外源脱落酸缓解UV-B胁迫对牛皮杜鹃气孔及光合特性的影响
2022-05-09李悦张庆怡董佳伟徐洪伟周晓馥
李悦 张庆怡 董佳伟 徐洪伟 周晓馥



摘要:为探讨外源脱落酸(abscisic acid,简称ABA)缓解紫外线-B(UV-B)胁迫对植物气孔及光合特性的影响。本试验以牛皮杜鹃( Rhododendron chrysanthum Pall)为试验材料,共设PAR、UV-B、 ABA+PAR及 ABA+UV-B等4组处理,测定气孔、花青素(anthocyan)含量以及叶绿素荧光参数,并进行对比分析。结果表明,UV-B胁迫使牛皮杜鹃气孔特征、花青素含量、 F m、 F v/ F m、 F v′/ F m′、 F v/ F o、 q P显著降低( P <0.05), q P快速光响应曲线下降幅度增大, ABA预处理显著提高了花青素的含量、 F m、 F v/ F m、 F v′/ F m′、 F v/ F o、 q P( P <0.05),且使 q P、NPQ快速光响应曲线呈现出最佳状态。这些结果证实了外源ABA能缓解UV-B对牛皮杜鹃光合系统造成的损伤,提高了其光合能力,从而提高了牛皮杜鹃对UV-B的耐受能力。
关键词:牛皮杜鹃;UV-B;外源脱落酸;花青素;叶绿素荧光参数
中图分类号: S685.210.1 文献标志码: A
文章编号:1002-1302(2022)07-0145-07
收稿日期:2021-07-09
基金项目:吉林省科技发展计划(编号:20190301059NY);中央指导地方科技发展资金吉林省基础研究专项(编号:202002016JC)。
作者简介:李 悦( 1998—),女,吉林白城人,硕士研究生,研究方向为植物遗传学。E-mail:13194363751@163.com。
通信作者:周晓馥,博士,教授,博士生导师,主要从事植物分子生物学研究。E-mail:zhouxiaofu@jlnu.Edu.cn。
牛皮杜鹃( Rhododendron chrysanthum Pall),别称牛皮茶,是一种具有重要观赏和药用价值的濒危物种,是我国长白山的特有物种,也是研究植物耐逆性的重要植物资源[1]。长白山环境恶劣,空气稀薄,太阳辐射强烈,其中紫外线-B(UV-B)辐射是该地区主要的非生物胁迫因子[2]。
UV-B脅迫会对植物的生理代谢造成影响[3]。孟凡来等都指出,UV-B辐射胁迫可造成叶绿体结构的破坏或使叶绿素发生光氧化,从而导致叶绿素含量的下降[4-5];徐超群表示,UV-B辐射会导致叶绿素和类胡萝卜素上的杂环与共轭双键产生活性氧,进而破坏细胞稳态,与此同时,类胡萝卜素的含量在UV-B的辐射下,也会发生显著变化[6]。研究显示,类胡萝卜素分子所含有的共轭双键既能参与光能的捕获又可耗散过剩光能,能够防御光氧化对叶绿素的不利影响,同时也导致了自身含量的减少[4-5,7]。
脱落酸(abscisic acid,简称ABA)具有引起休眠、抑制生长、促进脱落的生理作用。研究证明,ABA在植物遭受干旱、低温、盐渍等不良环境胁迫时可快速做出响应,其含量的增加能很好地增强植物的防御能力,因此又称ABA为“胁迫激素”[8-9]。王娟通过研究发现,外源ABA可以改善在干旱条件下的烟草幼苗的叶黄素循环,从而耗散了过剩的激发能,在一定程度上提高了烟草幼苗的抗旱性[10]。刘晓龙发现,外源 ABA 预处理显著抑制了碱胁迫导致的根系活性氧(ROS)过量积累,提高抗氧化酶活性并缓解碱胁迫导致的水稻根系损伤和幼苗死亡[11]。目前,对外源ABA增强牛皮杜鹃抗UV-B胁迫能力的研究还比较少。
气孔作为植物体和外界进行H 2O和CO 2交换的重要介质,当植物遭受胁迫时会通过调整气孔的开度等特征来抵御损伤,这也是植物对于胁迫耐受能力的体现。McAinsh等发现,脱落酸能够通过Ca2+依赖和Ca2+非依赖途径调节气孔大小,适应外部逆境[12-13]。然而,目前有关外施脱落酸对逆境胁迫下作物苗期叶片气孔特征和光合性能的研究还鲜见报道。通常外施脱落酸能够诱导气孔关闭和气孔密度,降低气孔导度,从而影响光合速率。
类黄酮(flavonoids)是植物中再生代谢产物的主要类型,它主要以糖苷的形式存在于植物液泡中,莫运才通过研究发现,在低UV-B辐射下,植物会诱导类黄酮合成,随着UV-B辐射强度的增加,植物会消耗类黄酮以抵御损伤[14]。
叶绿素荧光参数已经被认为是衡量植物受 UV-B 辐射伤害及植物抗性的重要指标。本试验采用PAM荧光计进行测量叶绿素荧光的各项参数,试验主要研究的参数如表1所示。
本研究以牛皮杜鹃为材料,探究外源ABA对牛皮杜鹃在UV-B胁迫下气孔、花青素含量以及叶绿素荧光参数所受影响的缓解效应,以期为ABA对植物的抗辐射功能提供理论支持,同时为外施ABA以增强植物耐受性与抗性提供可行方案与指导。
1 材料与方法
1.1 试验材料
牛皮杜鹃采自长白山(40.10° N、100.10° E),用 1/4 MS 培养基置于吉林师范大学吉林省植物资源科学与绿色生产重点实验室模拟高山植物智能人工气候室中正常培养[光暗周期为14 h/10 h;白天/夜晚温度为18 ℃/16 ℃;光子通量密度为 50 μmol/(m2·s)]。本试验选取 8 个月大且长势一致的牛皮杜鹃组培苗为试验材料。
1.2 试验方法
1.2.1 材料处理 选用相同生长状态的牛皮杜鹃组培苗作为试验材料。共设4组处理,分别记为PAR、UV-B、ABA+PAR、ABA+UV-B,每组处理均设置3次生物学重复。第1组与第2组采用空白处理的1/4 MS培养基对牛皮杜鹃进行移栽,第3组与第4组采用另添加ABA(100 μmol/L)的1/4 MS培养基对牛皮杜鹃进行移栽,移栽后置于人工气候室培养6 d后,将第2组与第4组植物于UV-B辐射(2.3 W/m2)下胁迫8 h/d,UV-B辐射处理2 d后进行各项指标的测定。本试验利用人造UV-B辐射灯管进行UV-B辐射,灯管的波长发射范围为280~320 nm。于进行UV-B处理的牛皮杜鹃上方放置295 nm滤光片(Edmund,Filter Long 2IN SQ,USA)以排除干扰;于不进行UV-B处理的牛皮杜鹃上方放置400 nm滤光片(Edmund,Filter Long 2IN SQ,USA)以过滤辐射。
1.2.2 牛皮杜鹃气孔特征的测定
取牛皮杜鹃叶片中部靠叶脉处2 mm×2 mm 小块,将透明指甲油均匀涂抹于叶片背面分别采集叶片气孔,制成临时装片,每个处理制作 3 张临时装片,每张临时装片里随机选择 5 个视野,利用显微摄像系统(Model C-SHG 1,Nikon Corp,日本)进行观察和拍照,每个处理得到15张气孔结构图片,随机选取 3 张图片。然后,利用Image J软件分别测量气孔的长度、宽度、周长和面积,进一步计算气孔形状指数。气孔形状指数的计算公式为 S=P/2 π A (以圆为参照几何形状)。式中:S为叶片气孔形状指数;P表示气孔周长;A表示气孔面积。
1.2.3 牛皮杜鹃花青素含量的测定
花青素含量用分光光度法测定。将新鲜叶片按照25 ∶9的比例在甲醇与盐酸(体积比为99 ∶1)的混合液中组成细胞匀浆,室温下黑暗中提取3 d,在546 nm处读取提取物的吸光度以测定花青素含量,吸光度用叶片处于新鲜状态时的质量表示。
1.2.4 牛皮杜鹃PS Ⅱ叶绿素荧光参数测定及相关参数计算
利用IMAGING-PAM 叶绿素荧光成像系统(Heinz Walz,德国)对其进行叶绿素荧光参数的测定,在相同叶龄的叶片上选取 3 个观测点,暗适应 20 min 后对初始荧光( F o)、最大荧光( F m)、光下最大荧光( F m′)以及光下最小荧光( F o′)进行测定并计算以下参数,同时对 q P、NPQ 和电子传递效率(ETR)快速光响应曲线进行测定。
光系统Ⅱ(PS Ⅱ)的最大量子产量: F v/ F m=( F m- F o)/ F m;
光系统Ⅱ(PS Ⅱ)的有效量子产量: F v′/ F m′=( F m′- F o′)/ F m′;
光系统Ⅱ(PS Ⅱ)的潜在活性: F v/ F o=( F m- F o)/ F o;
光化学淬灭系数: q P=( F m′- F s)/( F m′- F o′);
非光化学淬灭系数:NPQ=( F m- F m′)/ F m′。
1.2.5 數据统计与分析
采用统计软件SPSS 22.0对试验数据进行差异显著性分析( P <0.05),利用非线性拟合曲线对参数进行计算,采用Sigmaplot 12.5软件进行作图。
2 结果与分析
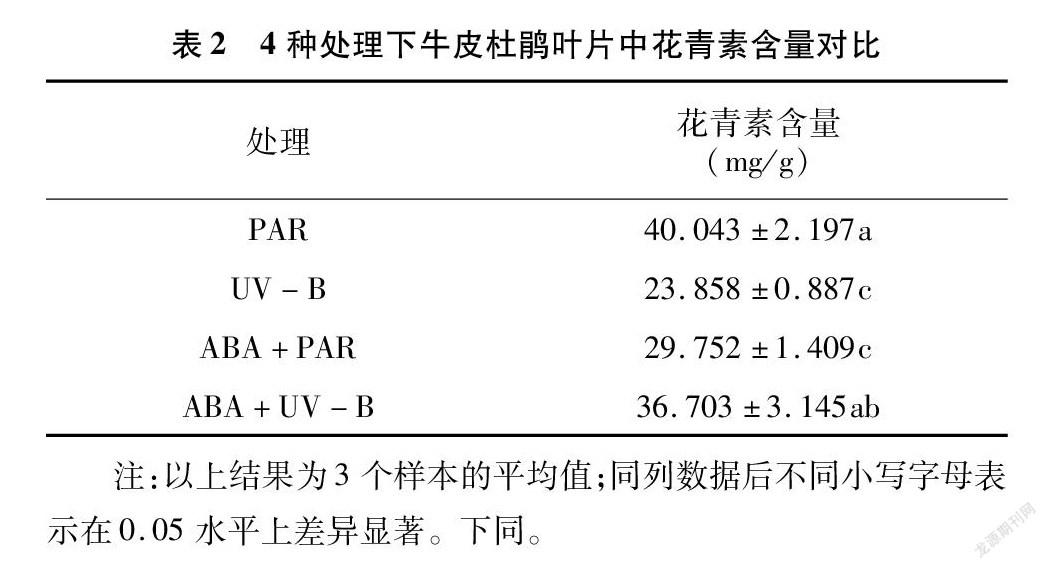
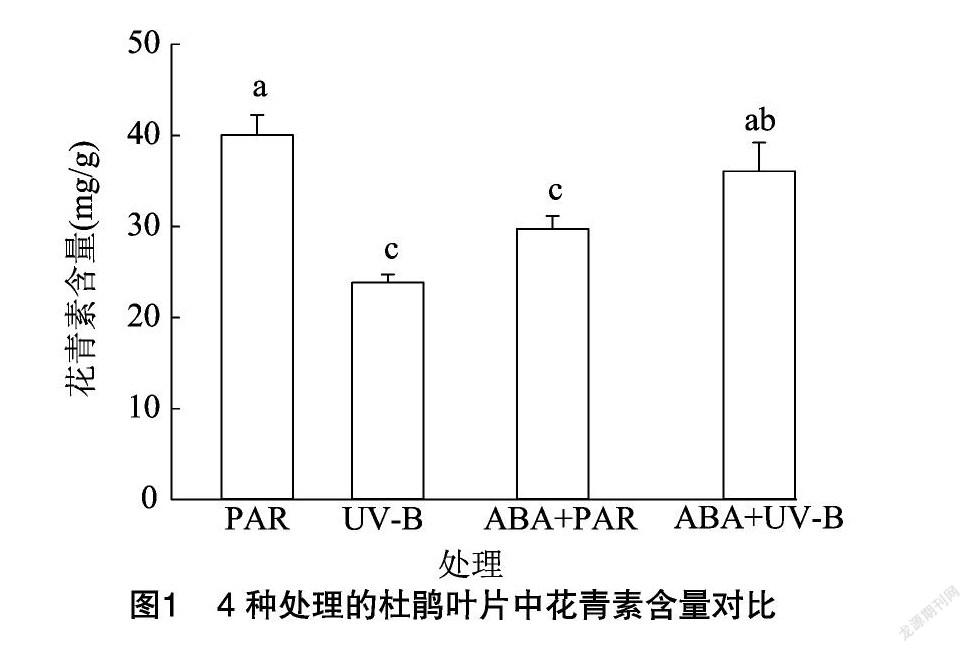
2.1 外源ABA与UV-B胁迫对牛皮杜鹃花青素含量的影响
通过对数据进行分析发现,与PAR组相比,UV-B 辐射处理显著降低了花青素含量( P <0.05),数值下降40.42%;ABA+PAR组的花青素含量显著降低( P <0.05),数值下降25.70%;经过UV-B、ABA双重处理的牛皮杜鹃表现出了更高的花青素含量,为36.703 mg/g(表2、图1)。花青素作为公认的抗紫外化合物,在植物抗紫外辐射中起到至关重要的作用,高强度的紫外辐射消耗了植物体中花青素的含量,降低了植物的色素含量,破坏了植物叶绿体,从而影响植物正常的光合作用。ABA单一处理使牛皮杜鹃花青素含量降低,但在ABA和UV-B的双重作用下提高了花青素含量水平,说明ABA通过调控花青素含量来保持牛皮杜鹃体内色素含量的稳定,增强其光合能力,具有缓解UV-B胁迫对牛皮杜鹃的负面影响的作用。
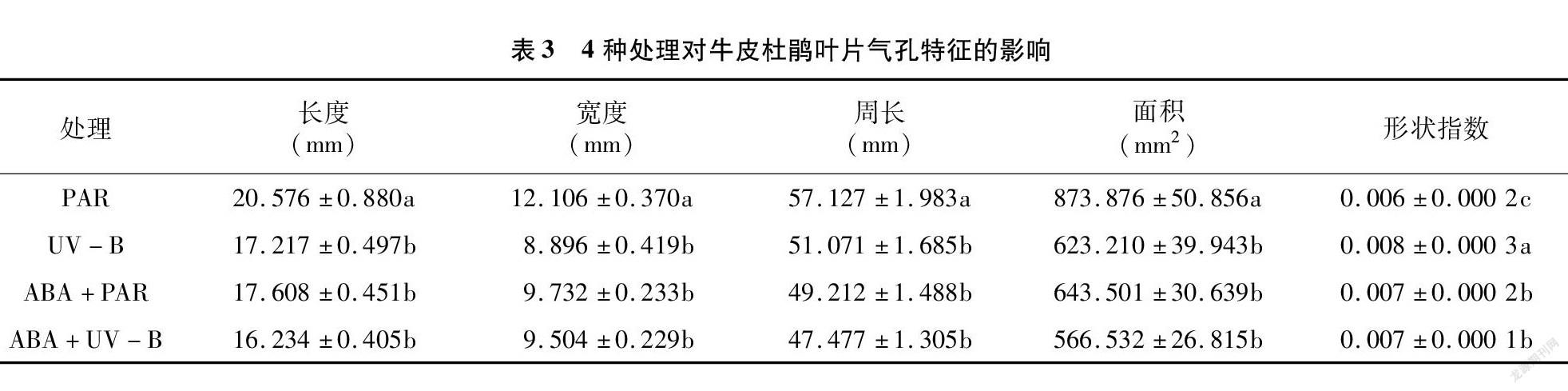
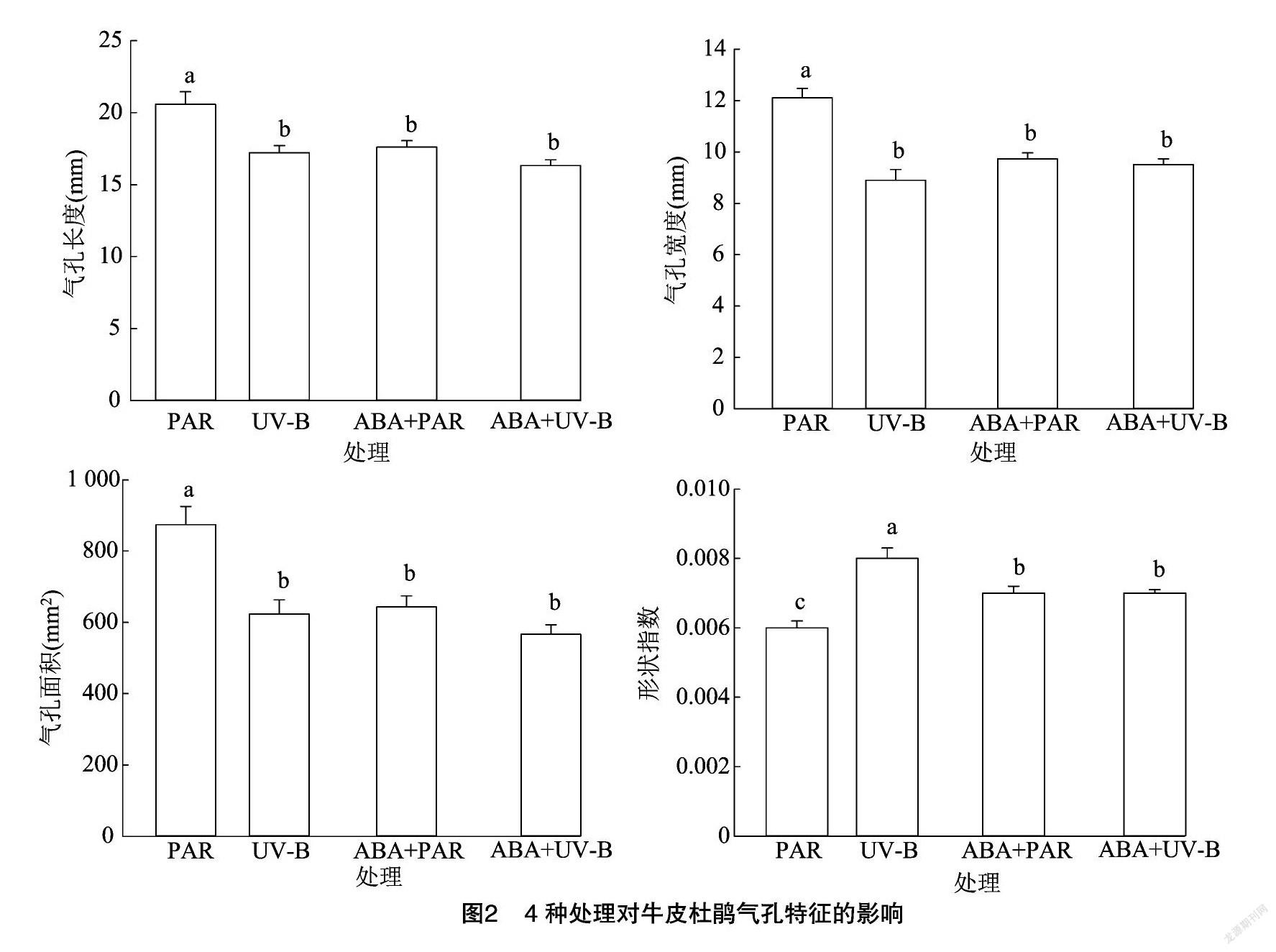
2.2 外源ABA与UV-B胁迫对牛皮杜鹃气孔的影响
为探究UV-B胁迫对气孔的影响,制作牛皮杜鹃气孔临时装片,在显微镜成像系统下观察并拍摄照片进行测量及数据分析。与PAR组进行对比,UV-B胁迫显著降低了气孔长度、宽度、 周长及面积,
数值分别降低16.32%、26.52%、11.85%、28.68%;UV-B胁迫同时显著增高了形状指数( P <0.05),数值升高33.33%;与PAR组相比,ABA+PAR组的气孔特征变化显著,气孔长度、宽度、周长及面积均显著下降( P <0.05),数值分别降低14.43%、19.61%、13.86%、26.35%;与UV-B组相比,ABA+UV-B组的气孔特征除宽度外均下降,但并不显著( P >0.05)。气孔作为植物和外界交换H 2O 和 CO 2的重要介质,当植物遭受胁迫后会通过调整气孔特征以抵御损伤。当牛皮杜鹃遭受UV-B胁迫后,气孔长度、宽度、周长及面积等指标均显著降低,气孔开度减小,从而减弱蒸腾作用减少水分散失,UV-B引起过多的光抑制,以致过多的热量无法通过蒸腾作用散发到外界环境中。外源ABA通过增加气孔宽度提高蒸腾速率,使由 UV-B 造成的光抑制而无法散出的热量散发,提高光合速率,并使CO 2更易通过气孔扩散进入到牛皮杜鹃叶片组织内部,影响光合作用的最大反应速率,从而提高牛皮杜鹃对UV-B胁迫的耐受性(表3、图2)。
2.3 外源ABA与UV-B对牛皮杜鹃叶绿素荧光图像的影响
为探究ABA与UV-B胁迫对牛皮杜鹃叶绿素荧光图像的影响,将2组材料分别进行PAR和UV-B辐射处理,在胁迫前后分别进行叶绿素荧光参数的测定。图3显示了在PSⅡ光系统中经过各种处理的牛皮杜鹃的 F o和 F m图像。 其中 F o是初始荧光值,其数值大小取决于PSⅡ光系统中心是否受到损伤,当PSⅡ光系统反应中心遭到破坏时, F o数值会提高。 F m是荧光的最大值,是处于黑暗适应状态下关闭植物光反应中心时的荧光强度。 F m数值反映PSⅡ光系统电子传递的情况,是体现植物承受胁迫能力的一个重要特性。由图3可知,与PAR组相比,牛皮杜鹃在经受UV-B胁迫后,生长受到限制,其生长态势并无明显变化,说明UV-B胁迫使牛皮杜鹃的生长速度减缓;并且通过图像可以发现,在经受UV-B胁迫后,牛皮杜鹃的发病区域多为叶片,叶片承受UV-B胁迫,这也体现了 UV-B 胁迫最初以及最主要的作用目标便是光合作用。
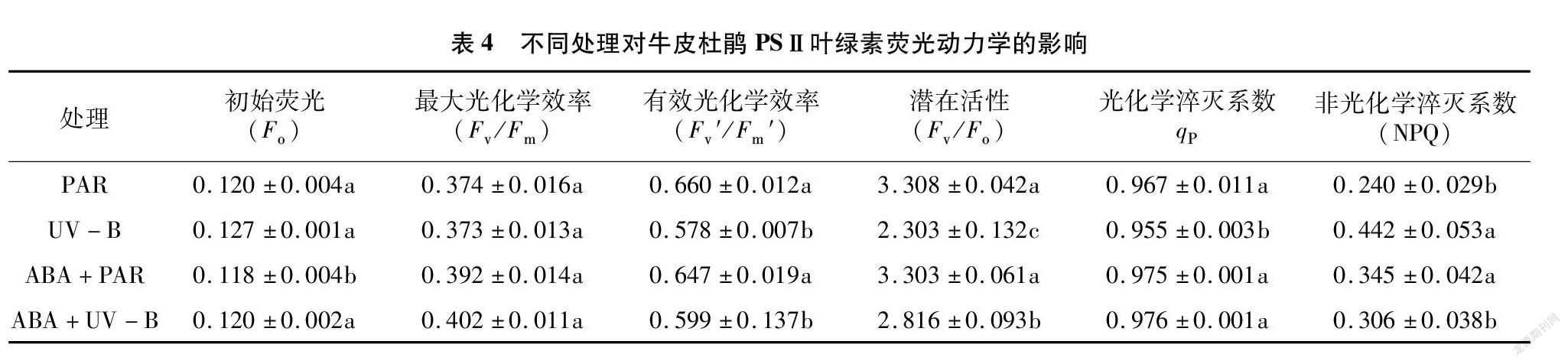
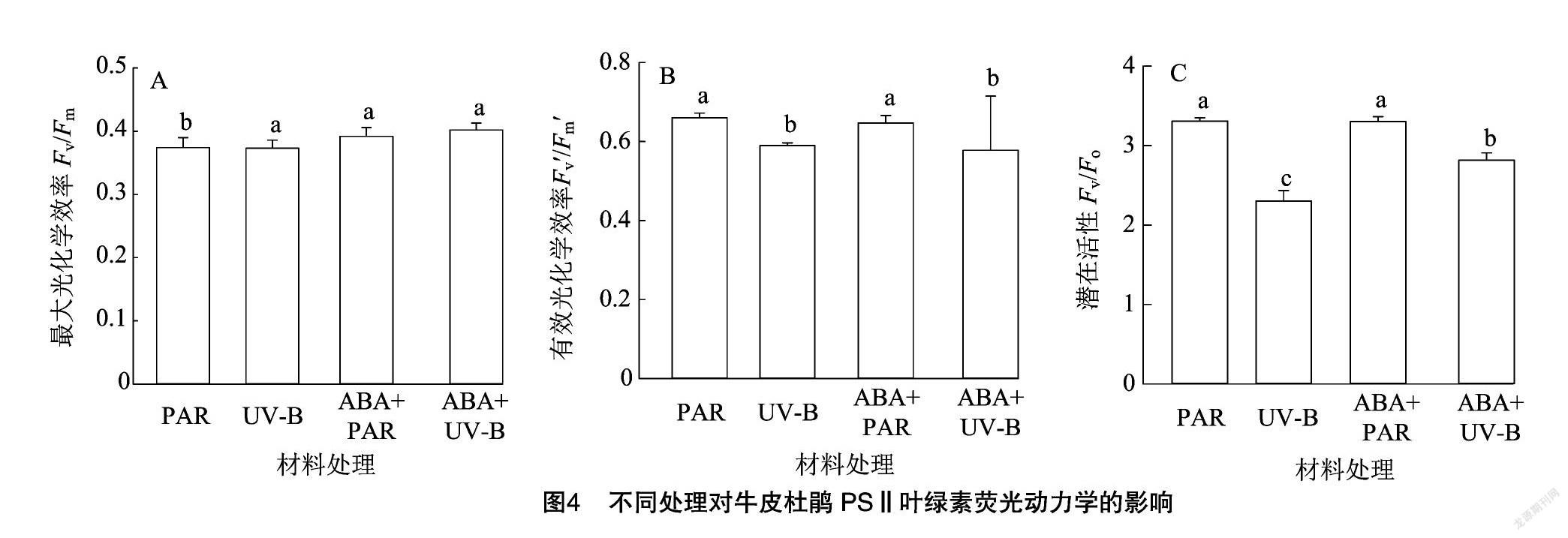
2.4 外源ABA与UV-B胁迫对牛皮杜鹃PSⅡ叶绿素荧光动力学的影响
2.4.1 UV-B胁迫对牛皮杜鹃PSⅡ叶绿素荧光动力学的影响
F o为初始荧光,其数值大小取决于 PSⅡ 光系统中心是否受到损伤,当PSⅡ光系统反应中心遭到破坏时, F o提高。PSⅡ光系统的最大光化学效率( F v/ F m)也是原初光能转化效率,反映PSⅡ光系统活性,以及植物在受逆境胁迫下的光抑制程度。当光合系统吸收过量的光能时,PSⅡ 光系统活性就会下降,这种现象称为“光抑制”。 F v′/ F m′为PSⅡ光系统有效光化学效率,可以将捕获的光能转化为生物所需的化学能,反映了PSⅡ光系统反应中心原初光能的捕获效率。 F v/ F o作为植物PSⅡ光系统的潜在活性,其变化反映出植物在受到胁迫后光合生理活性的变化。通过分析得知,与PAR组相比,UV-B组 F o值显著升高( P <0.05), F v/ F m的值降低但无显著影响( P >0.05), F v′/ F m′、 F v/ F o的值显著降低( P <0.05)(表4、图4)。说明牛皮杜鹃在受到UV-B胁迫后,初始荧光和有效光化学效率降低,光能利用率降低,导致PSⅡ光系统潜在活性受损,最终导致光化学电子进入碳还原过程受到抑制,降低了牛皮杜鹃的光合能力。
q P是光化学猝灭系数,反映了PSⅡ光系统反应中心开放的比例和参与碳固定转移的电子, q P越大,说明植物进行光合作用利用的光能越多。NPQ是非光化学猝灭系数,反映了植物PSⅡ天线色素吸收光能后不能用于光化学电子传递,而以热能形式耗散掉未利用光能的部分。与PAR组相比,UV-B组的 q P显著降低,NPQ显著升高。 q P的降低说明 PSⅡ光化学电子传递活性降低,与 F v/ F m、 F v′/ F m′ 和 F v/ F o 的下降相吻合。而NPQ的升高,说明 UV-B 引起了过多的光抑制(表4)。
2.4.2 外源ABA缓解UV-B胁迫对牛皮杜鹃 PSⅡ 叶绿素荧光动力学的影响
与PAR组相比,外源ABA处理后初始荧光、最大光化学效率、有效光化学效率无明显改变,与PAR组持平;与UV-B组相比,在ABA+UV-B处理后,牛皮杜鹃的 F o降低,但并无显著差异( P >0.05), F v/ F m、 F v′/ F m′ 、 F v/ F o以及 q P显著提高( P <0.05),并与PAR组基本持平,而NPQ与PAR组相比显著升高( P <0.05)。这些结果都说明外源ABA可以提高牛皮杜鹃PSⅡ的光合电子传递,从而缓解了UV-B胁迫对牛皮杜鹃的负面影响(表4)。
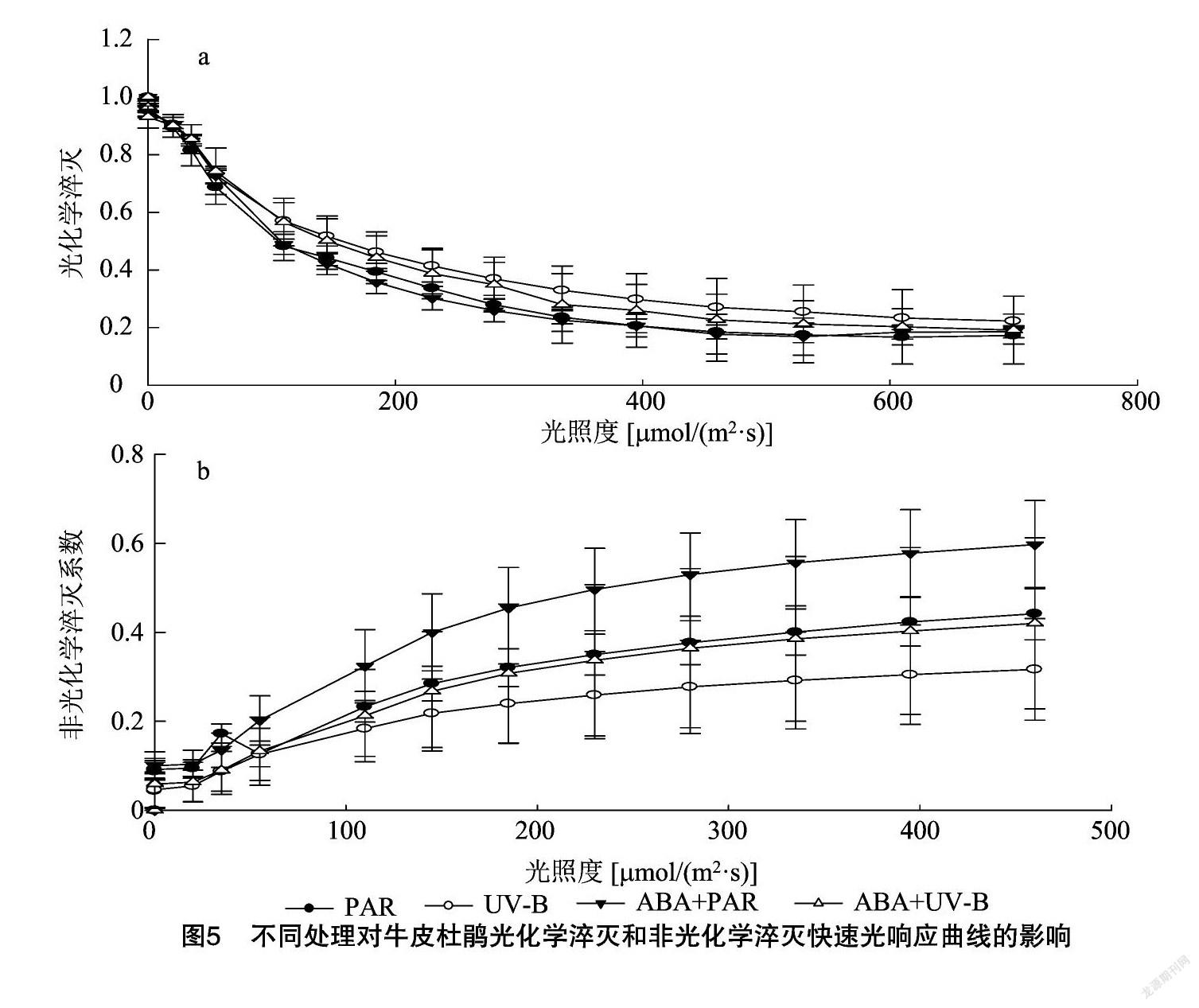
2.5 外源ABA与UV-B胁迫对牛皮杜鹃PSⅡ叶绿素荧光快速光响应曲线的影响
2.5.1 外源ABA缓解UV-B胁迫对牛皮杜鹃 PSⅡ q P快速光响应曲线的影响
q P是光化学淬灭系数,反映了PSⅡ反应中心开放的比例和参与了碳固定转移的电子, q P数值越大说明植物进行光合作用利用的光能越多。对结果进行分析可知,不同处理的 q P快速光响应曲线呈现出不同的变化趋势,与PAR相比,UV-B辐射处理后 q P快速光响应曲线呈现明显的下降趋势,且随着光照度的增加,其 q P下降的幅度最小,这与 q P动力学数值的下降表现一致,且随着光照度的增加,ABA 预处理的 q P 快速光响应曲线呈现出最佳状态,在光照度不断增加的过程中,UV-B处理的 q P下降的幅度最小。由此可见,经UV-B胁迫的牛皮杜鹃光合能力变弱,对光能的利用率大大降低,而ABA能够提高光能利用率,从而缓解UV-B的负面影响(表4、图5)。
2.5.2 外源ABA与UV-B胁迫对牛皮杜鹃PSⅡNPQ快速光响应曲线的影响
植物处于UV-B胁迫时,PSⅡ天线色素吸收UV-B的高能光量子,不能用于光合电子传递的过量激发能以熱能形式淬灭,即非光化学淬灭,其份额可由非光化学淬灭系数NPQ来反映。通过对结果进行分析可知,与PAR组相比,牛皮杜鹃在受到UV-B胁迫后,NPQ快速光响应曲线呈现下降趋势,这与NPQ数值的显著降低( P <0.05)表现一致(表4、图5)。说明UV-B胁迫导致了牛皮杜鹃光合能力的显著降低,引起了过多的光抑制,出于自我保护,牛皮杜鹃使NPQ上升,将不能利用的光能以热能的形式散失以求部分减轻因PSⅡ吸收光能过多而导致的光氧化。
与PAR组相比,ABA+PAR组NPQ显著上升( P <0.05);而与UV-B组相比,ABA+UV-B组NPQ显著降低( P <0.05)。并且NPQ快速光响应曲线在不同处理下随着光照度的增大而呈现出不同趋势,其上升幅度表现为ABA+PAR>PAR>ABA+UV-B>UV-B(表4、图5)。这说明外源ABA预处理后可使NPQ快速光响应曲线上升,即减轻了牛皮杜鹃PSⅡ中过剩光能的光抑制效应,进而增强光保护能力。
3 讨论与结论
植物在受到UV-B辐射胁迫时,会产生各种适应、伤害与修复机制。梁东雨等研究发现,UV-B辐射会使植物光系统Ⅱ受损,使光合作用的原初反应过程受到抑制[2]。陈娟等研究发现,ABA可以诱导叶片气孔开度受抑或气孔关闭、蒸腾作用下降,因而水分耗散减少,最终使植物保水能力和对干旱的耐受性提高;在外源ABA作用下可加强活性氧的清除,减缓膜脂过氧化对膜系统造成的损伤,从而维护光合系统的正常功能[16]。
本试验中,UV-B胁迫限制了牛皮杜鹃的生长发育过程,具体表现为植株气孔特征的降低,叶片的 F v′/ F m′、 F v/ F o显著降低。然而,外施ABA可以通过调整气孔的形态特征及其空间分布格局有效缓解了UV-B胁迫对牛皮杜鹃造成的负面影响。牛皮杜鹃在单一的UV-B辐射和外施ABA下其光合色素含量并没有显著变化,而ABA与UV-B双重处理的花青素含量却发生了显著变化,花青素作为次生代谢物质类黄酮的一种,是植物抵御UV-B胁迫的主要物质,花青素含量的升高是牛皮杜鵑对UV-B的一种适应。在UV-B胁迫下,牛皮杜鹃PSⅡ的 F v/ F m受到影响但并不显著,而 F v′/ F m′、 F v/ F o数值均显著降低,使潜在活性降低,导致光化学电子进入碳还原过程受到抑制,从而引起光系统损伤,导致光合生理活性下降。而ABA可以缓解UV-B胁迫对牛皮杜鹃PSⅡ光合特性造成的负面影响。结果表明,在ABA与UV-B的双重处理下 F o数值回落以及 F v/ F m、 F v′/ F m′、 F v/ F o有所升高,并能基本恢复到PAR组水平。并且在外源ABA作用下,ABA+UV-B组的 q P和 NPQ快速光响应曲线都呈现出最佳状态。
综上所述,这些结果都证明了ABA可以缓解UV-B对牛皮杜鹃造成的负面影响,可以增强光合作用且使花青素等物质积累以减轻伤害,并帮助其更好地适应逆境。
参考文献:
[1]Zhou X F,Chen S L,Wu H,et al. Biochemical and proteomics analyses of antioxidant enzymes reveal the potential stress tolerance in Rhododendron chrysanthum Pall[J]. Biology Direct,2017,12(1):10.
[2]梁东雨,王禹佳,苏敖纯,等. 外源茉莉酸对牛皮杜鹃UV-B辐射缓解作用研究[J]. 生物技术通报,2019,35(10):64-70.
[3]李 元,祖艳群,高召华,等. UV-B辐射对报春花的生理生化效应[J]. 西北植物学报,2006,26(1):179-182.
[4]孟凡来,郭华春. UV-B辐射增强对甘薯光合特性和紫外吸收物质的影响[J]. 作物杂志,2019(5):114-119.
[5]Masih J.Effects of ultraviolet-B radiation on the photosynthetic pigments and protein content of strawberry[J]. Asian Journal of Applied Chemistry Research,2019,3(4):1-7.
[6]徐超群. 高山松高海拔适应性的光合生理学和转录组学基础[D]. 北京:北京林业大学,2019.
[7]刘羽霞. 长白山苔原带优势植物的光合特性及其对氮沉降与草本入侵的响应[D]. 长春:东北师范大学,2018.
[8]韩小强,肖玉梅,路慧哲,等. 脱落酸抗代谢与光稳定性类似物研究进展[J]. 植物学报,2013,48(3):329-343.
[9]李 俊,牛金文,杨 芳,等. 不同马铃薯品种(系)对增强UV-B辐射的形态响应[J]. 中国生态农业学报,2016,24(6):770-779.
[10]王 娟. 干旱条件下外源ABA提高烟草幼苗抗旱性的作用机制[D]. 哈尔滨:东北林业大学,2014.
[11]刘晓龙. 脱落酸(ABA)对水稻耐碱胁迫的诱抗效应及机理研究[D]. 长春:中国科学院大学(中国科学院东北地理与农业生态研究所),2019.
[12]McAinsh M R,Brownlee C,Hetherington A M.Visualizing changes in cytosolic-free Ca2+ during the response of stomatal guard cells to abscisic acid[J]. Plant Cell,1992,4:1113-1122.
[13]Li C L,Wang M,Ma X Y,et al. NRGA1,a putative mitochondrial pyruvate carrier,mediates ABA regulation of guard cell ion channels and drought stress responses in Arabidopsis [J]. Molecular Plant,2014,7(10):1508-1521.
[14]莫运才. UV-B辐射结合不同氮素营养水平对铁皮石斛生长及主要活性成分的影响[D]. 广州:广东药科大学,2016.
[15]岑海燕,姚洁妮,翁海勇,等. 叶绿素荧光技术在植物表型分析的研究进展[J]. 光谱学与光谱分析,2018,38(12):3773-3779.
[16]陈 娟,潘开文,辜 彬. 逆境胁迫下植物体内脱落酸的生理功能和作用机制[J]. 植物生理学通讯,2006,42(6):1176-1182.
