条件次生林,对其他相近区域内这4个树种生物量的估算也有参考价值。
采用收获法研究树木生物量模型存在环境破坏大、取样困难、工作量大等缺点。本研究存在样本量较少、研究结果较为粗糙的问题,同样的问题也存在于其他类似研究中。遥感技术的发展使得对树木进行3D重建成为可能,配合地面实测进行校正将大大提高遥感对生物量估测的精度,引发该研究领域的变革。受采样区域林龄限制,本研究样木径级的涵盖范围不够广,模型对大径级树木生物量的估算可能存在较大偏差,在应用时要慎重选择。
5 结论
本研究所选的南亚热带常绿阔叶林4个常见树种中,阳生树种千年桐分配较多生物量给树干,耐荫树种中华锥分配较多生物量给枝叶,耐荫树种中华锥和华润楠的根系更为发达。随着胸径的增加,4个树种的树干生物量分配比例呈下降趋势而树枝的生物量分配比例呈上升趋势。
以D为单一自变量的生物量模型对4个树种各部分生物量的估算均有较好精度。将H作为自变量加入模型会降低模型精度或增加多重共线性问题,加之野外较难准确测得树高,因此不建议将H包含在模型中。将ρ作为独立第二自变量包含在模型中,能提高模型估算精度但升幅不大,建议实际应用中根据调查目的在准确性和工作难度间做出取舍。
