不同光照处理对赤苍藤光合生理和生化特性的影响
2022-05-09覃凌薇王凌晖
卢 庸,覃凌薇,李 琳,3,王凌晖
(1.南宁树木园,广西南宁 530000;2.广西大学 林学院,广西南宁 530004;3.玉林师范学院,广西玉林 537000)
赤苍藤(Erythropalum scandens)为铁青树科(Olacaceae)赤苍藤属多年生常绿藤本植物,别名菜藤、侧苋或牛耳藤等[1-3]。赤苍藤主要种植于我国云南海拔1 000 ~1 500 m 及广西和广东海拔280 ~550 m 的地区,多见于低山丘陵、山区溪边、山谷、疏林、林缘或灌丛中[4]。赤苍藤被称为零污染、无公害及富有营养的天然绿色药食同源产品,内含丰富的蛋白质、胡萝卜素、维生素B、维生素C 及对人体有益的矿物质和铁、锌、铜及锰等微量元素,硫胺素和核黄素的含量远高于一般家用蔬菜[5]。赤苍藤具有清热祛湿、祛风活血的功效,在广西民间常用于肝炎、肿瘤、尿道炎和急性肾炎等疾病的治疗[6]。潘乔丹等[7]研究表明,赤苍藤能够抑制黄腺嘌呤氧化酶(XOD)的活性,降低高尿酸血症血清中的尿酸含量,起到保护血管的作用;许崇摇等[8]研究表明,赤苍藤茎叶中的提取物(ASLE),具有抗痛风的功效。赤苍藤还具有园林观赏价值,花冠白色,种子蓝绿色,果实成熟时下垂,形似灯笼。赤苍藤是一种价值丰富、潜力巨大的野生蔬菜及园林藤本。
光是植物生长发育的重要条件。当光环境发生变化时,植物会产生一系列生理和生化响应过程,如光合色素合成过程加快或减慢、渗透物质变化及活性氧增加或减少[9]。许多学者已对不同光环境对植物的生理响应进行探究[10-14]。目前对赤苍藤的研究主要集中在药物作用[7-8]、施肥[15]和人工栽培[16-17]等方面,不同光环境对赤苍藤的影响尚未见报道。本研究通过比较不同光照条件下赤苍藤幼苗的光合生理和生化指标,利用主成分分析法,探讨其最适合的种植条件,以期为赤苍藤的人工栽植及耐荫性研究提供参考。
1 材料与方法
1.1 试验地概况
试验地位于广西南宁树木园新塘管理区林地(108°22′E,22°46′N),属典型亚热带季风气候,年均气温22 ℃,夏秋季雨水充沛,年均降水量1 650 mm,无霜期长,适合赤苍藤幼苗的生长。
1.2 试验材料
赤苍藤幼苗由南宁树木园珍稀树种花卉苗木繁育中心提供,为2 ~3年生实生苗。选取90 株长势一致、无病虫害赤苍藤实生苗进行试验。
1.3 试验方法
参照陈军[18]完全随机试验设计,将赤苍藤幼苗随机分为3 组。设置3 个光照处理:自然光照(100%NS)、遮光率为35%的光照强度(65%NS)及遮光率为70% 的光照强度(30%NS)。采用LI-6400XT 便携式光合作用测量仪测定每个遮荫处理的透光率。每个光照设置3 个重复,每重复10 株。2019年5月,开始进行遮荫处理,试验期间保持水肥管理一致。
1.4 指标测定
2019年11月,采摘成熟功能叶。每个处理选取长势大体一致、叶片完好且无病虫害的植株3 ~4株,采集中上部功能叶3 ~4 片,每个处理重复3 次。采集后将叶子放入装有冰袋的盒子中,带回实验室进行各项光合生理和生化指标的测定。参考《植物生理学实验指导》[19]测定各项光合生理和生化指标。叶绿素采用95%乙醇研磨、提取及测定,采用蒽酮比色法测定可溶性糖含量,采用考马斯亮蓝法测定叶片可溶性蛋白含量,采用硫代巴比妥酸比色法(TBA)测定丙二醛(MDA)含量,采用茚三酮法测定游离脯氨酸含量,采用LI-6400XT 便携式光合作用测量仪测定植株的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间二氧化碳浓度(Ci)。
1.5 数据处理
采用Excel 2019 软件进行数据处理与图表制作;采用SPSS软件进行数据分析。
2 结果与分析
2.1 不同光照处理对赤苍藤叶片光合色素的影响
65%NS 处理的叶绿素a、叶绿素b、类胡萝卜素和叶绿素a + b 含量均极显著高于100%NS 和30%NS 处理(P<0.01),叶绿素a/b 极显著低于100%NS 和30%NS 处理(P<0.01);100%NS 和30%NS 处理的叶绿素a、叶绿素b 和叶绿素a + b 含量均差异显著(P<0.05),叶绿素a/b 和类胡萝卜素含量差异不显著(表1)。与100%NS 和30%NS 处理相比,65%NS 处理的叶绿素a + b 含量分别高出70.62%和37.14%,类胡萝卜素含量分别高出29.03%和16.50%。表明在适度遮光处理下,叶绿素a、叶绿素b、类胡萝卜素和叶绿素a+b 含量均有一定程度的增加。
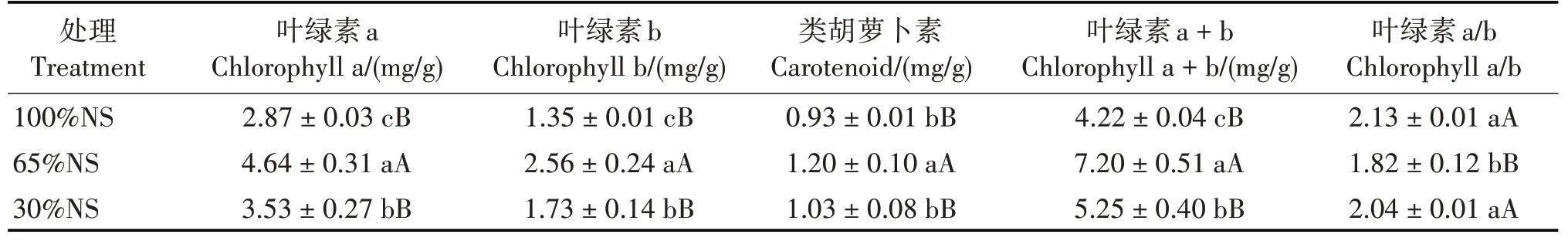
表1 不同光照处理下赤苍藤实生苗的光合色素含量Tab.1 Photosynthetic pigment contents of E.scandens seedlings in different light treatments
2.2 不同光照处理对赤苍藤叶片光合特性的影响
随光照强度减弱,赤苍藤幼苗的Pn呈先降后升的趋 势;30%NS 和100%NS 处 理的Pn极显著高于65%NS 处理(P<0.01),分别高出71.43% 和33.96%;30%NS 和100%NS 处理间差异不显著(表2)。随光照强度减弱,Gs逐渐降低;100%NS 处理的Gs极显著高于30%NS 和65%NS 处理(P<0.01),均高出66.67%;30%NS 和65%NS 处理间差异不显著。随光照强度减弱,Ci逐渐降低;100%NS 和65%NS 处理的Ci极显著高于30%NS 处理(P<0.01),分别高出44.47%和24.69%;100%NS和65%NS处理间差异显著(P<0.05)。随光照强度减弱,Tr呈先升后降的趋势;3 个处理间均差异极显著(P<0.01),30%NS处理的蒸腾速率最低。

表2 不同光照处理下赤苍藤实生苗的光合特性Tab.2 Photosynthetic characteristics of E.scandens seedlings in different light treatments
2.3 不同光照处理对赤苍藤叶片渗透调节物质含量的影响
随光照强度减弱,赤苍藤叶片中的可溶性糖含量呈不断增加的趋势,3 种光照处理的可溶性糖含量差异均不显著(图1a)。随光照强度减弱,可溶性蛋白含量呈先降后升的趋势,100%NS 和30%NS 处理均极显著高于65%NS 处理(P<0.01),100%NS 和30%NS 处理间差异不显著(图1b)。随光照强度减弱,游离脯氨酸含量呈先升后降的趋势,65%NS 和30%NS 处理均极显著高于100%NS 处理(P<0.01),65%NS 和30%NS 处理间差异不显著(图1c)。随光照强度减弱,MDA 含量呈先升后降的趋势,100%NS和30%NS 处理极显著低于65%NS 处理(P<0.01),100%NS和30%NS处理间差异不显著(图1d)。
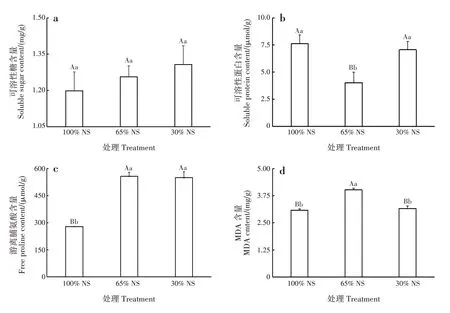
图1 不同光照处理对赤苍藤实生苗渗透调节物质含量的影响Fig.1 Effects of different light treatments on osmotic adjustment substance contents of E.scandens seedlings
2.4 主成分分析
对赤苍藤在3 种不同光照处理下的13 个光合生理和生化指标进行主成分分析,两个主成分贡献率分别为70.215% 和29.785%,累积贡献率为100%,表明这些指标能最大程度地保留原始性状的所有信息(表3)。第1 主成分主要包括叶绿素a、叶绿素b 和类胡萝卜素等指标,特征值为9.128,贡献率为70.215%;第2 主成分主要包括Pn、可溶性糖和游离脯氨酸等指标,特征值为3.872,贡献率为29.785%。根据各综合指标的标准化值和标准化特征向量,计算两个主成分的得分及不同光照处理的综合得分。线性组合方程分别为:
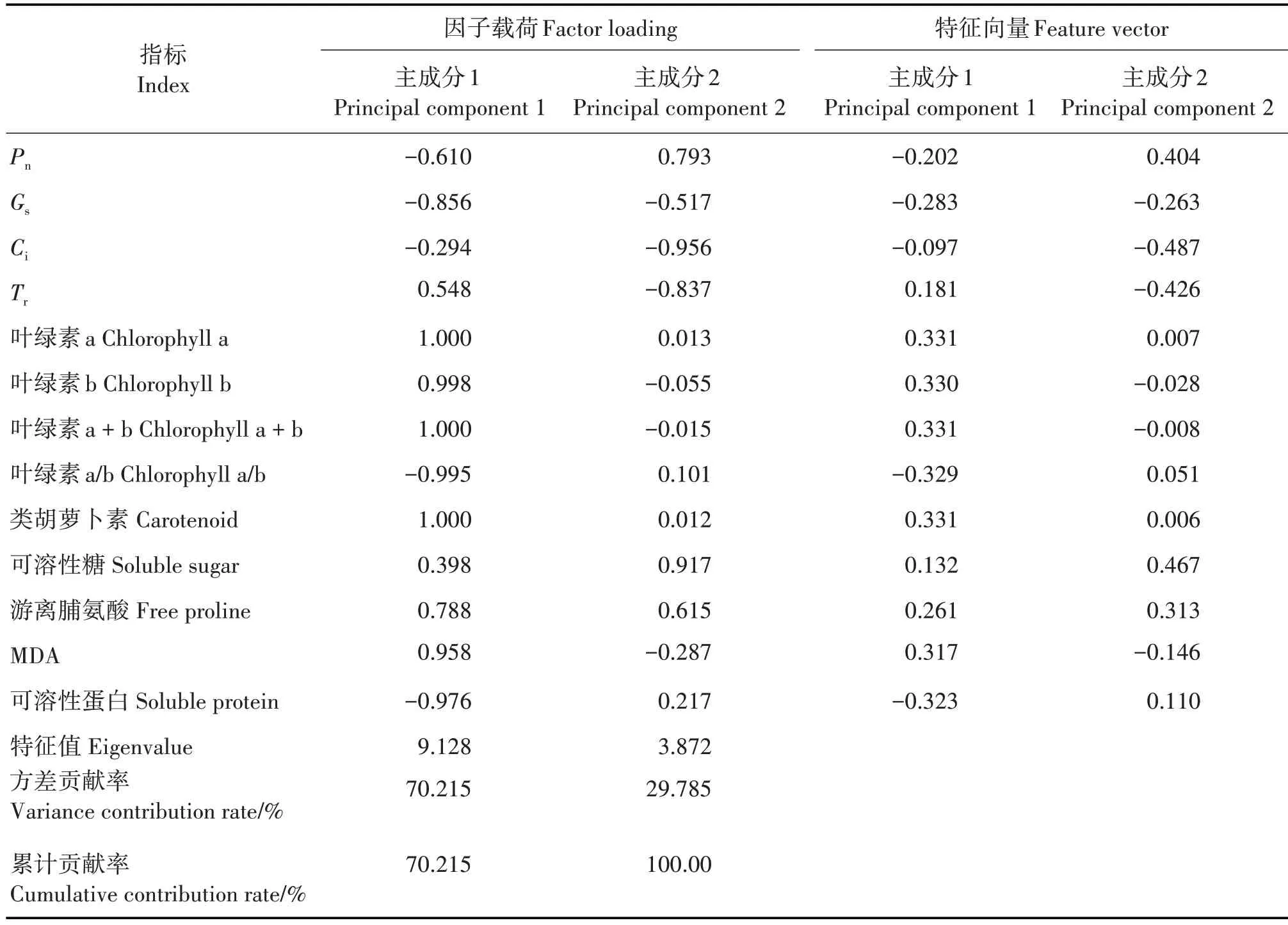
表3 不同光照处理下赤苍藤实生苗光合生理和生化指标的主成分分析Tab.3 Principal component analysis on photosynthetic physiological and biochemical indexes of E.scandens seedlings in differient light treatments
Y1= -0.202X1- 0.283X2- 0.097X3+ 0.181X4+0.331X5+ 0.33X6+ 0.331X7- 0.329X8+ 0.331X9+0.132X10+0.261X11+0.317X12-0.323X13
Y2= 0.404X1- 0.263X2- 0.487X3- 0.426X4+0.007X5- 0.028X6- 0.008X7+ 0.051X8+ 0.006X9+0.467X10+0.313X11-0.146X12+0.11X13
式中,X1为Pn;X2为Gs;X3为Ci;X4为Tr;X5为叶绿素a;X6为叶绿素b;X7为叶绿素a+b;X8为叶绿素a/b;X9为类胡萝卜素;X10为可溶性糖;X11为游离脯氨酸;X12为丙二醛和X13为可溶性蛋白。
按照以上线性组合方程计算,综合得分排名由大到小依次为65%NS、30%NS和100%NS(表4)。
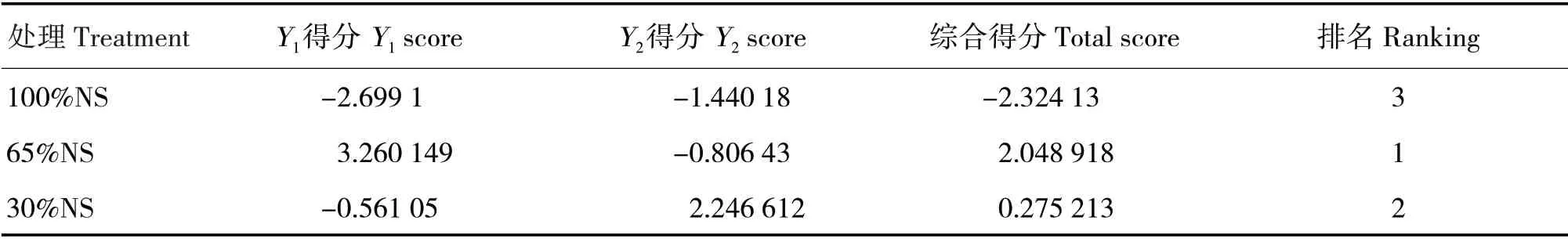
表4 不同光照处理主成分得分和综合得分Tab.4 Principal component scores and comprehensive scores in different light treatments
3 讨论与结论
叶片是合成光合色素的主要器官,是光合作用的重要器官。光照对总叶绿素含量的影响主要有两种观点。一种是随光强的减弱,植物叶片的总叶绿素含量上升而叶绿素a/b 下降[20];另一种是叶绿素含量和叶绿素a/b 均随光照强度减弱而降低[21]。本研究结果表明,赤苍藤幼苗在65%NS 条件下,叶片中的叶绿素a、叶绿素b、叶绿素总量和类胡萝卜素含量均增加,但叶绿素a/b 比值降低。随光照强度减弱,叶绿素含量呈先升后降的趋势,各处理间差异显著。与全光照相比,遮光条件下,赤苍藤幼苗可通过提高叶片叶绿素a、叶绿素b和类胡萝卜素含量,提高植株的捕光能力,由此适应弱光环境。这与熊静等[22]对朱砂根(Ardisia crenata)幼苗、陈艺群等[23]对茄子(Solanum melongena)幼苗和杨亚男等[14]对四季桂(Osmanthus fragrans)的研究结论一致,遮光条件下,植物光合色素呈增加趋势。有研究表明,光照逆境下,限制植物光合作用主要有两个因素,分别为气孔因素和非气孔因素,当Ci下降,Pn下降,气孔因素为光合速率降低的主要因素;Ci上升,而Pn下降,非气孔因素为光合速率降低的主导因素[24]。本研究结果显示,随光照强度减弱,赤苍藤幼苗Pn呈先降后升的趋势;与100%NS 相比,65%NS 处理的叶片Ci降低,此时降低Pn也降低,主要原因为气孔因素;与100%NS相比较,30%NS处理的叶片Ci降低,但此时Pn增加,主要原因为非气孔因素。
渗透物质对维持细胞内渗透压、防止水分丧失、增强膜蛋白分子间水合作用具有重要意义[25]。可溶性糖、可溶性蛋白、游离脯氨酸和MDA 都是植物细胞内重要的渗透调节物质。逆境胁迫下,可溶性糖可作为渗透保护剂,保持活性氧平衡,缓解植物逆境伤害[26]。本研究中,可溶性糖含量随光照强度减弱逐渐上升,但处理间差异不显著,这与陈敬忠 等[27]对罗甸小米核桃(Juglans regiaf.luodianense)幼苗生理响应的研究结论一致。可能原因是可溶性糖在赤苍藤幼苗光照胁迫调节中并未发挥重要作用。随光照强度减弱,可溶性蛋白含量呈先下降后上升的趋势,这与郭品湘等[28]对双色木番茄(Solanum wrightii)幼苗随光照强度减弱,可溶性蛋白含量先降后升的研究结论一致,与翟玫瑰等[29]对茶花(Camellia japonica)幼苗随光照强度减弱,可溶性蛋白含量下降研究结论不一致。具体原因有待有待更进一步探究,有可能是植物之间的差异[30],也有可能与本研究中设置的光照强度梯度过大有关。当植物处于光照胁迫下,无论强光还是弱光,都会导致叶片中膜脂过氧化加剧,MDA 含量增加[31]。在本研究中,随光照强度增加,叶片中可溶性蛋白含量呈先上升后下降的趋势,MDA 含量呈先下降后上升的趋势,表明在一定光强范围内,提高光照强度,有利于延缓植物叶片的衰老。本研究中,随光照强度减弱,游离脯氨酸含量呈上升的趋势,可能是过度遮荫对赤苍藤造成了胁迫危害。还需进一步研究赤苍藤叶片中抗氧化物酶等生理生化指标,以便进一步确定植物的耐荫机制。
根据主成分分析可知最佳处理为65%NS。本研究的试验材料为2 ~3年生实生苗,可能与多年生赤苍藤的研究结果有些差异,同时人工种植生长环境与自然生长环境有所差异,研究结果可能会受到一定影响。
