四川贡嘎山国家级自然保护区白马鸡与血雉的时空生态位分化
2022-05-07李永东杨旭贾国清王宇郑笑傲蒋勇杨彪李生强
李永东, 杨旭, 贾国清, 王宇, 郑笑傲, 蒋勇, 杨彪, 李生强
1. 四川贡嘎山国家级自然保护区管理局,四川 康定 626000;
2. 成都兴艾信息技术有限公司,四川 成都 610051;
3. 西华师范大学,西南野生动植物资源保护教育部重点实验室,四川 南充 637002;
4. 四川省大熊猫科学研究院,四川 成都 610057;
5. 广西师范大学,广西珍稀濒危动物生态学重点实验室,广西 桂林 541004
作为生态学中重要的理论之一,生态位研究有助于评估物种在特定资源环境下多维空间中综合利用资源的能力和竞争水平[1-3],目前已发展成为科学理解群落结构、分析种内和种间竞争关系及共存机制、评估种群在群落中地位等方面的重要手段[4-6]。根据生态位理论,同域分布且生态需求相近的物种,通过生态位分化可有效减少或避免有限资源条件下的竞争[7],且必须通过一定的生态位分化才可达到稳定共存[8]。科学对比同域分布近缘物种的生态位特征与分化程度,对于针对性保护与管理策略的制定具有重要的现实意义[9]。
横断山区是我国重要的生态安全屏障区,是全球36个生物多样性热点地区之一(https://www.conservation.org/priorities/biodiversity-hotspots)。 贡嘎山是横断山区中断大雪山的主脉,该区域物种丰富且特有性高[10-12]。早期的研究已发现该地区分布有白马鸡(Crossoptilon crossoptilon)与多种雉类物种共存现象[13],且白马鸡和血雉(Ithaginis cruentus)的优势度较高(见表1)。白马鸡和血雉同为鸡形目(Galliformes)雉科(Phasianidae)物种,同属古北界,均为留鸟,同被IUCN濒危物种红色名录列为近危(Near Threatened, NT)等级,分布型均为喜马拉雅-横断山区型[14,15]。二者还具有相近的体型特征和食性需求[16],同域分布情况下很可能成为潜在竞争关系,因此可以作为研究大型雉类物种生态位分化的理想选择。白马鸡和血雉同为国家II级重点保护野生动物,且白马鸡为我国特有种[16],二者本身具有重要的保护意义和研究价值。国内关于白马鸡和血雉的研究报道较早,在生境选择和空间分布等方面已有部分研究[17-22]。然而,目前关于二者同域分布情况下的生态位分化研究仅限于生境选择和空间利用(均为生境维度)上[17,19,22],尚缺乏食物和时间维度[23]上的研究报道。
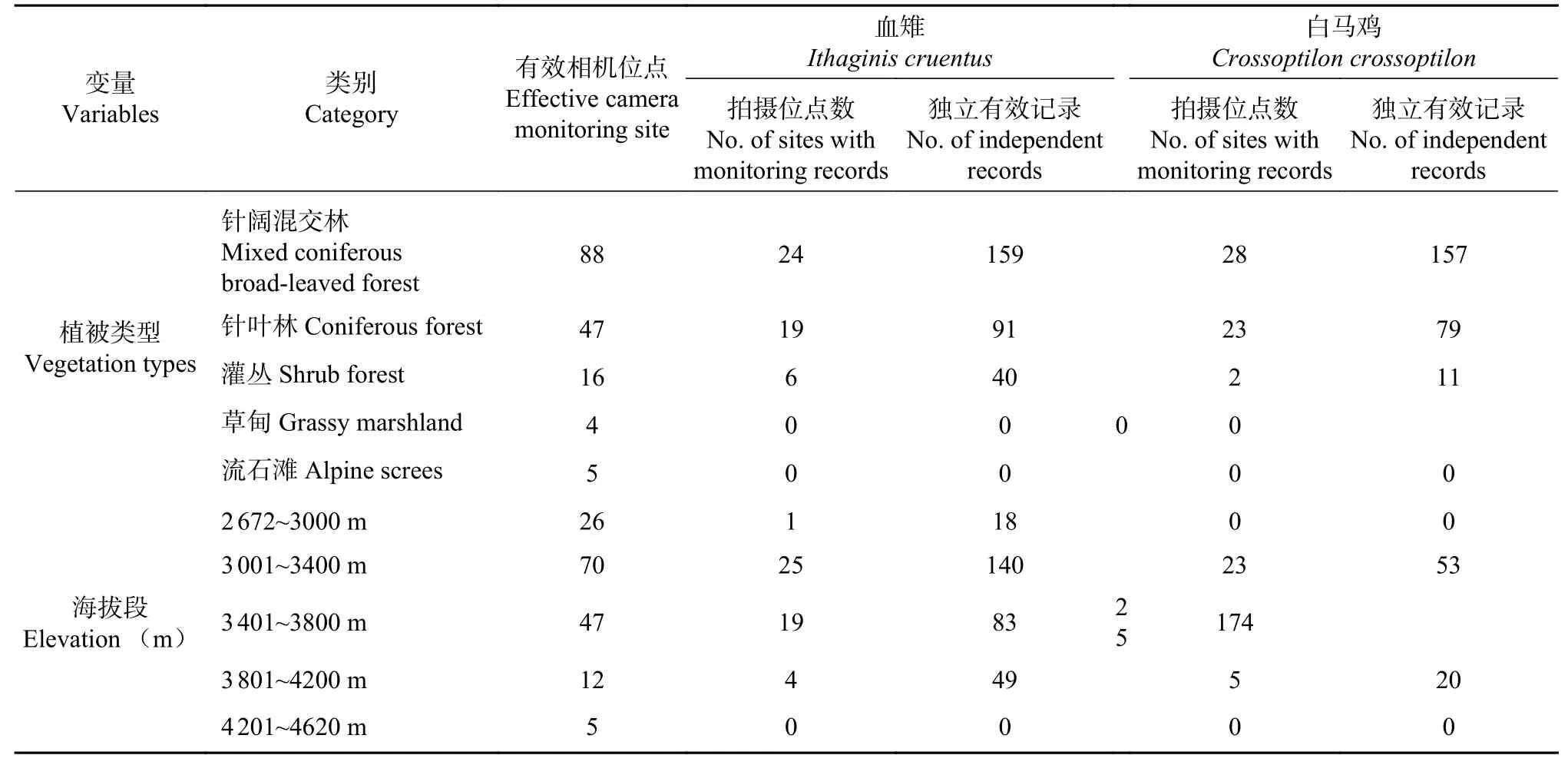
表1 不同植被类型和海拔段下相机位点分布和物种监测情况Tab. 1 Distribution of camera-trap sites and species monitoring conditions in different vegetation types and elevations
以往的研究主要基于传统调查方法(如样带法)开展,在针对目标物种全天候24 h分布和行为数据的获取上具有一定局限,而红外相机技术在这方面具有明显优势[24-25]。本研究在四川贡嘎山国家级自然保护区利用红外相机技术对白马鸡和血雉开展持续一年的监测研究,利用红外相机数据来比较分析二者在分布海拔与植被类型以及日活动节律方面的差异性,为进一步探究同域分布下近缘物种的共存机制和针对性保护管理策略的制定提供重要的研究数据。
1 研究方法
1.1 研究区概况
四川贡嘎山国家级自然保护区位于青藏高原的东南缘(101°29'—102°12' E, 29°1'— 30°5' N),总面积4 091.4 km2,保护区以保护高山生物多样性及多元生态系统为主,主要保护对象为大雪山系以贡嘎山为主的森林生态系统、各类珍稀野生动植物资源、低海拔现代冰川。区内山体高大,谷岭高差悬殊,主峰海拔7 556 m,是青藏高原东部的最高峰和东亚地区的第一高峰。保护区属温带高原气候,气候变化较大,每年6—10月为雨季,11月至翌年5月为旱季。年降水量800~900 mm,多集中在7、8、9三月。区内具有复杂多样的植被垂直带谱且东西坡差异明显,东坡植被垂直带谱完整而复杂,海拔1 000~2 200 m为常绿阔叶林带(含河谷灌丛)、2 200~2 500 m为山地针阔叶混交林带、2 500~3 600 m为亚高山针叶林带、3 600~4 600 m为高山灌丛草甸带、4 600~5 100 m为高山流石滩稀疏植被带、5 100 m以上为永久冰雪带。西坡植被类型及垂直带谱较简单,包括2 800~4 000 m亚高山针叶林带、4 000~4 600 m高山灌丛草甸带、4 600~5 200 m高山流石滩稀疏植被带以及5 200 m以上的永久冰雪带[26-28]。
1.2 相机布设与数据收集
2017年5月~2018年4月,结合当地以往监测、野生动物的活动痕迹、人力可到达性以及红外相机在野外环境的布设效果等[29],选择了160个相机位点安装红外相机(Ltl 6 210),研究期间红外相机覆盖海拔区间为2 672~4 620 m,涉及针阔混交林(88台)、针叶林(47台)、灌丛(16台)、草甸(4台)、流石滩(5台)5种植被类型(见图1)。保证相邻红外相机位点间距至少300 m,安装高度控制在0.5~1.3 m,不使用引诱剂,安装时保证相机正前方不被遮挡,最佳聚焦拍摄点距离相机镜头约4~6 m。相机统一设置为混合模式(3张照片+1段视频),视频长度设置为10 s,拍摄时间间隔为2 s。详细记录每个相机位点的编号、生境、坐标等信息。每3~5个月收取一次数据,同时检查相机性能和更换储存卡。
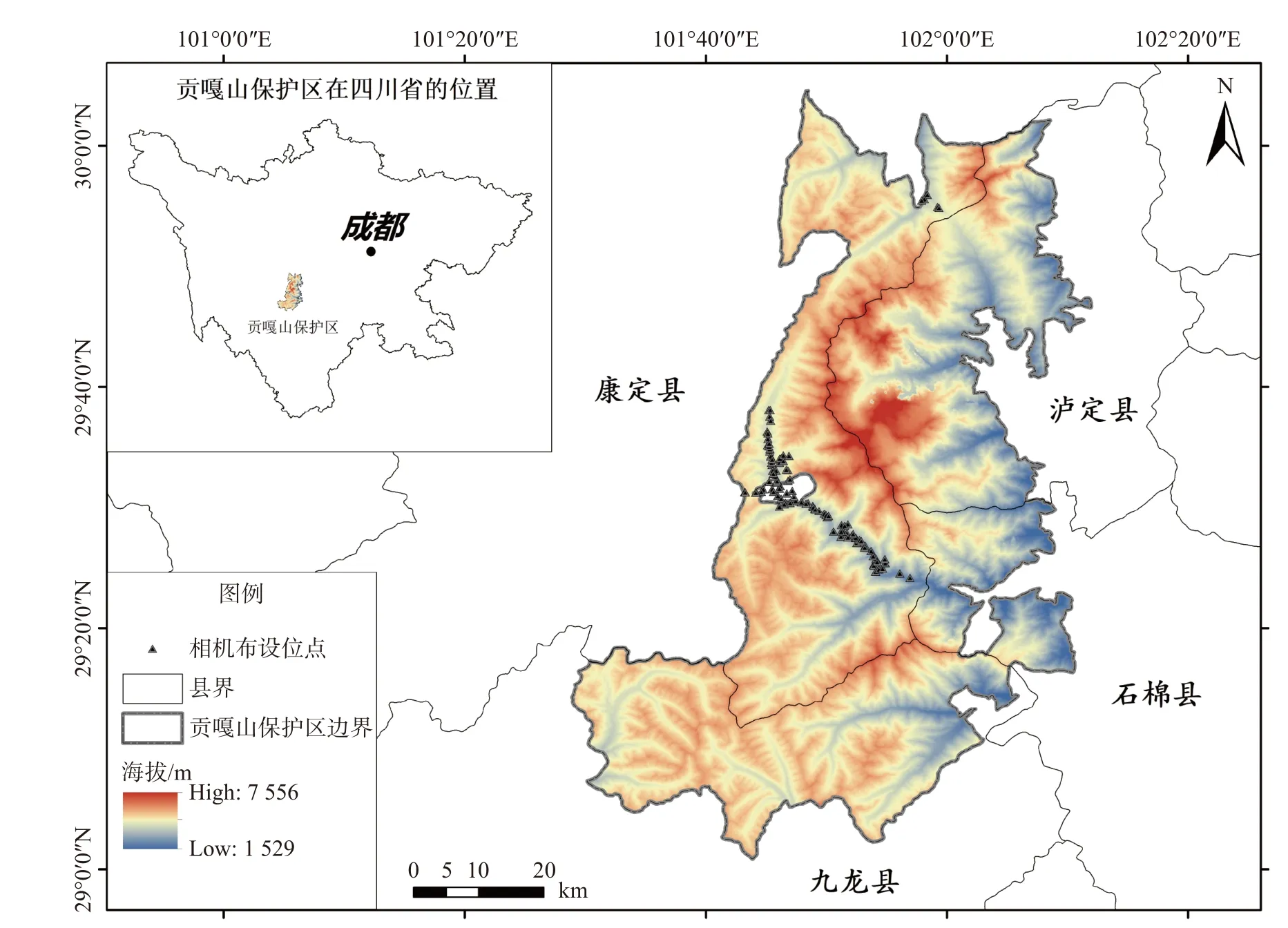
图1 研究期间红外相机布设位点Fig. 1 Distribution of infrared camera traps in Gongga Mountain National Nature Reserve, Sichuan province
1.3 数据分析
将研究期间回收的所有红外相机数据与点位信息进行系统归类整理,统一利用四川自然保护红外相机数据管理信息化平台(http://www.datawild.cn:9 090/Bioplatform/)[30]完成监测数据的录入、影像数据的物种识别与相关信息挖掘,参照《中国哺乳动物多样性(第2版)》[31]和《中国鸟类观鸟手册》[32]对红外相机拍摄的鸟兽物种进行识别并分类,最后从平台上导出所有数据Excel汇总表用于后期分析。为避免过高估计物种的探测率,将同一相机位点下同一物种连续30 min (±1min)的拍摄记录(包括照片或视频)作为1次独立有效记录且每次独立有效记录与红外相机拍到的动物个体数量无关[33]。
1.3.1 空间利用
充分结合整个红外相机布设海拔区间,按照每400 m划分一个水平,包括2 672~3 000 m、3 001~3 400 m、3 401~3 800 m、3 801~4 200 m、4 201~4 620 m,共5个海拔段。采用k个独立样本的非参数检验(Kruskal-Wallis H test)来分析同一季节不同物种之间以及单一物种不同季节之间的差异性,进而对比白马鸡和血雉对不同海拔段的利用情况。详细统计白马鸡和血雉在不同季节不同植被类型下的独立有效记录数,并分析其独立有效记录百分比,通过对比白马鸡和血雉在不同植被类型中被拍摄到的独立有效记录百分比,进而分析不同季节下两物种对不同植被类型的利用情况。
1.3.2 日活动节律
采用核密度估计方法(Kernel density estimation)分析白马鸡和血雉的日活动节律,使用R软件中的overlap包[34]和activity包[35]绘制物种的核密度曲线图。采用条件密度等值线法来确定物种的活动高峰,通常以50%的内核阈值来计算活动比例集中的周期并以此来表征活动高峰期,条件密度等值线图利用R中的circular包“modal. region”函数来实现[36]。采用activity包“overlapEst”函数来绘制日活动节律的重叠图,并利用该函数的分布曲线重叠面积比Δ(当Δ=0时,表示完全分离;当Δ=1时,表示完全重叠)来表示同一季节不同物种以及单一物种不同季节的日活动节律重叠程度,重叠指数根据成对物种较小的样本数选取,当样本量小于50时采用Dhat 1值,当样本量大于75时采用Dhat 4值[37-38]。利用activity包中的“compareCkern”函数的Wald test对白马鸡和血雉日活动节律进行概率检验,循环检测设定为1 000次[35],以此来分析日活动节律的差异性。
所有检验的差异显著水平设定为P<0.05,统计分析和作图均在R 4.1.0中完成[39]。
2 结果与分析
2.1 监测概况
研究期间共计160个有效相机位点,拍摄到白马鸡和血雉的相机位点数分别有53个与49个,二者均被拍摄到的相机位点数有24个。共计获得白马鸡的独立有效记录247次,血雉290次。本次监测均仅在针阔混交林、针叶林和灌丛3种植被类型中发现白马鸡和血雉活动,未在草甸和流石滩生境以及4 200m以上海拔段发现二者活动(见表1)。
2.2 空间利用差异
研究发现,白马鸡和血雉均对针阔混交林表现出明显偏好。白马鸡对针阔混交林的利用率全年最高,雨季占比更多(旱季:56.05%;雨季:76.67%);其次是针叶林,但旱季占比大于雨季(旱季:40.13%;雨季:17.78%);而全年对灌丛植被的利用率均较低。血雉在旱季更多出现在针叶林(47.06%)和针阔混交林(42.94%)中,对灌丛植被的利用率仅为10.00%;而在雨季主要在针阔混交林(71.67%)中活动,针叶林的利用率仅为9.17%,但灌丛植被的利用率有所提高,达到了19.17%(见图2)。
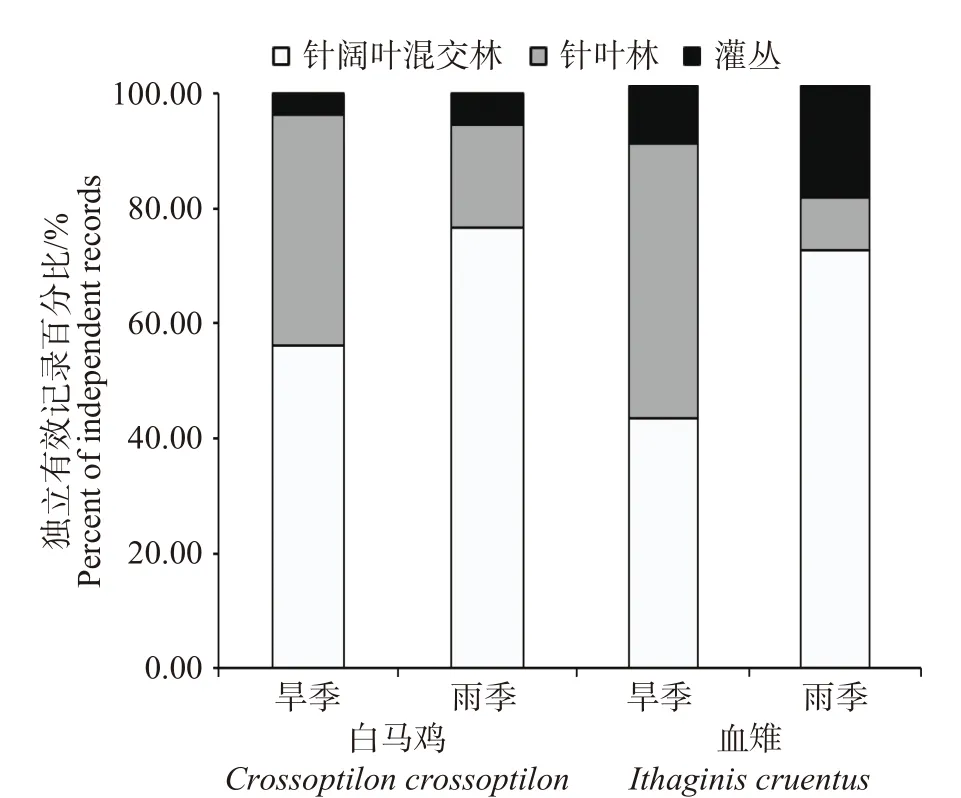
图2 白马鸡和血雉在不同季节对不同植被类型的利用对比Fig. 2 Comparison of different vegetation types utilized by Crossoptilon crossoptilon and Ithaginis cruentus in different seasons
血雉(2 981~3 930 m)和白马鸡(3 067~3 930 m)分布的海拔范围相近,但白马鸡平均分布海拔显著更高(3 600.44 m ± 227.97 m vs. 3 412.67 m ± 313.32m;Z= -7.022,P<0.01)。血雉在雨季的活动海拔范围略高于旱季,但差异不明显(3 415.62 ± 366.95 m vs.3 410.59 m ± 269.08 m; Z= -0.058,P=0.953)。白马鸡在雨季的活动海拔范围明显高于旱季(3 654.96 ±209.48 m vs. 3569.19 m ± 232.21 m; Z= -3.248,P=0.001)(见图3)。
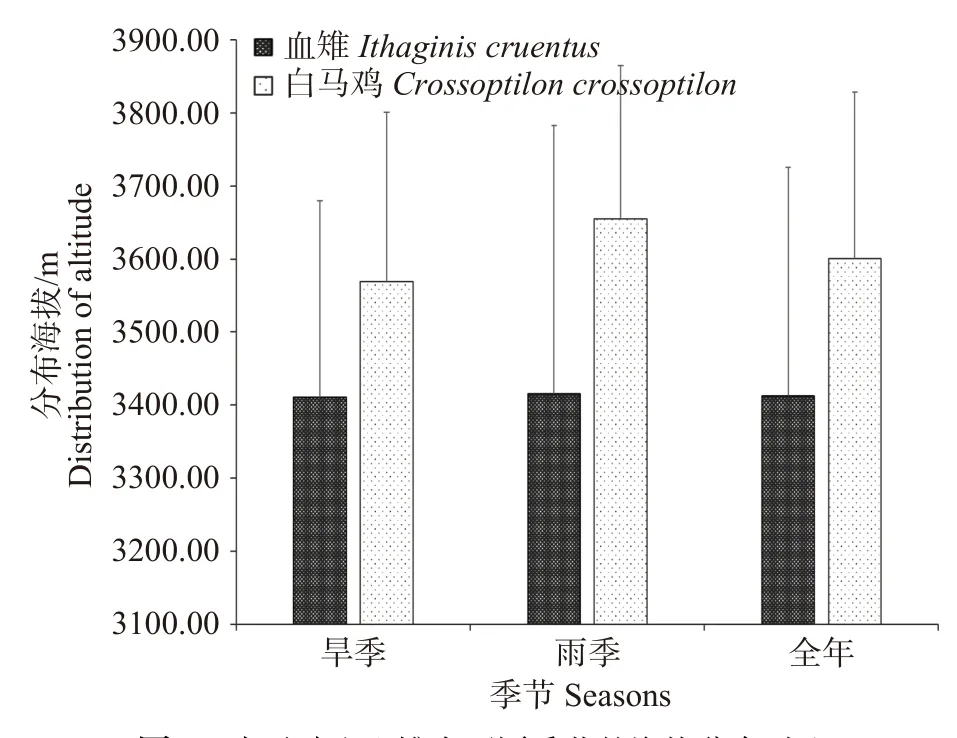
图3 白马鸡和血雉在不同季节的海拔分布对比Fig. 3 Comparison of altitude distribution of Crossoptilon crossoptilon and Ithaginis cruentus in different seasons
2.3 活动节律差异
2.3.1 日活动节律整体比较
统计分析发现,白马鸡和血雉的日活动节律曲线存在极显著差异(Δ=0.85,P=0.002<0.01),表明二者的日活动节律模式相似度较低(见图4)。白马鸡的日活动节律表现出3个活动高峰,分别在早晨(7:00—9:00)、中午(11:00—13:00)、傍晚(19:30—20:30),且早晨活动峰值最大,傍晚活动峰值最小。23:00—05:00几乎没有发现活动迹象,表明白马鸡属于典型的昼行性动物且具有一定的晨昏活动习性(见图4 A)。血雉的日活动节律呈现出1个明显的活动高峰(7:00—13:00),在20:00左右有一个小的活动波动,23:00—05:00活动迹象少甚至部分时段未见活动迹象,表明血雉属于典型的昼行性动物(见图4 B)。

图4 白马鸡(A)和血雉(B)的日活动节律分布图Fig. 4 Daily activity pattern of Crossoptilon crossoptilon (A) and Ithaginis cruentus (B)
2.3.2 日活动节律的季节性差异比较
白马鸡在旱季的活动峰值要大于雨季,雨季比旱季多一个活动高峰(约12:00)。血雉在雨季的活动高峰(约11:00)要比旱季(8:00)晚3个小时。整体上白马鸡和血雉在季节间日活动节律均不存在显著差异(白马鸡:Δ=0.89,P=0.40, 图5 a; 血雉:Δ=0.89,P=0.29, 图5 b)。白马鸡和血雉的日活动节律在雨季无显著差异(Δ=0.84,P=0.07, 图5 d),而在旱季存在显著差异(Δ=0.86,P=0.04, 图5 c)。

图5 白马鸡和血雉的日活动节律比较:单一物种季节间对比(a和b)以及单一季节物种间对比(c和d)Fig. 5 Comparison of daily activity rhythm between Crossoptilon crossoptilon and Ithaginis cruentus: seasonal comparison of single species (a and b) and comparison between species in a single season (c and d)
3 讨论
研究同域分布物种的生态位分化,对于物种的区域整合保护与管理具有重要的现实意义[40-41]。本研究以四川贡嘎山国家级自然保护区内同域分布的白马鸡和血雉为研究对象,发现二者的独立有效记录数均较高且数量相近,分布相同的相机位点数有24个,因此在对比分析同域分布雉类物种生态位差异上具有一定代表性。生态位差异分析发现,二者在生境维度(分布海拔与植被类型)和时间维度(日活动节律)均具有一定差异,表明二者存在一定的生态位分化。
以往的研究指出,血雉的活动海拔在2 000~4 500 m,最高可至雪线区域,而白马鸡主要在3 000~4 000 m海拔段活动,血雉活动海拔范围更宽[42]。本研究中,血雉(2 981~3 930 m)和白马鸡(3 067~3 930 m)分布的海拔范围相近且分布海拔均较高,体现出二者在贡嘎山保护区特殊的空间分布,这种分布差异可能受不同地区植被组成和空间异质性的影响[43]。已有研究发现,同域分布物种的生态位分化主要在空间、食物和时间3个尺度上体现,而近缘物种同域分布时其生态位分化首先会体现在空间分布差异上[23],由于时间具有刚性不易改变的特性,因此在有限的活动空间或者受到干扰的情况下,种间竞争导致的动物日活动节律的改变往往被认为是最后被启动的共存机制[44-45]。白马鸡分布的相机位点数和平均分布海拔要高于血雉(见图3),其日活动节律表现出3个活动高峰,与血雉仅有的1个活动高峰明显不同(见图4),二者在时空生态位上的差异变化同样印证了上述规律。
白马鸡和血雉均会在针叶林、针阔混交林和灌丛植被类型中活动[42],本研究发现二者均仅在以上3种植被类型中活动,且植被类型偏好程度均表现为:针阔混交林>针叶林>灌丛,整体分布生境上具有一定相似性,但二者不同植被类型的利用率不同且旱雨季之间存在差异。一般来讲,生境越复杂,空间异质性越高,其中蕴藏的食物资源也会越丰富,具备野生动物栖息的场所种类越多样,会吸引更多的野生动物分布,而复杂的生境下也有利于野生动物隐蔽以降低被捕食的概率[46]。在白马鸡和血雉分布的3种植被类型中,针阔混交林是最为复杂的生境类型,这可能是二者均偏好在针阔混交林分布的重要原因。这种生境偏好,在雨季(6—10月)表现得尤为突出(二者在雨季对针阔混交林的利用率均超过了70%),这可能与不同季节下环境差异有关,包括食物资源、气温变化等[47]。贡嘎山保护区旱季(11月至翌年5月)气温整体较低,尤其是3 000 m以上的区域会受霜雪影响,食物资源相对短缺;而雨季气温相对较高,许多被积雪覆盖的地面会逐渐露出来,许多植物会长出新叶和新芽,食物资源变得相对丰富。旱季里,白马鸡和血雉对针叶林的利用率均较高,尤其是血雉(占比达47.06%),可能原因是旱季气温较低,在厚重的雪盖下,高大的针叶林更利于遮挡积雪,其林下更易寻找食物。雨季里,白马鸡和血雉对灌丛的利用率均有提高,尤其是血雉(达到19.17%),这可能与该季节灌丛植被中可提供的食物资源增多有关。不同季节下白马鸡和血雉的活动节律存在差异,表明不同季节下的环境变化不仅会影响物种的生境偏好,而且会影响不同季节下物种的活动强度,尤其是食物资源较为短缺、气温较低的旱季。在寒冷的旱季,动物往往需要增加活动强度来寻觅足够的食物[48-49],这种行为差异可能是物种应对特定环境中有限资源竞争而采取的适应性取食策略[50-50]。
本研究为同域分布雉类物种生态位分化研究积累了一定的基础数据,但主要在于描述分化差异,其分化差异背后的生理机制和生存策略可能还需要积累多方面数据来加以解释。而同域分布物种的生态位分化程度在不同地理区域和不同年份下可能不同,因此,未来的研究工作可以关注其他区域的生态位分化研究来科学对比,同时加强基础数据的积累,如利用红外相机对研究区域内白马鸡及同域野生动物开展持续监测。最后,鉴于本研究中对高海拔区域(如高山草甸和流石滩)布设相机数量较少,可能带来对高海拔段监测强度不够的影响,特建议在未来的监测工作中适当增加高海拔区域相机监测位点以积累更加全面的科学数据。
