基于群体行为的根系构型仿真
2022-05-09李松阳高继勋王淼
李松阳 高继勋 王淼




摘要:为了分析根系群体行为机制对环境刺激下根系生长发育的影响,提出了一种基于群体行为的根系构型可塑性分析模型。把整个根系视为由智能个体单根组成的智能群体,利用智能个体单根间的排斥、对齐和偏好构建环境感知的根系群体行为模型,结合通用根系模型ArchiSimple构建群体行为影响的根系生长发育模型。设置随机、分层、梯度3种不同的环境条件,比较分析了群体行为、随机生长和向性生长机制对根系长度、体积、分布和资源吸收量等特征影响。显著性检验结果表明,群体行为、随机生长、向性生长机制有显著不同;在分层和梯度环境条件下,3种生长机制对资源吸收量的影响有显著差异;在其他情况下3种生长机制对根长、体积、分布等特征的影响无显著差异。进一步分析表明,群体行为机制能够提升根系资源利用效率,群体行为模型能够被用于环境影响的根系构型可塑性分析。
关键词:群体行为;根生长策略;土壤环境;功能结构根模型;根可塑性
中图分类号:TP391.9 文献标志码: A
文章编号:1002-1302(2022)07-0196-05
收稿日期:2021-06-23
基金项目:国家自然科学基金(编号:61501174);国家自然科学基金-河南联合基金(编号:U1704124);河省高等学校重点科研项目(编号:21A520005);河南工程学院科研培育基金(编号:PYXM202020)。
作者简介:李松阳(1985—),男,河南南阳人,博士,副教授,主要从事图像处理、植物建模仿真研究。E-mail:lsy_cqu@cqu.edu.cn。
根系结构是植物生存策略中非常重要的组成部分,表达了根系对土壤中可用养分的适应性。根系会形成复杂结构来优化利用土壤养分。在根系与土壤的交互中,根系可以通过其结构变化来适应环境[1]。
研究表明,根系在定向环境刺激下展现出可塑性定向生长。例如,根系能够依据水分或营养的分布梯度朝向资源丰富的区域生长。根系这种特性(向性)能够寻找充足的水分和其他营养物质[2]。除了向性之外,根系采用不同的生长和分枝策略来影响根系结构。例如,当根系处于营养丰富的区域时,会增加根系的吸收面积。在营养充足的条件下,根系分枝密度会提高。但当营养缺乏时,根系分枝的出现会受到抑制[3]。除了上述策略外,侧根与其父根(产生侧根的根)的生长率差异也是根系适应营养可用性的一种策略。比如在营养缺乏区,根系的侧根薄而短,而在营养丰富区,根系会形成更厚更长的分枝[4]。可见,当根系与周围土壤环境交互时,向性、分枝率以及生长率等可塑性对根系构型具有重要的影响,从而形成复杂的根系。
由于根系非常复杂,难于直接观察,根系模型被提出用于分析可塑性对根系构型的影响。其中,ROOTMAP模型最早被提出,用于模拟须根系的生长[5]。随后,RootTyp、SimRoot、ArchiSimple、SPACSYS、R-SWMS、RootBox等根系模型被提出,用于模拟根系、根系构型可塑性及其与土壤的相互作用。最近,CRootBox、OpenSimRoot根系模型被提出,用于模拟根系构型及其与静态、动态土壤环境的相互作用[6-7]。这些根系模型都能根据根系的生长发育过程再现接近自然环境的根系结构。但是这些模型都是基于数学方法,依据统计结果,来生成根系。即使这些模型完美地再现了根系构型,但不能清楚地解释根系生长发育的决策过程。
最近研究表明,根系中存在群体行为。Ciszak等提出基于根系群体行为理论模型,揭示了根系生长会受到“邻居”的影响,导致根生长方向趋于一致[8]。Matos等研究了根尖集体决策在要探索土壤营养中的作用[9]。McCleery等将根系视为一群耦合的多细胞结构,并提出根系觅食可视为这些结构的集体行为[10]。因此群体行为对根系适应环境变化起着重要作用。
虽然已有许多根系模型被提出,但很少有模型考虑到群体行为对根系构型的影响。本研究提出一种根系群体行为模型,并与通用根系模型ArchiSimple集成,比较分析了在随机、分层、梯度3种不同的虚拟土壤条件下,群体行为(swarming)、随机生长(random)和向性(chemotropism)3种机制对根系结构的影响。
1 模型与方法
1.1 根系生长模型
ArchiSimple是一个简单通用的根系模型。该模型中根系表示为单根集合,已经在香蕉、豌豆、桃树、玉米等物种进行了校准和评估[11]。因此,ArchiSimple模型被用作根系生长发育的基础模型。假设单根( sr)为三元组,包含根尖(称为rt)、根段(rs)和根原基(rp),如式(1)所示:
sr=(rt1,rs+,rp+)。(1)
式中:rt1表示1个根尖(rt);rs+表示设有多个根段(rs);rp+表示设有多个根原基(rp )。
根系可以描述为一组单根(sr)构成。单根伸长和分枝是影响根系生长发育的主要因素。在ArchiSimple模型中单根伸长过程见式(2)。
pgr=0,d≤d min
E×(d-d min), d>d min。(2)
式中: E是潜在伸长率影响因子;d min是最小根直径,当根直徑小于 d min时,根不再伸长; d max表示最大根直径; d为当前根直径。可见,单根潜在伸长长度(pel)是潜在伸长率(pgr )和时间间隔(Δ t )的乘积。
在ArchiSimple模型中单根分枝过程为随着单根的伸长,会产生新的根原基,根原基出现并发育为侧根。假设根原基沿单根方向按照根原基间距离( ipd)序列初始。根原基在一定的时间延迟后,依据潜在侧根出现概率(plre )形成一个新的单根。该单根的初始直径如式(3)所示:
d=d m×d bm0×eφ×R。(3)
式中:d m表示父根根段直径;d bm0是相对于其父根最大分枝直径;φ为调节方差常数;R 是0和-1之间均匀分布的随机数。
在根系生长过程中,须要从土壤中吸收养分。当需求和吸收不平衡时,根系生长受到胁迫,将根系生长压力( rgs)定义为式(4)。
rgs(u)=0 u min≥ u
u-u min u opt -u min 其他
1u opt ≤u。(4)
式中:u是每个时间间隔内的局部资源吸收量;u min是最小资源需求; u opt是最优的资源需求。根系实际生长速率和实际侧根出现概率受到根系生长压力的影响。
1.2 群体行为模型
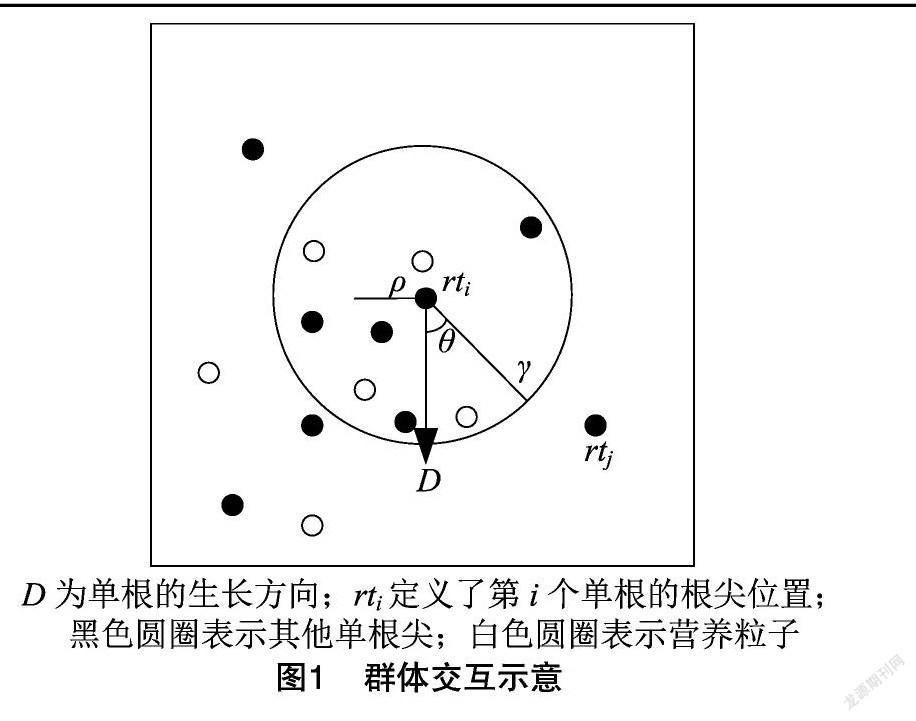
研究表明,考虑排斥、对齐、偏好3种作用,能够产生与真实群体非常相似的行为模式[12]。在根系群体行为模型中,把单根看作一个移动的个体,将单根的长度视为个体经过的路径。假设每个单根与其相邻的单根通过排斥区或对齐区,感知资源,通过对齐、排斥和偏好相互交互,建模根系群体行为(图1)。
由图1可知,以第 i个单根(rt i)的根尖位置为球心,半径为ρ 的球形区域称为排斥区。排斥区是单根根尖和另一个单根根尖之间保持的最小距离。单根根尖将在该区域中远离它的邻居。如果在排斥区内没有检测到邻域单根,则以第 i个单根(rt i) 的根尖位置为球心、 γ 为半径构造一个对齐区。该区域内单根与其他单根趋向方向一致。
在排斥区,能够获得第 i个单根(rt i)的邻居单
根集C r。它们之间的相关作用如式(5)所示:
D C=-∑j≠ij∈C rrt j-rt i|rt j-rt i|。(5)
式中:D C是第i个单根和其邻居单根集合C r之间的排斥力;rt j定义第j个单根的根尖位置。如果在排斥区没有监测到邻居单根(C r为空集合),则能够获得对齐区内邻居单根集合 (A r ),它们之间的相关作用如式(6)所示:
D A=∑j≠ij∈A rrt j-rt i|rt j-rt i|+∑j≠ij∈A rD j|D j|。(6)
式中:D A是第i个单根和其邻居单根集合(A r)之间的对齐力;D j 表示第j 个单根的生长方向。
单根偏好是由单根感知到的土壤环境资源分布决定的。由图1可知,排斥区内分布的资源粒子被单根吸收。构造以第 i个单根(rt i)的根尖位置为锥心、γ为半径、θ为锥角的资源粒子感知锥形区域。当资源粒子位于排斥区中时,该粒子从土壤中移除,被添加到单根的局部资源获取池中。当资源粒子位于锥形区域中而不在排斥区中时,该资源粒子影响单根的生长偏好。偏好力(D P )通过式(7)计算:
D P=M-rt i|M-rt i|。(7)
式中:M 表示位于锥形区域中而不在排斥区中时资源粒子的平均位置。
2 仿真结果
2.1 土壤环境设置
土壤中环境资源采用球形粒子表示。在0.5 m×1.0 m×0.5 m的虚拟土壤中,土壤按照厚度0.05 m被分成一定数量的土壤水平层。然后在每个水平层设置一定数目的环境资源粒子来模拟土壤中资源的分布。本研究模拟3种不同的土壤环境资源分布来分析群体行为模型对根系生长的影响。(1)随机分布。在随机资源分布中,资源粒子随机分布在每个土壤水平层中。(2)分层分布。在分层分布中,土壤层分为4个部分,每个部分有5个水平层。在第1、第3部分中,资源粒子随机分布在每個水平层中;在第2、第4部分中,数目减半的资源粒子粒子随机分布在每个水平层中。(3)梯度分布。在梯度分布中,资源粒子的数量从虚拟土壤的顶部到底部逐渐减少。
2.2 结果分析
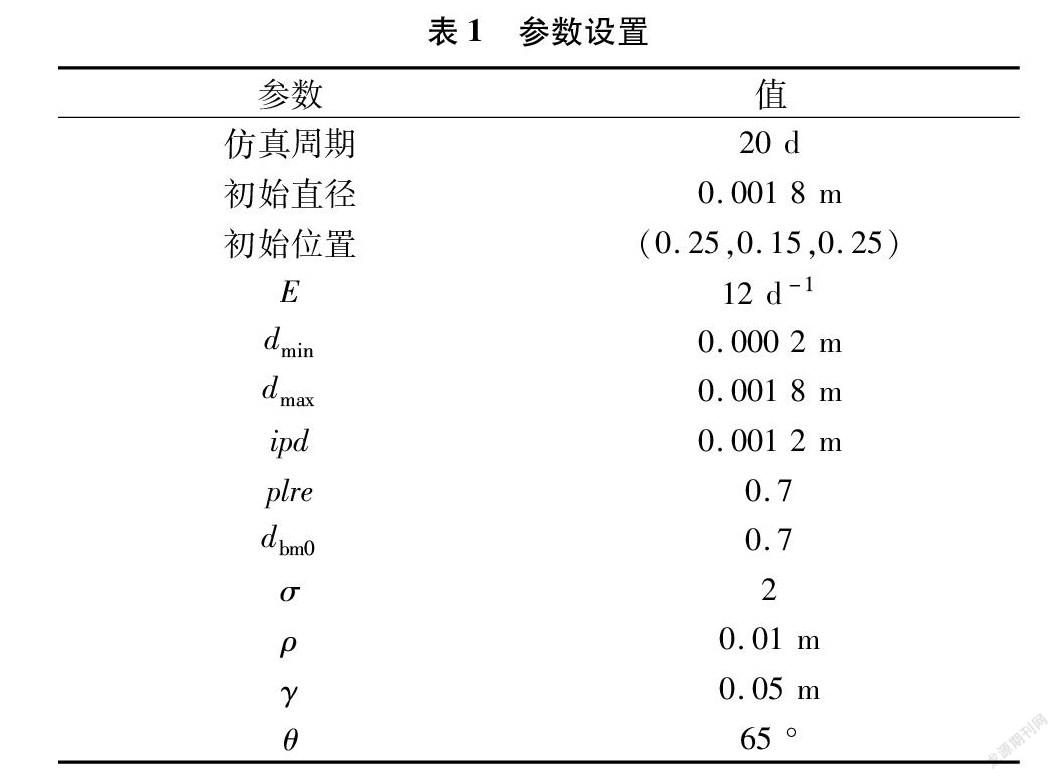
本研究仿真分析了在土壤资源随机、分层、梯度分布下群体行为、随机生长[11]、向性[13]3种机制对根系结构的影响,针对每种土壤资源分布和每种生长机制,进行了30次仿真,根生长周期设置为 20 d,仿真使用的参数见表1。采用方差分析对30个模拟的数据进行分析;采用 t检验(P <0.05)比较3种机制的差异。 t 检验的数据为每次仿真的最后一个模拟日的根生长数据。
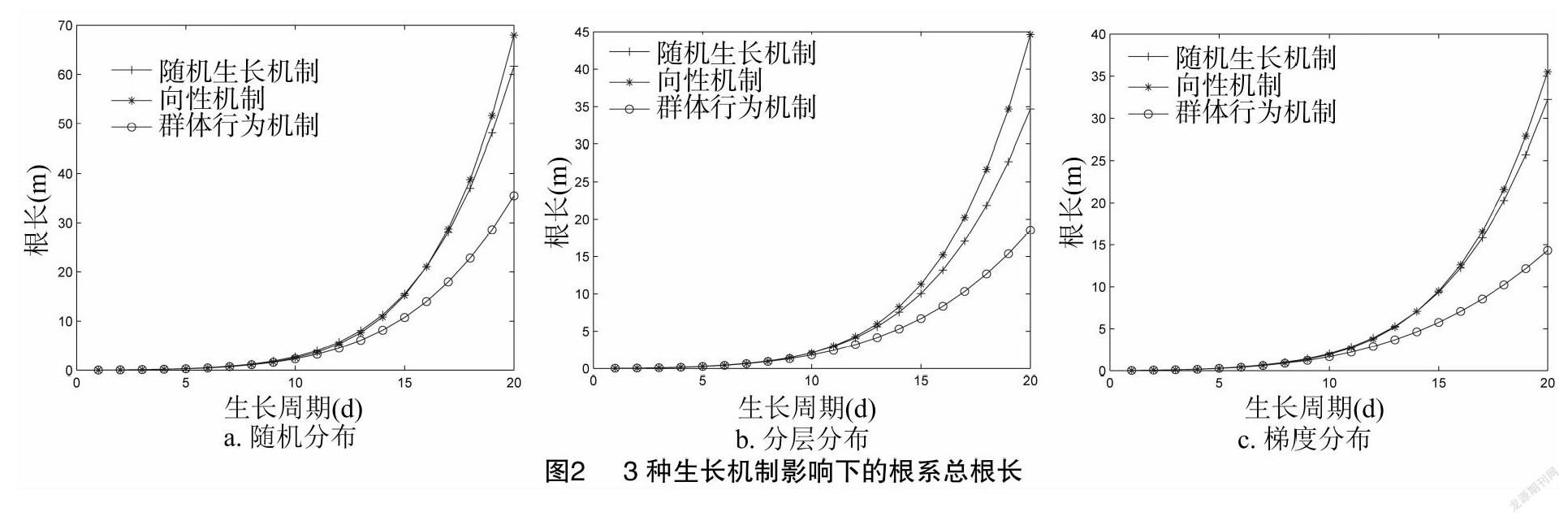
由图2可知,在土壤资源随机分布下,能够获得较长的根,而在分层和梯度分布下,总根长相对较短。在3种土壤资源分布中,向性机制获得了最长的根系,群体行为机制产生最短的根系。根据方差分析,在3种土壤资源分布下,3种生长机制产生的总根长没有显著差异。但是基于 t 检验,3种生长机制在3种土壤分布中产生的总根长有显著的不同。
由图3可知,在3种土壤资源分布下,向性机制总是获得最大根体积,群体行为机制获得最小根体积。根据方差分析,在3种土壤资源分布下,3种生长机制产生的根体积没有显著差异。基于 t 检验,在土壤资源随机分布和梯度分布下,随机生长机制和向性机制没有显著差异。在其他情况下,3种生长机制有显著的不同。
在土壤资源随机分布、分层分布和梯度分布下,群体行为机制、随机生长机制、向性机制对根系分布(不同土壤深度的器官数量)的影响见图4。向性机制产生最深的根,在土壤资源随机分布、分层分布和梯度分布下,分别达到了0.75、0.65、0.70 m。在土壤资源分层分布下,随机生长机制有54.9%的根系位于土壤上部0.25 m内,而向性和群体行为机制分别有54.4%、32.7%的根系位于该区域;随机生长机制有45.0 %的根系位于0.25~0.50 m的土壤中,而向性和群体行为机制分别有44.8%、67.1%的根系位于该区域。在土壤资源梯度分布下,在虚拟土壤的顶部0.30 m,随机生长机制、群体行为机制、向性机制中分别产生84.6%、58.3%、81.4%根系分布。基于方差分析,在3种土壤资源分布下,3种生长机制产生的根分布没有显著差异。
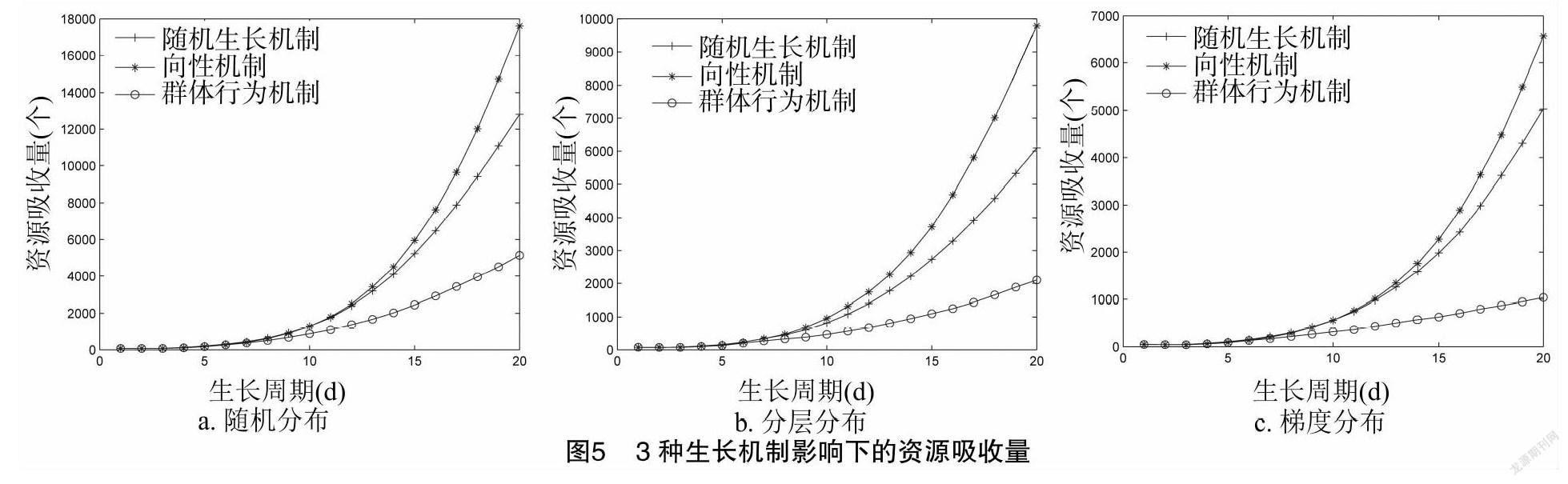
由圖5可知,群体行为机制影响下的根系比其他2种机制影响下的根系明显资源吸收量较低。例如,在土壤资源随机分布中,群体行为机制影响下的根系资源吸收量分别为随机生长机制、向性机制影响下的根系资源吸收量的46.3%、37.9%。在土壤资源分层分布中,群体行为机制影响下的根系资源吸收量分别为随机生长机制和向性机制影响下的根系资源吸收量的41.0%、28.6%。在土壤资源梯度分布中,群体行为机制影响下的根系资源吸收量分别为随机生长机制和向性机制影响下的根系资源吸收量的30.8%、25.8%。基于方差分析,在土壤资源随机分布下,3种机制的根系资源吸收量没有显著差异,但在土壤资源分层和梯度分布下,3种机制的根系资源吸收量存在显著差异。根据 t 检验结果,3种机制在3种土壤资源分布下均存在显著差异。
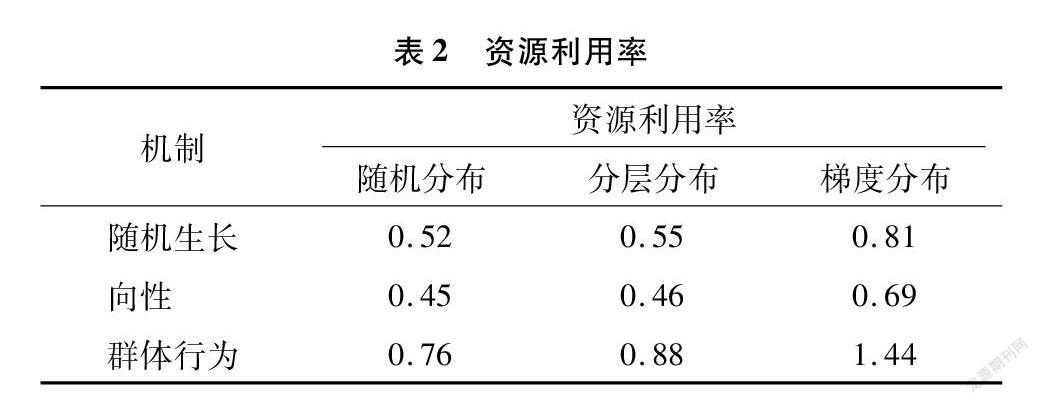
依据对3种不同土壤资源下3种不同生长机制控制的根系特征数据(总根长、根体积、根分布和资源吸收量)的方差分析,在土壤资源分层和梯度分布下,3种机制的根系资源吸收量存在显著差异,在其他情况下总根长、根体积、根分布没有显著差异。依据对根系特征数据的 t 检验,发现3种不同生长机制有显著差异。分析结果表明,根系可能采用这3种机制中的1种或多种来适应复杂的环境。与随机生长机制和向性机制不同,群体行为机制影响下的根系具有高的资源利用率(资源利用率=根体积/总资源吸收量)(表2)。仿真结果表明群体行为机制可能是根系提高资源利用效率的策略之一,结果与Wang等的研究结果[14]一致。
3 结论
本研究提出了一种集成ArchiSimple模型受根系群体行为影响的根系生长模型。该模型可用于研究根系生长发育决策过程中的机制。3种不同土壤环境设置对3种不同生长机制影响的根系特征数据的比较分析结果表明,群体行为能够提升根系资源利用效率,为进一步研究群体行为对根系生长发育的影响提供基础。
参考文献:
[1]段海霞,罗崇亮,Minha N,等. 优化作物根系构型,发展生态农业:2017年作物根系与根际互作国际研讨会综述[J]. 干旱地区农业研究,2019,37(2):1-9,51.
[2]苗青霞,方 燕,陈应龙.小麦根系特征对干旱胁迫的响应[J]. 植物学报,2019,54(5):652-661.
[3]潘晓迪,张 颖,邵 萌,等. 作物根系结构对干旱胁迫的适应性研究进展[J]. 中国农业科技导报,2017,19(2):51-58.
[4]Araya T,Kubo T,von Wirén N,et al. Statistical modeling of nitrogen-dependent modulation of root system architecture in Arabidopsis thaliana [J]. Journal of Integrative Plant Biology,2016,58(3):254-265.
[5]Diggle A J.ROOTMAP—a model in three-dimensional coordinates of the growth and structure of fibrous root systems[J]. Plant and Soil,1988,105(2):169-178.
[6]Schnepf A,Leitner D,Landl M,et al. CRootBox:a structural-functional modelling framework for root systems[J]. Annals of Botany,2018,121(5):1033-1053.
[7]Postma J A,Kuppe C,Owen M R,et al. OpenSimRoot:widening the scope and application of root architectural models[J]. New Phytologist,2017,215(3):1274-1286.
[8]Ciszak M,Comparini D,Mazzolai B,et al. Swarming behavior in plant roots[J]. PLoS One,2012,7(1):e29759.
[9]Matos T,Cruz C,Correia L.Root growth model based on swarm intelligenceroot engineering[M]. Berlin:Springer,2014.
[10]McCleery W T,Mohd-Radzman N A,Grieneisen V A.Root branching plasticity:collective decision-making results from local and global signalling[J]. Current Opinion in Cell Biology,2017,44:51-58.
[11]Pagès L,Bécel C,Boukcim H,et al. Calibration and evaluation of ArchiSimple,a simple model of root system architecture[J]. Ecological Modelling,2014,290:76-84.
[12]Couzin I.Collective minds[J]. Nature,2007,445(7129):715.
[13]Henke M,Sarlikioti V,Kurth W,et al. Exploring root developmental plasticity to nitrogen with a three-dimensional architectural model[J]. Plant and Soil,2014,385(1/2):49-62.
[14]Wang X R,Shen J B,Liao H.Acquisition or utilization,which is more critical for enhancing phosphorus efficiency in modern crops?[J]. Plant Science,2010,179(4):302-306.
